

Full Length Research Article
Effect of Cadmium and Copper on Morphological and Physiological Parameters of Stevia rebaudiana Bert. Under in Vitro Conditions
Afshan Shahzadi*, Ayishm Altaf, Humera Afrasiab
Adv. life sci., vol. 9, no. 4, pp. 473-482, December 2022
*– Corresponding Author: Afshan Shahzadi (Email: Afshan.phd.botany@pu.edu.pk)
Authors' Affiliations
[Date Received: 10/09/2021; Date Revised: 01/11/2022; Date Published: 31/12/2022]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Stevia rebaudiana Bertoni is a popular non-caloric sweetener being used all over the world for its anti-diabetic properties. Stevia contains phytochemical compounds known as diterpene glycosides that helps to reduce blood sugar, cholesterol, and blood pressure. In vitro raised cultures are considered as an efficient way to produce essential secondary metabolites, especially through elicitation. This study aims to optimize culture conditions for the large-scale production of these economically significant metabolites using abiotic elicitors.
Methods: Apical shoot, nodes as well as leaf explants from Stevia mother plants were used for micropropagation and callogenesis respectively. Murashige and Skoog (MS) medium supplemented with different cytokinins and auxins alone or in combinations were tried for obtaining the maximum yield of calli and plantlets. The calli and plantlets were subjected to different concentrations of copper sulphate and cadmium chloride. After 40 days of treatment, the data was recorded for morphological characters of elicitor-treated plants and calli. Chlorophyll content and antioxidant activity using DPPH assay were performed using treated leaves and calli extracts.
Results: Among all the cytokinins tried for micropropagation, BAP at 0.5 mg/L in MS medium proved to be the best with 100% response by inducing 34 and 40 shoots/culture within 12 days of inoculation from shoot tip and nodal explants, respectively. For rooting, MS medium containing 0.6 mg/L of IBA was best producing 15-17 roots/culture within 10 days. Green friable and embryogenic calli was obtained in MS medium containing 1.0 mg/L TDZ with 0.1 mg/L NAA from leaf explants. A significant increase was observed in the elicitors-treated calli and plantlets biomass, chlorophyll content as well as antioxidant potential as compared to the control.
Conclusion: The results of this study indicate that the cytokinins and auxins at lower concentrations gave significant results for micropropagation and callogenesis. Further, elicitation with copper sulphate and cadmium chloride has improved the biomass and antioxidant activity of calli and plantlets.
Keywords: Stevia; Micropropagation; Callus induction; Elicitation; Antioxidant activity; Biomass
Introduction![]()
Stevia rebaudiana Bertoni became popular all over the world during the twenty-first century as a powerful non-caloric and anti-diabetic sweetener [1]. It is native to Brazil and Paraguay where approximately 2500 of the 20,000 native plant species are being traditionally used for medicinal purposes by the native population [2]. Traditional medicine, derived from medicinal plants, is used being used by almost 80% of the world’s population [3]. It is commonly known as sugar leaf plant, sweet herb, or sweet treat, and belongs to the family Asteraceae, featuring sweet leaves which is claimed as a perfect sugar substitute [4-6]. The leaves of the Stevia plant contain several secondary metabolites, including steviol glycosides, notably stevioside and rebaudioside A, which are non-glycemic and are more than 300 times sweeter than table sugar [7]. Besides this, it has many therapeutic values: it is reported as having anti-aging [8], anti-cancerous [9,10], anti-hypersensitive [9,11] as well as contraceptive properties [12], and also helpful in preventing dental caries [13]. It also has anti-bacterial and anti-fungal potential [14]. Stevia plants are traditionally cultivated via cuttings due to poor germination rates, resulting in lower yields [1]. Tissue culture is an efficient and alternative approach for mass production of Stevia in a short period of time [15,16]. Similarly, in vitro culture systems based on plant tissue culture serve as a good source for the generation of secondary metabolites, preserving natural resources [17]. Plant calli cultures are viewed as viable source for the long-term production of pharmaceutically important secondary metabolites [18].
For commercialization of the plant cell cultures, accumulation of high secondary metabolite contents is essential. The incorporation of elicitors in plant cultures is believed to be the most efficient way for fixing productivity concerns. Although various elicitation strategies are available, the enhancement of bioactive compounds using chemical elicitors such as metal ions, other inorganic compounds, or even UV radiation and electric current is predominantly considered because of its high efficiency and yield [19]. Oxidative stress is caused by the production of free radicals and other reactive oxygen species in the human body. It is a significant risk factor in the pathogenesis of many chronic diseases such as aging [20,21], asthma, inflammatory arthropathies, Parkinson’s and Alzheimer’s diseases, cancers, atherosclerosis, and diabetes mellitus [22-24]. According to the recent report of World Health Organization, diabetes alone was the direct cause of 1.5 million deaths [25]. Aside from hyperglycemia, increased oxidative pressure is also thought to play a role in the parthenogenesis of this illness [26]. Recently, dietary antioxidants capable of suppressing active oxygen radicals that intervened in oxidative stress have received a lot of attention [27]. Therefore, the Stevia plant is the best choice to work on its antioxidant potential for the yield of natural and safe antioxidants to lower the risk of diseases.
The present investigation was carried out to standardize protocols for in vitro production of Stevia plants and calli cultures. Two chemical elicitors (CuSO4 .5H2O and CdCl2) were used to enhance the antioxidant potential of Stevia. After elicitation some physiological and biochemical parameters such as chlorophyll content and antioxidant activity through DPPH (2,2-diphenyl-1-picryl-hydrazyl-hydrate) assay were studied.
Methods![]()
Parent material and growth conditions
The field grown plants of Stevia (S. rebaudiana) were obtained from Ayub Agriculture Research Institute, Faisalabad and maintained in a glasshouse at the Institute of Botany, University of the Punjab, Lahore, Pakistan. Shoot tips and nodal explants (15-30 mm) were excised and washed with running tap water and multiple rinses were given in a glass jar for about 8-10 minutes so that the explants become dust-free. In the next step 1% (v/v) Tween 20 was added as surfactant and kept on a magnetic stirrer for about 3-5 minutes, decanted the solution and rinsed with distilled water to remove all the surfactant. The explants were then submerged in 5% sodium hypochlorite (NaOCl) solution for 3-5 minutes, rinsed with distilled water, and finally treated with 0.1% mercuric chloride (HgCl2) for 3 minutes followed by 3-5 times washing with autoclaved distilled water. After decantation the explants were trimmed from both sides to remove the dead part and inoculated in 20×150 mm size culture tubes, each having 15 mL of MS (Murashige and Skoog) [28] medium supplemented with 0.6 mg/L IBA (Indole-3-butyric acid) and 3% sucrose and 2.5 % phytagel (Sigma cell culture). The pH of the medium was adjusted between 5.7-5.8 before autoclaving at 121°C (15 lb/inch2) for 15 minutes. Culture tubes were incubated in the culture room with 16 hours photoperiod at 25+2°C. After 1 month, the plants were ready to use for further experiments.
Micropropagation
For micropropagation and multiple shoot formation, the shoot tips and nodal explants of in vitro-grown plant were inoculated on the MS medium containing different cytokinins in different concentrations. Two cytokinins Kinetin (KN), 6-Benzylaminopurine (BAP) in the concentrations of 0.5, 1.0, 1.5 mg/L were used. For rooting, NAA (1-Naphthaleneacetic acid), IAA (Indole-3-acetic acid) and IBA (Indole-3-butyric acid) were used in different concentrations.
Callus cultures
For callus induction from leaf explants of Stevia, 5-10 mm size explants were cultured on Murashige and Skoog (MS) medium supplemented with different plant growth regulators (NAA, 2,4-D, BAP, TDZ) in different combinations. The culture vessels were kept in a temperature-controlled culture room (25±2 °C). The data was recorded after one week of culturing and calli were maintained through subsequent sub-culturing.
Preparation of elicitors
YE, CH, PEC, and ALG (Sigma Aldrich) were dissolved with double distilled water and the pH of the elicitor solutions was adjusted to 5.8. The MeJA and SA (Sigma Aldrich) were dissolved in 96 % ethanol and the solutions were then diluted with double distilled water in concentrations as required. Finally, the pH of both solutions was adjusted to 5.8. Stock solution of CHI (from crab shells) was prepared in 2.0 mL of 1 % (v/v) acetic acid (Sigma-Aldrich) by stirring with a glass rod and then diluted with double distilled water. Finally, the pH of the solution was adjusted to 5.8. The stock solutions of ALG, MeJA, SA, and CHI were filter-sterilized through a 0.22 lm syringe Millipore filter (MinisartÒ, Sartorius, Germany), and then added to the autoclaved WPM aseptically at the desired concentrations. The stock solutions of PEC, YE and CH were added to the media at the desired concentrations before autoclaving at 121 °C for 15 min. In the present study, ALG, CH, PEC or YE at the concentrations of 0.5–2.0 g/L and CHI, MeJA or SA at the concentrations of 50–100 lM was added individually in semi-solidified WPM as elicitors
Preparation of elicitors
YE, CH, PEC, and ALG (Sigma Aldrich) were dissolved with double distilled water and the pH of the elicitor solutions was adjusted to 5.8. The MeJA and SA (Sigma Aldrich) were dissolved in 96 % ethanol and the solutions were then diluted with double distilled water in concentrations as required. Finally, the pH of both solutions was adjusted to 5.8. Stock solution of CHI (from crab shells) was prepared in 2.0 mL of 1 % (v/v) acetic acid (Sigma- Aldrich) by stirring with a glass rod and then diluted with double distilled water. Finally, the pH of the solution was adjusted to 5.8. The stock solutions of ALG, MeJA, SA, and CHI were filter-sterilized through a 0.22 lm syringe Millipore filter (Minisart Ò, Sartorius, Germany), and then added to the autoclaved WPM aseptically at the desired concentrations. The stock solutions of PEC, YE and CH were added to the media at the desired concentrations before autoclaving at 121 °C for 15 min. In the present study, ALG, CH, PEC or YE at the concentrations of 0.5–2.0 g/L and CHI, MeJA or SA at the concentrations of 50–100 lM was added individually in semi-solidified WPM as elicitors
Elicitor Preparation and Treatments
The stock solutions of the two elicitors, CuSO4 5H2O (Panreac) and CdCl2 (Beijing Chemical Works), were prepared in sterile distilled water and stored in a refrigerator. After adding elicitor solutions in different concentrations (10 µM, 25 µM, 50 µM, 75 µM, 100 µM, 150 µM, 200 µM/L) to the MS medium supplemented with different hormones for shoots and callus respectively. Treatments of S. rebaudiana plantlets with elicitors were performed with one month old in vitro grown plants whereas for calli treatments, one-and-a-half-month old callus was used. For the control, elicitors were not added in the medium with growth regulators. Control plants and treated plants and calli were then harvested after 40 days of elicitation.
Morphological Characters
At the time of harvest, plant height, number of leaves, leaf area, internodal length, fresh weight (FW) and dry weight (DW) of leaves/plant were calculated. For calli and leaves biomass analysis, fresh weight of leaves and callus cultures was noted, and the initial weight of callus and leaves were subtracted from final weight (WC=Wf-Wi). For dry weight (DW), the pre-weighed leaves and calli were placed in an oven at 70℃ for 2 days to measure the dry weight.
Biochemical Characters
Estimation of Chlorophyll content
Chlorophyll was extracted by incubating 0.05 g fresh leaves and calli in 5.0 mL dimethyl sulfoxide (DMSO). The test tubes were incubated in dark at room temperature for 72 hrs. The absorbance of chlorophyll extract was measured at 663 nm and 645 nm with a spectrophotometer. Total chlorophyll content (mg/g fresh weight) was calculated from the absorbance at 663nm (D663) and 645nm (D645) using the formula of Arnon [29]. Blank determination was carried out with DMSO.
DPPH (2,2-diphenyl-1-picryl-hydrazyl-hydrate) Assay
Extracts were prepared by the given procedure. After 40 days, the leaves and calli from culture tubes were harvested and dried in an oven at 40 ᵒC for 48 hours. With the help of pestle and mortar, 2.0 g dried leaves and callus of each concentration of all elicitors were grinded to fine powder. Then the powder was subjected to Soxhlet apparatus in which 50 mL methanol was used as extractant. Methanol was evaporated with rotary evaporator and after weighing, the extracts were preserved at -20ᵒC further use [30].
The antioxidant activity of leaves and calli extracts of Stevia was determined by using the DPPH free radical scavenging test [31]. All extracts of different concentrations were further diluted (30, 60, 90, 120,150, 180, 210, 240, 270, 300 µg/mL) and transferred to separate test tubes. In each test tube, 1.0 mL of these doses and 3.0 mL of DPPH (0.1 mM) were added and after shaking kept for 30 minutes incubation in dark at room temperature. Blank was prepared with the same procedure but without any extract. All the stocks and dilutions were made in high grade methanol. BHT (Butylated hydroxytoluene) and BHA (Beta hydroxy acids) were used as standards. Absorbance at 517 nm was measured with a spectrophotometer (Hitachi). The percentage of inhibition was calculated by the following formula:

Statistical Analysis
The treatments were set up in a completely randomized design with three replicates per treatment. Significant differences among the treatments were determined by Analysis of Variance (ANOVA) followed by Duncan’s Multiple Range test at a significant level of p ≤ 0.05 (n = 3) using IBM SPSS Statistics data editor (V.21).
Results![]()
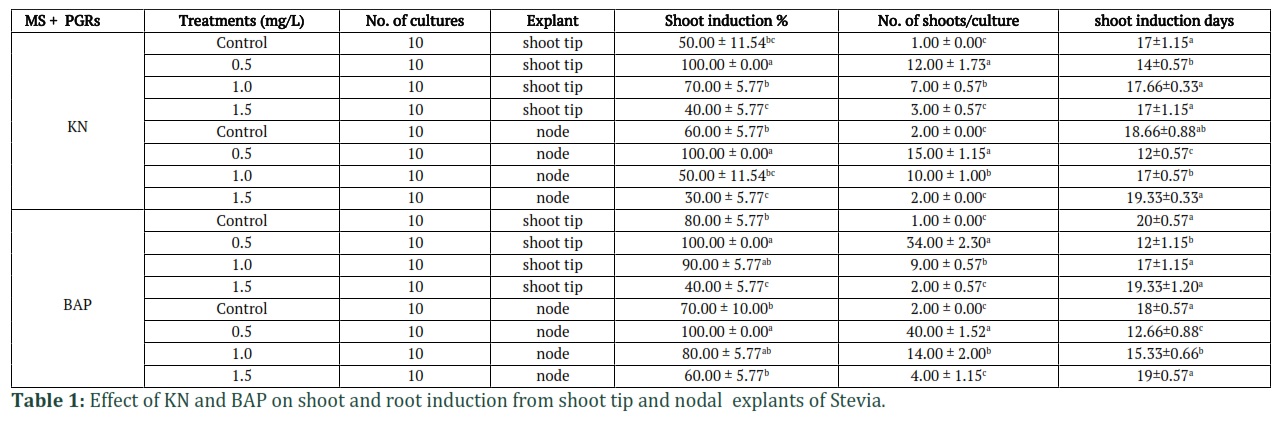
Multiple Shoot induction
The data of different hormones used for micropropagation of S rebaudiana from apical and nodal explants showed significant (p≤0.05) variation in terms of number of shoots and number of days for shoot induction (Table 1). The findings revealed that both the hormones tried for micropropagation gave 100% response at their lower concentration (0.5 mg/L), the only difference was seen with respect to number of induction days and number of proliferated shoots/culture vessel. It was observed that Kinetin (KN) at 0.5 mg/L concentration in MS medium induced 100% shoot induction from both shoot tip and nodal explants after 14 and 12 days, with an average of 12 and 15 shoots per culture, respectively (Fig.1a, 1b). MS medium supplemented with BAP at 0.5 mg/L concentration showed better results with respect to the number of shoots from both explants. A significantly high number of shoots from shoot apex (34) and nodal explant (40) per culture tube respectively, were recorded after 12 days of inoculation (Fig. 2a, 2b).
Root induction
The results of for root induction showed that among different auxins (NAA, IAA, and IBA) tried, only the IBA gave better response. Maximum number of roots was obtained on MS medium supplemented with 0.6 mg/L IBA. It produced an average of 15 roots/culture shoot basal part and 17 roots/culture from node derived shoots with 100% induction response (Fig. 3c, 3d) as compared to control which was MS basal medium in all micropropagation media. Also, 70% of medical students believe that herbal medicines (medicinal herbs) for anxiety and depression should be under the supervision of a physician. 75% of medical students believe that the use of herbal medicines for anxiety and depression is only for Iran and 80% of them believe that herbal medicines for anxiety and depression are effective. Almost half of the students believe that herbal medicines for anxiety and depression must be preparation from pharmacies, and the other half do not believe in this (Table 4).
Callus induction
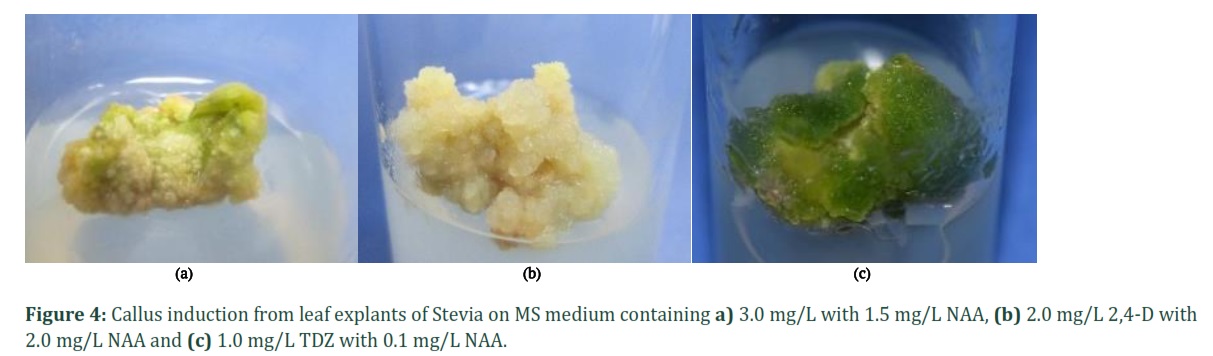
For callus induction from leaf explants of Stevia, different combinations of plant growth regulators (PGRs) were tried. Callus response percentage, time and morphological characters are given in Table 2. The most significant result was observed in MS medium containing 1.0 mg/L TDZ along with 0.1 mg/L NAA, which gave 100% response with green and friable, embryogenic callus induced within 6 to 7 days of inoculation (Fig. 4c). A decrease in the percentage callus induction was observed with an increase in the concentration of TDZ. In another combination of 2, 4-D with NAA, 96% callus induction response was observed using 2.0 mg/L of both the PGRs after 15 days of induction. The callus obtained was creamy to yellow in color and compact in nature (Fig. 4b).
A combination of BAP and NAA at higher concentration gave better results as compared to lower concentrations tried. In MS medium containing 3.0 mg/L BAP with 1.5 mg/L NAA, 96% callus induction rate after 23 days of inoculation. The callus was light green and non-proliferating in nature (Fig. 4a). While at a lower concentration of BAP (0.5 mg/L) with higher concentration of NAA (3.0 mg/L) produced 93% small, green, compact callus after 13 days of culture. The medium with the highest callus response was selected to sub-culture for the treatment with varying concentrations of two chemical elicitors.
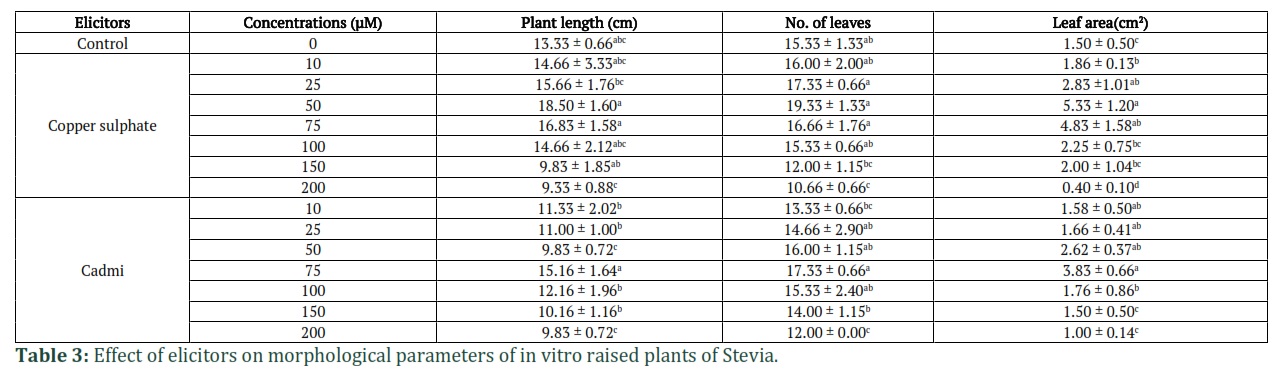
Effect of elicitors on morphological parameters of plants and calli
The data given in Table 3 shows the effect on in vitro grown plants treated with two elicitors, copper sulphate and cadmium chloride, on different morphological parameters. In case of plantlets treated with different concentrations of CuSO4.5H2O, an increase in all the morphological parameters was observed up to 50 µM concentration as compared to control and higher concentrations used. Maximum increase in the height of the plant was from 13.33 cm (control) to 18.5 cm, the number of leaves increased from 15.33 to 18.50, the leaf area of control was 1.5 cm2 which increased to 5.33 cm2 at 50 µM of CuSO4.5H2O.
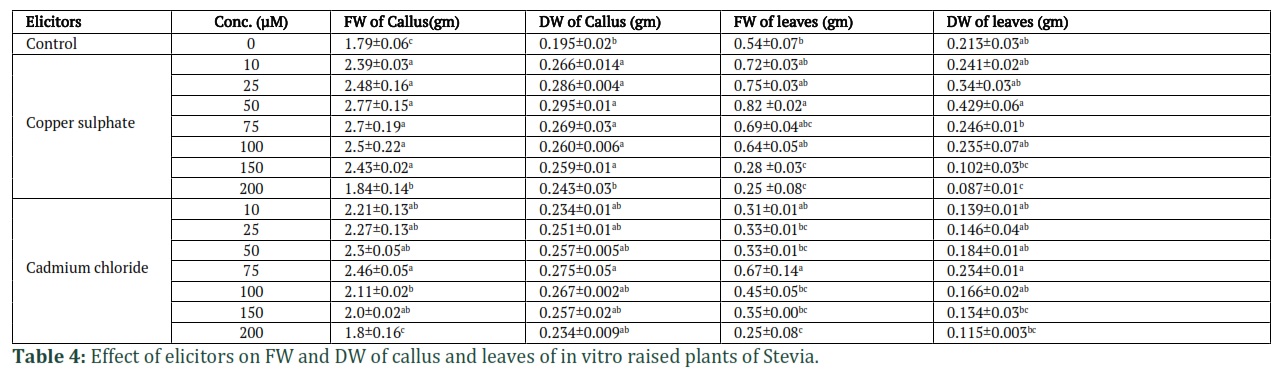
A similar trend of increase in all the parameters up to 75 µM of CdCl2 was recorded. Maximum increase in all morphological parameters was observed at 75 µM concentration of CdCl2 as compared to other concentrations and control (Table 3). However, treatment with higher concentrations of both the heavy metals lead to the decrease in all the parameters studied. As given in Table 4, the calli treated with different concentrations of CuSO4.5H2O, showed an increase in the FW and DW up to 50 µM concentration as compared to control, which decreased with further increase in elicitor concentration. The FW of callus showed an increase from 1.79 gm (control) to 2.77 gm. Similarly, the DW of callus also showed an increase from 0.195 gm (control) to 0.295 gm at 50 µM concentration of CuSO4.5H2O. In case of leaves from elicitor treated plants, an increase in FW up to 50 µM concentration of CuSO4.5H2O was observed which decreased with further increase in elicitor concentration. From 0.54 gm in control, the FW increased to 0.82 gm and the DW increased from 0.213 gm in control to 0.429 gm at 50 µM concentration of CuSO4.5H2O. In case of callus and leaves obtained after treatment with CdCl2, maximum increase in FW (2.46 gm) and DW (0.275 gm) of callus was observed at a lower concentration of 75 µM as compared to control, which decreased with further increase in elicitor concentration. Similar results were recorded for leaves biomass (Table 4).
Effect of elicitors on chlorophyll content and antioxidant activity of plants and calli
As given in Table 5, the total chlorophyll content of calli was highest at 50 µM (48±0.01 mg/g) as well as 75 µM/L (48±0.02 mg/g) of CuSO4.5H2O in comparison with control (0.25 mg/g) and other treatments. Leaves gave increased chlorophyll content as compared to control (0.53±0.08 mg/g) and other concentrations of the CuSO4.5H2O at 100 µM/L (2.53±0.08 mg/g) whereas cadmium chloride treatment exhibited maximum chlorophyll content at 50µM/L (0.53±0.02 mg/g) which was more than the copper sulphate highest treated results. However, the leaf chlorophyll content of CdCl2-treated plants showed a higher amount at 75 µM/L (1.53±0.02 mg/g) than other treatments and control (0.53±0.08 mg/g).
Antioxidant activities of methanolic extracts of elicitor treated Stevia leaf and callus extracts were tested at 30, 60, 90, 120, 150, 180, 210, 240, 270 and 300 ug/mL. In case of copper sulphate, 150 µM/L treatment showed highest DPPH activity with 48.16% for leaf extract and 48.38% inhibition for calli extract, respectively. On the other hand, cadmium chloride treated calli and leaf extracts showed best antioxidant potential at 75 µM/L (87.26%) and 100 µM/L (71.26%), respectively.
Figures & Tables




In the present study, culture conditions were optimized for micropropagation and callogenesis from leaf explants of Stevia. After the optimization of protocols, the in vitro raised plants and calli were subjected to elicitation with two heavy metal salts to increase the antioxidant potential. Due to the commercial importance of Stevia, efforts have been made to increase its production. Conventional methods have not been very successful owing to the very low seed germination percentage [32]. The plants produced via seeds are not homogeneous and show variability in terms of sweetening levels and composition [33]. In addition, vegetative propagation methods also do not meet the demand of individuals required for commercial cultivation [34]. Therefore, there is a need for a reliable procedure and in vitro propagation via plant tissue culture technique is a promising alternative to solve these issues. Among the tissue culture techniques, callus culture can be used directly for regeneration of plantlets and for the extraction of primary and secondary metabolites. Callus and suspension cultures are considered as the basic techniques for the production of desired metabolites from plants [35].
In the present study shoot tip and nodal explants were used for micropropagation as in general, nodal segments are known as the best explants source for plant production under in vitro conditions [26,36,37]. Previous studies reported healthy plantlets of S. rebaudiana obtained through tissue culture using meristem or shoot tip culture [38] and nodal segments [39]. In the present work, significant number (34-40) of shoots were obtained at lower concentration (0.5 mg/L) of BAP as compared to KN, from both the explants. According to Yang and Chang [40], about 50-70 shoots were formed per explant at a higher (2.0 mg/L) concentration of BAP. While Pandey and Chikara [41] also reported that MS medium with 0.5 mg/L BAP produced highest mean number of shoots (30) in Stevia.
It is generally accepted that auxins have a central role in root initiation. In our work, MS medium supplemented with 0.6 mg/L IBA showed 100% rooting from isolated shoots of Stevia. Previously, it is reported that efficient rooting response was observed in half MS medium supplemented with 1.0 mg/L IBA in Stevia [42] which is higher as compared to our results. Similarly, it also reported that S. rebaudiana was rooted on MS medium supplemented with a higher (2.0 mg/ L) concentration of IBA [43].
In the present work, for callus induction from leaf explants of stevia, NAA with BAP, 2,4-D with NAA and TDZ with NAA was tried. Out of the different PGR combinations tried, the best response (100%) was obtained in MS medium containing TDZ (1.0 mg/L) with NAA (0.1 mg/L) from nodal explants of stevia within a week of culturing. The calli obtained was dark green, friable, and embryogenic in nature. Other PGRs combinations tried i.e., NAA with BAP and 2.4-D with NAA also gave significant (96%) callus induction rate but the callus produced was compact and non-proliferating in nature and was initiated after 2-3 weeks of culturing. The combination of NAA with BAP for callus induction has been reported by many authors for Stevia [44-46] while Gupta and his co-authors developed a protocol for callus induction and multiplication and obtained 100% callus induction on MS medium fortified with 0.75 mg/L NAA with 1.0 mg/L 2, 4-D from leaf, nodal and root explants [47]. In our studies a higher concentration (2.0 mg/L) of both these PGRs gave 96% callogenic response, while there is no report about the use of TDZ with NAA for callus induction in Stevia.
Heavy metal salts act as major abiotic stressors that can limit plant growth and development. Cadmium is a toxic heavy metal and once absorbed it causes a negative effect on plants by disturbing several cellular processes resulting in the reduction of plant growth and chlorosis leading to senescence [48]. It causes oxidative stress by generating reactive oxygen species (ROS) which react with lipids, proteins, pigments, and nucleic acids and causes lipid peroxidation, inactivation of enzymes, and damage to the membrane resulting in growth reduction [49]. The heavy metal copper is required in minute quantities by plants as it plays an important role in signaling of transcription, movement of proteins in cell, iron mobilization, oxidative phosphorylation and also acts as a co-factor for catalytic enzyme activity [50,51]. Excess of copper may result in the disruption of protein structure and function by binding to sulfhydryl groups due to its redox-active transition metal nature [51].
Differences in morphological and biochemical parameters were observed when they were treated with varying concentrations of CuSO4.5H2O and CdCl2. The effect of abiotic stress on the stevia plant showed that the physiological traits were more sensitive to salinity stress than the morphological traits. Application of CuSO4.5H2O and CdCl2 at lower concentrations (50 and 75 µM respectively) showed an increase in shoot length, number of leaves and leaf area as compared to control, while a reduction in all these parameters were observed with further increase in elicitors concentration. A similar increase in FW and DW was also recorded in both callus and leaves of treated plant at the same concentrations. A reduction the shoot length at higher salt concentrations have also been reported in bean, wheat, alfalfa and Zea mays [52-55]. Reduction in FW and DW at higher heavy metal concentrations may be referred to increased toxicity, thereby resulting in the breakdown normal physiological mechanisms and finally having negative influences on biomass [56].
In our study, chlorophyll content showed an increase at lower concentrations of both the heavy metals but as the concentration increased a decline was observed. In case of callus, an increase in chlorophyll content was observed up to 75 µM while for leaves up to 100 µM of CuSO4.5H2O. Similarly, in case of CdCl2, the increase was up to 50 µM and 75 µM respectively for callus and leaves. The leaves showed significant increase in chlorophyll content as compared to callus. In low concentrations, copper is not toxic but actually increases cell growth by becoming a cofactor for various growth enzymes. Copper is an important part of plastocyanin in the electron transport chain in photosynthetic reactions [57]. The copper (Cu) toxicity causes disruption of cell wall function by reducing the concentration of K+ ions in the cell and disrupt electron transport. The excess of copper will accumulate on the cell wall, absorbed into the cell and affect the enzyme by binding to the sulfhydryl group [58]. Similarly, excess cadmium disrupts the function of chloroplasts by damaging their photosynthetic pigments and inhibiting the photosynthetic activity. Without the photosynthesis process, the energy required for cell metabolic processes is disrupted which causes growth inhibition [59,60].
In our experiment, free radical scavenging activity through DPPH assay was expressed in terms of % inhibition. The results show that the leaves of stevia plant had the highest percentage inhibition at 75 µM/L cadmium chloride and 150 µM concentration for copper sulphate. In case of calli the higher % inhibition for copper sulphate was at 150 µM and at 100 µM for cadmium chloride, which was not very significantly different from control. The difference in chemical behaviors of cadmium and copper may be due to the fact that cadmium is a non-redox reactive heavy metal and copper is a transition metal [61]. Our results showed difference in antioxidant activity of callus and leaves which may be due to greater tolerance to cadmium stress as compared to copper in leaves. Previous studies have shown that intoxication by increased concentrations of Cd and Cu resulted in a decreased antioxidant capacity. This may be attributed to the accumulation of H2O2 resulting in growth reduction [61]. It can be concluded that the methanolic extract of Stevia plant leaves possessed strong antioxidant and free radical scavenging properties as compared to calli, which possibly may be due to the presence of its natural antioxidants.
Author Contributions
Afshan Shahzadi conducted most of the research work and drafted the manuscript. Ayishm Altaf did some part of work and analyzed the data. Humera Afrasiab supervised, reviewed, edited, and finalized the manuscript.
![]()
The authors declare that there is no conflict of interest.![]()
References
- Singh S, Rao G. Stevia: The herbal sugar of 21st century. Sugar tech, (2005); 7(1): 17-24.
- Akerele O. Nature's medicinal bounty: don't throw it away; 1993.
- Manandhar B. Ethnobiology of the Chepangs: A case study of Makwanpur District. Unpublished MSc dissertation, Kirtipur, Nepal: Tribhuvan University, (2000).
- Liu J, Li S. Separation and determination of Stevia sweeteners by capillary electrophoresis and high performance liquid chromatography. Journal of Liquid Chromatography & Related Technologies, (1995); 18(9): 1703-1719.
- Chalapathi M, Thimmegowda S, Sridhara S, Ramakrishna Parama V, Prasad T. Natural non-calorie sweetener stevia (Stevia rebaudiana Bertoni)-A future crop of India. CROP RESEARCH-HISAR-, (1997); 14347-350.
- Ahmed M, Salahin M, Karim R, Razvy M, Hannan M, et al. An efficient method for in vitro clonal propagation of a newly introduced sweetener plant (Stevia rebaudiana Bertoni.) in Bangladesh. American-Eurasian Journal of Scientific Research, (2007); 2(2): 121-125.
- Yoshida S. Studies on the Production of Sweet Substances in Stevia rebaudiana: 1. Simple determination of sweet glucosides in Stevia plant by thinlayer chromato-scanner and their accumulation patterns with plant growth. Japanese Journal of Crop Science, (1986); 55(2): 189-195.
- Chaiyana W, Charoensup W, Sriyab S, Punyoyai C, Neimkhum W. Herbal Extracts as Potential Antioxidant, Anti‐Aging, Anti‐Inflammatory, and Whitening Cosmeceutical Ingredients. Chemistry & Biodiversity, (2021); 18(7): e2100245.
- Jeppesen PB, Gregersen S, Alstrup K, Hermansen K. Stevioside induces antihyperglycaemic, insulinotropic and glucagonostatic effects in vivo: studies in the diabetic Goto-Kakizaki (GK) rats. Phytomedicine, (2002); 9(1): 9-14.
- Jeppesen P, Gregersen S, Rolfsen S, Jepsen M, Colombo M, et al. Antihyperglycemic and blood pressure-reducing effects of stevioside in the diabetic Goto-Kakizaki rat. Metabolism, (2003); 52(3): 372-378.
- Chan P, Xu D-Y, Liu J-C, Chen Y-J, Tomlinson B, et al. The effect of stevioside on blood pressure and plasma catecholamines in spontaneously hypertensive rats. Life sciences, (1998); 63(19): 1679-1684.
- Melis MS. Effects of chronic administration of Stevia rebaudiana on fertility in rats. Journal of Ethnopharmacology, (1999); 67(2): 157-161.
- Fujita H. Edahira. Safety utilization of Stevia sweetener. The Food Industry, (1979); 8265-72.
- Cerda-García-Rojas C, Pereda-Miranda R. The Genus Stevia. The phytochemistry of Stevia: A general survey; Taylor & Francis: New York, NY, USA, (2002); 86-118.
- Taware A, Mukadam D, Chavan A, Taware S. Comparative studies of in vitro and in vivo grown plants and callus of Stevia rebaudiana (Bertoni). International Journal of Integrative Biology, (2010); 9(1): 10-15.
- Hassanen SA, Khalil RM. Biotechnological studies for improving of stevia (Stevia rebaudiana Bertoni) in vitro plantlets. Middle-East Journal of Scientific Research, (2013); 14(1): 93-106.
- Murthy HN, Lee E-J, Paek K-Y. Production of secondary metabolites from cell and organ cultures: strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell, Tissue and Organ Culture (PCTOC), (2014); 118(1): 1-16.
- Yue W, Ming Q-l, Lin B, Rahman K, Zheng C-J, et al. Medicinal plant cell suspension cultures: pharmaceutical applications and high-yielding strategies for the desired secondary metabolites. Critical Reviews in Biotechnology, (2016); 36(2): 215-232.
- Hussain MS, Fareed S, Ansari S, Rahman MA, Ahmad IZ, et al. Current approaches toward production of secondary plant metabolites. Journal of Pharmacy & Bioallied Sciences, (2012); 4(1): 10.
- Ames BN, Shigenaga MK, Hagen TM. Oxidants, antioxidants, and the degenerative diseases of aging. Proceedings of the National Academy of Sciences, (1993); 90(17): 7915-7922.
- Harman D. Role of antioxidant nutrients in aging: overview. Age, (1995); 18(2): 51-62.
- Kanwar JR, Kanwar RK, Burrow H, Baratchi S. Recent advances on the roles of NO in cancer and chronic inflammatory disorders. Current Medicinal Chemistry, (2009); 16(19): 2373-2394.
- Chiavaroli V, Giannini C, De Marco S, Chiarelli F, Mohn A. Unbalanced oxidant–antioxidant status and its effects in pediatric diseases. Redox Report, (2011); 16(3): 101-107.
- Sharma V, Singh P, Pandey AK, Dhawan A. Induction of oxidative stress, DNA damage and apoptosis in mouse liver after sub-acute oral exposure to zinc oxide nanoparticles. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, (2012); 745(1-2): 84-91.
- Organization WH The state of food security and nutrition in the world 2019: safeguarding against economic slowdowns and downturns. Chapter: Book Name. 2019 of publication; 2019; Food & Agriculture Org.
- Singh S, Prasad SM. Growth, photosynthesis and oxidative responses of Solanum melongena L. seedlings to cadmium stress: mechanism of toxicity amelioration by kinetin. Scientia Horticulturae, (2014); 1761-10.
- Ghanta S, Banerjee A, Poddar A, Chattopadhyay S. Oxidative DNA damage preventive activity and antioxidant potential of Stevia rebaudiana (Bertoni) Bertoni, a natural sweetener. Journal of Agricultural and Food Chemistry, (2007); 55(26): 10962-10967.
- Skoog F, Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum, (1962); 15473-497.
- Arnon DI. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiology, (1949); 24(1): 1.
- Alabri THA, Al Musalami AHS, Hossain MA, Weli AM, Al-Riyami Q. Comparative study of phytochemical screening, antioxidant and antimicrobial capacities of fresh and dry leaves crude plant extracts of Datura metel L. Journal of King Saud University-Science, (2014); 26(3): 237-243.
- Nickavar B, Kamalinejad M, Haj-Yahya M, Shafaghi B. Comparison of the free radical scavenging activity of six Iranian Achillea. species. Pharmaceutical Biology, (2006); 44(3): 208-212.
- Maiti RK, Purohit S Stevia: A miracle plant for human health. Chapter: Book Name. 2008 of publication; Agrobios.
- Nakamura S, Tamura Y. Variation in the main glycosides of Stevia (Stevia rebaudiana Bertoni). Japanese Journal of Tropical Agriculture, (1985); 29(2): 109-115.
- Sakaguchi M, Kan T. Japanese research on Stevia rebaudiana (Bert.) Bertoni and stevioside. Ciencia e cultura-Sociedade Brasileira para o Progresso da Ciencia, (1982).
- Vyas SP, Dixit, V. K. In vitro culture on Stevia rebaudiana Bert. Pharmaceutical Biotechnology, (1999); 298-299.
- Sivaram L, Mukundan U. In vitro culture studies on Stevia rebaudiana. In vitro Cellular & Developmental Biology-Plant, (2003); 39(5): 520-523.
- Yücesan B, Mohammed A, Büyükgöçmen R, Altuğ C, Kavas Ö, et al. In vitro and ex vitro propagation of Stevia rebaudiana Bertoni with high Rebaudioside-A content—A commercial scale application. Scientia Horticulturae, (2016); 20320-28.
- Anbazhagan M, Kalpana M, Rajendran R, Natarajan V, Dhanavel D. In vitro production of Stevia rebaudiana Bertoni. Emirates Journal of Food and Agriculture, (2010); 216-222.
- Singh P, Dwivedi P, Atri N. In vitro shoot regeneration of Stevia rebaudiana through callus and nodal segments. International Journal of Agriculture, Environment and Biotechnology, (2012); 5(2): 101-108.
- Yang YW, Chang WC. In vitro plant regeneration from leaf explants of Stevia rebaudiana Bertoni. Zeitschrift für Pflanzenphysiologie, (1979); 93(4): 337-343.
- Pandey M, Chikara SK. In vitro regeneration and effect of abiotic stress on physiology and biochemical content of Stevia rebaudiana ‘Bertoni’. Journal of Plant Science and Research, (2014); 1(3): 113.
- Autade R, Fargade S, Borhade P, Udmale S, Choudhary R. In vitro propagation of Stevia rebaudiana (Bert.) A natural, non caloric sweetener herb. Journal of Cell and Tissue Research, (2014); 14(3): 4659.
- Mousumi D. Clonal propagation and antimicrobial activity of an endemic medicinal plant Stevia rebaudiana. Journal of Medicinal Plants Research, (2008); 2(2): 045-051.
- Patel R, Shah R. Regeneration of Stevia plant through callus culture. Indian Journal of Pharmaceutical Sciences, (2009); 71(1): 46.
- Keshvari T, Najaphy A, Kahrizi D, Zebarjadi A. Callus induction and somatic embryogenesis in Stevia rebaudiana Bertoni as a medicinal plant. Cellular and Molecular Biology, (2018); 64(2): 46-49.
- Jawad A, Naafe M. Effect of plant growth regulators on callus formation in Stevia rebaudiana. International Journal of Environmental Sciences & Natural Resources, (2018); 14(5): 120-125.
- Gupta P, Sharma S, Saxena S. Callusing in Stevia rebaudiana (natural sweetener) for steviol glycoside production. International Journal of Agricultural and Biological Sciences, (2010); 1(1): 30-34.
- DalCorso G, Farinati S, Maistri S, Furini A. How plants cope with cadmium: staking all on metabolism and gene expression. Journal of Integrative Plant Biology, (2008); 50(10): 1268-1280.
- Li FT, Qi JM, Zhang GY, Lin LH, Fang PP, et al. Effect of cadmium stress on the growth, antioxidative enzymes and lipid peroxidation in two kenaf (Hibiscus cannabinus L.) plant seedlings. Journal of Integrative Agriculture, (2013); 12(4): 610-620.
- Yruela I. Copper in plants. Brazilian Journal of Plant Physiology, (2005); 17145-156.
- Tanyolac D, Ekmekçi Y, Ünalan Ş. Changes in photochemical and antioxidant enzyme activities in maize (Zea mays L.) leaves exposed to excess copper. Chemosphere, (2007); 67(1): 89-98.
- Bhardwaj P, Chaturvedi AK, Prasad P. Effect of enhanced lead and cadmium in soil on physiological and biochemical attributes of Phaseolus vulgaris L. Nature and Science, (2009); 7(8): 63-75.
- Veselov D, Kudoyarova G, Symonyan M, Veselov S. Effect of cadmium on ion uptake, transpiration and cytokinin content in wheat seedlings. Bulgarian Journal of Plant Physiology, (2003); 29(3-4): 353-359.
- Aydinalp C, Marinova S. The effects of heavy metals on seed germination and plant growth on alfalfa plant (Medicago sativa). Bulgarian Journal of Agricultural Science, (2009); 15(4): 347-350.
- Mihalescu L, Mare-Rosca O, Marian M, Bildar C. Research on the growth intensity of the Zea mays L. plantlets aerial parts under cadmium treatment. Analele Universitatii din Oradea, Fascicula Biologie, (2010); 17(1): 147-151.
- Bahmani R, Bihamta M, Habibi D, Forozesh P, Ahmadvand S. Effect of cadmium chloride on growth parameters of different bean genotypes (Phaseolus vulgaris L.). ARPN Journal of Agriculture and Biological Science, (2012); 735-40.
- Arunakumara K, Zhang X. Heavy metal bioaccumulation and toxicity with special reference to microalgae. Journal of Ocean University of China, (2008); 7(1): 60-64.
- Suratno S, Puspitasari R, Purbonegoro T, Mansur D. Copper and cadmium toxicity to marine phytoplankton, Chaetoceros gracilis and Isochrysis sp. Indonesian Journal of Chemistry, (2015); 15(2): 172-178.
- Qian H, Li J, Sun L, Chen W, Sheng GD, et al. Combined effect of copper and cadmium on Chlorella vulgaris growth and photosynthesis-related gene transcription. Aquatic Toxicology, (2009); 94(1): 56-61.
- Hindarti D, Larasati A. Copper (Cu) and Cadmium (Cd) toxicity on growth, chlorophyll-a and carotenoid content of phytoplankton Nitzschia sp; (2019). IOP Publishing. pp. 012053.
- Schutzendubel A, Polle A. Plant responses to abiotic stresses: heavy metal‐induced oxidative stress and protection by mycorrhization. Journal of Experimental Botany, (2002); 53(372): 1351-1365.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0