

Full Length Research Article
Quantitative analysis of biofuel MET6 expression level in a treated Saccharomyces cerevisiae with garlic (Allium sativum L.) extract
Alawiah Mohammad Alhebshi, Nouf Khalid Al-Qahtani*
Adv. life sci., vol. 12, no. 2, pp. 304-311, May 2025
*- Corresponding Author: Nouf Khalid Al-Qahtani (Email: Nouf.qh2@gmail.com)
Authors' Affiliations
[Date Received: 20/10/2024; Date Revised: 05/01/2025; Date Published: 22/03/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Methionine and cysteine are the common sulfur-containing amino acids integrated into proteins. However, methionine is the immediate initiating amino acid in the biosynthesis of almost all eukaryotic proteins. Metabolism of methionine initiates with activating S adenosylmethionine, where this cofactor plays numerous roles in transfer methyl group pathways. There are various biosynthetic paths of methionine, in fungi, the ultimate step of methionine biosynthesis is motivated by cobalamin-independent methionine synthase (MET6). Chemically produced methionine is widely used in various applications. However, the issue is the sulfur source, which is typically supplied as inorganic sulfate, which needs to be reduced to create methionine.
Methods: Herein, testing the novel hypothesis that using natural resources such as garlic (Allium sativum), which is rich in organosulfur nitrogen-containing compounds, might enhance the biosynthesis of methionine. A quantitative analysis of MET6 mRNA was performed using qRT-PCR, eemploying RNA additions in all groups for treated Saccharomyces cerevisiae (WT-BY4742) and met15Δ mutant strains with and minus garlic extract.
Results: In this study, according to the Two-way ANOVA test, Met6 expression level was up-regulated significantly after 2 hours garlic exposure, and a significant increase in cells growth was observed of both mutant and wild-type strains compared with untreated control.
Conclusion: It has been concluded that garlic is a prominent natural source that may stimulate methionine biosynthesis, and this may owe to its organosulfur-containing compounds, such plant source could offer new natural approaches for methionine production, also the prevention and management of certain diseases in low cost and stable components.
Keywords: Organosulfur compounds; qRT-PCR; Saccharomyces cerevisiae; MET6 gene; Methionine biosynthesis; Garlic extract
Introduction![]()
Methionine is one of the vital sulfur-containing amino acids crucial for the metabolic processes in various organisms, both animal and plant. Despite most organisms rely on external sources for methionine, it can produce internally by plants. Beyond its role as a protein building block, methionine serves additional functions. It acts as a precursor to S-adenosylmethionine (SAM), which plays key roles in methyl-group transfer reactions, polyamine synthesis, and the production of the plant hormone ethylene [1]. Furthermore, a study of Walvekar et al [2] presented that the presence of methionine (even when other amino acids are limited) causes the production of all other amino acids and nucleotides needed to maintain anabolism where the generation of metabolites and co-factors is necessary for reductive biosynthesis forms and the basis of this anabolic program.
Many plants, fungi, and bacteria possess the ability to synthesize methionine from a variety of sources, including carbohydrates, as well as inorganic and organic sources of nitrogen with sulfur. In contrast, all animals, including humans, are reliant on dietary intake to obtain methionine. Currently, the industrial production of methionine primarily relies on chemosynthesis encompassing acrolein, methyl mercaptan, and hydrogen cyanide. This chemo-synthesized methionine is extensively utilized across various industries [1,3]. Yet, a key challenge in this process is the requirement for a sulfur origin, which is typically supplied as inorganic sulfate, which needs to be reduced to create methionine. Therefore, in current years, there has been rising delight in alternative and more sustainable processes that utilize natural resources[4,5].
Biofuels have been featured as a promising alternative to conventional fossil fuels, presenting potential solutions to environmental concerns and energy sustainability. Among various biofuel sources, Saccharomyces cerevisiae, a yeast species widely utilized in bioethanol production, stands out for its efficiency in fermenting sugars into ethanol. However, enhancing yeast performance through genetic modifications and bioactive compound treatments has garnered significant attention to improving biofuel production efficiency [6].
Garlic (Allium sativum L.), a member of the family Liliaceae, contains an abundance of chemical compounds encompassing water, carbohydrates, saponins, phenolic compounds, polysaccharides organosulfur compounds, proteins, free amino acids, Vitamins, minerals, and fiber (Figure 2.6) organic sulfides compounds are major bioactive components in garlic include allicin, alliin, diallyl sulfide, diallyl disulfide, diallyl trisulfide, ajoene, and S-allyl-cysteine, that have been shown to possess beneficial effects to protect against several diseases, and its play a lot of biological functions, such as antioxidant, anti-inflammatory, immunomodulatory, cardiovascular protective, anticancer, hepatoprotective, digestive system protective, anti-diabetic, anti-obesity, neuroprotective, renal protective, antibacterial, and antifungal activities[7,8,9].
This study aimed to explore the effectiveness of these extracts on enhancing the biosynthesis of methionine by comparing the expression level of the MET6 gene of a treated S. cerevisiae with extracts of garlic and with untreated (control) samples. Overall, this research will bridge the gap between molecular biology and biofuel, offering a comprehensive of the potential synergistic effects of natural extracts on gene expression for MET6, which may be able to enhance the MET6 in Saccharomyces cerevisiae, therefore, may lead to enhance cell viability, and thereby increase the biofuels production, making the process more efficient and potentially leading to more sustainable energy sources.
Methods![]()
Strains
Saccharomyces cerevisiae (wild-type strain BY4742) with genotype (MATa; his3∆1; leu2∆0; lys2∆0; ura3∆0) and the deletion-mutant (met15Δ- BY4741) (MATa; his3∆1; leu2∆0; met15∆0; ura3∆0) was obtained from Euroscarf (Frankfurt, Germany) [10].
Yeast extract peptone dextrose (YEPD) agar medium
Yeast extract peptone dextrose (YEPD) agar (3%yeast extract (w/v), peptic 5 % (w/v), agar 15% (w/v) (Microgen-NED) was prepared according to the manufacturing instructions: 23.0 g from YEPD was dissolved in 1 Liter of distilled water; the solution's pH was adjusted to 5.5.
Synthetic Complete (SC) medium
Based upon yeast nitrogen base without amino acids (YNB) (sodium chloride, magnesium sulfate, potassium sulfate, calcium chloride, ammonium sulfate (FORMEDIUM-UK), and a complete supplement mixture (CSM) containing the appropriate amino acids, but lacking methionine (FORMEDIUM- UK), synthetic complete (SC) medium was prepared according to the manufacturing instructions: 6,9 g from YNB, and 770 mg of (CSM) complete supplement mixture were dissolved in 1 Litter of distilled water, then pH was adjusted to 5.5.
Garlic extract preparation
Garlic cloves were obtained from a local market. To prepare garlic extract: 40 grams of garlic were peeled, washed with distilled water, and then crushed in a screw mill to extract juice. The garlic juice was then mixed with 200 ml of distilled water and filtered via sterile gauze to remove solids. For further purification this juice was transferred to centrifuge tube and precipitated via centrifuge at 4500 rpm for 30 min at 20 °C. A complete filtering yielded 200 ml of collected garlic extract for experimental use [11].
Yeast culture
growth conditions of yeast strains
The Saccharomyces cerevisiae strains BY4742 (WT) and the deletion-mutant (met15Δ- BY4741) were routinely maintained and cultured in (YEPD) broth medium, or YEPD agar, or in (YNB) medium (sodium chloride, magnesium sulfate, potassium sulfate, calcium chloride, ammonium sulfate (FORMEDIUM-UK) supplemented with a complete supplement mixture (CSM) which containing all amino acids, but lacking methionine (FORMEDIUM- UK). The cultures were incubated overnight at 30°C with shaking at 120 rpm until reaching the stationary phase in standard YEPD or selective YNB broth medium. The cultures were then diluted into fresh medium and allowed to grow for an additional 4-5 hours before being used in experiments [12].
All Saccharomyces cerevisiae strains were maintained at 4°C on appropriate agar plates for short-term storage. For long-term preservation, glycerol stocks were prepared with 20% (w/v) glycerol and stored at -80°C.
Growth curve assay in broth media
The SC medium, which consisted of (YNB) lacking all amino acids and supplemented with a complete supplement mixture that containing the optimal amino acids, but without methionine (CSM-MET), was used as described by Sherman [13]. For the cells' growth assay, single colonies were transferred from YEPD agar plate into 50 mL of SC medium using a 250 ml shake flask. Cultures were grown and incubated at 28°C with shaking (120 rev/min) overnight until they reached the stationary phase. Subsequently, samples were diluted to OD600~0.02 and tenfold diluted aliquots transferred to fresh SC medium and allowed to grow for further 4 hours to exponential phase at the same conditions before experimental use. For growth curve assays, cultures were divided equally into two groups:
Group I: BY4742 (WT) and the deletion-mutant (met15Δ- BY4741) cells were left without treatments as a control.
Group II: BY4742 (WT) and the deletion-mutant (met15Δ- BY4741) cells were treated with garlic extract at a final concentration of 10%.
All these cultures were incubated with shaking and the optical density (OD) at a wavelength of 600 nm (OD600) recorded continuously after 2 hours using a BioTek 16 Powerwave XS microplate spectrophotometer (BIO TEK SYNERGY 2 MICROPLATE READER, Thermo Scientific™, USA), as described Khozoie et al [14]. Three independent biological replicates for each group were performed for statistical analysis.
RNA extraction
RNA was extracted from S. cerevisiae of all groups using the RNeasy Mini Kit (Cat. No. 74104, Qiagen, Germany). All samples containing 1x cells were placed into sterile microfuge tubes and centrifuged for 10 minutes at 6000 rpm at 4 °C. Cell pellets were immediately placed and ground in liquid nitrogen with glass beads using a cooled pestle and mortar. After that, steps were followed as the manufacturer's instructions for the RNeasy mini kit.
RNA Purification
The RNA template was treated with DNase I to remove contaminating DNA using kit RNase-Free DNase (Cat. No.79254- Qiagen-DE). Briefly, the RNA was incubated with 10 μL of 1x DNase I Reaction Buffer, 1 μL of DNase I enzyme, and nuclease-free water to a final volume of 50 μL. The reaction was carried out at 37°C for 15 minutes. After DNase I treatment, RNA was purified using a phenol-chloroform extraction and ethanol precipitation method as described by Vomelova [15]. An equal volume (100 μL) of phenol-chloroform-isoamyl alcohol (25:24:1, pH 4.5-5) (Invitrogen™-USA) was added to the reaction mixture. The solution was mixed thoroughly by shaking and then centrifuged at 16,000 x g for 5 minutes at 4°C. The upper aqueous layer containing the RNA was carefully transferred to a clean tube and 10 μL of 2M sodium acetate (pH 5.2) (Invitrogen™-USA) and 400 μL of ice-cold 100% ethanol (Invitrogen™-USA) were added to the aqueous layer to precipitate the RNA. The solution was mixed gently by inversion and incubated at -80°C for 1 hour to allow for RNA precipitation. The mixture was centrifuged at 16,000 x g for 15 minutes at 4°C. The supernatant was carefully removed and discarded. The RNA pellet was washed once with 1 mL of 70% ethanol by centrifugation at 16,000 x g for 15 minutes at 4°C. The supernatant was discarded, and the remaining pellet was air-dried for 5-10 minutes. Finally, the purified RNA was resuspended in 25 μL of sterile, nuclease-free water for downstream applications. Then, RNA concentration and purity were measured by using (NanoDrop Thermo Sscientific, USA) and gel electrophoresis.
RNA Integrity Assessment Using Agarose Gel Electrophoresis
Dissolved agarose gels were used to separate nucleic acids based on size. These gels were prepared by heating agarose in 1x TAE buffer (RBI® Research Products International) In 1x TAE buffer (containing 242 g Tris-base, 200 ml of 0.5M EDTA pH 8.0, 57.1 ml CH3COOH, and water to the required volume), the solution was heated in a microwave for 1 minute and 40 seconds. After cooling, ethidium bromide was added to visualize the separated RNA. The gels were then placed in a specialized tray, electrophoresed in a tank, and loaded onto the gel. The electrophoreses were run for 40-60 min at 100 V, and the separated RNA was visualized using BioSpectrum Imaging Systems.
First-strand cDNA synthesis method
RNA, being a single-stranded molecule, cannot be amplified directly. To overcome this limitation, reverse transcription was performed to convert the RNA into double-stranded DNA, making it amenable to amplification. The reverse transcription reaction was carried out using a full RT kit (CT.NO. A5001, Promega GoScript® Reverse Transcription System-USA), and a master mix was prepared. The master mix was prepared in a biosafety cabinet (BSC) to minimize cross-contamination. To maintain the integrity of the RT reagents, which are temperature-sensitive, they were kept on ice until ready for use. Reverse transcription reactions were set up using Reverse Transcriptase (RT), deoxynucleotide (dNTP) solution mix (10 mM each dATP, dCTP, dTTP, dGTP), and Oligo (dT) Primer. The amount of master mix made was determined by the number of samples analyzed and according to the manufacturer’s instructions.
Quantitative RT-PCR (qRT-PCR)
Real-time PCR was performed using an Applied Biosystems™ StepOne™ Real-Time PCR System machine where performed in three independent reactions, each comprising 100ng purified cDNA, 100nM MET6, and ACT1 -specific primers, a 20 µL reaction containing 2 μL of first-strand cDNA, 10 µL of master mix qPCR Kit (EverGreen, HavenSci, KSA), 1 µL of 10M F primer, 1 µL of 10M R primer, and 6 of nuclease-free ddH2O. MET6 gene-specific primers and ACT1 housekeeping gene primers were selected as a reference for measuring the amount of total RNA in the sample. The optimal annealing temperature was determined based on Taq DNA polymerase's gradient PCR reactions. The expression of the housekeeping control gene ACT1 and the data were analyzed by the 2(-Delta Delta C(T)) method [16].
Statistical Analysis
Statistical analysis of the data was conducted using two-way ANOVA with Prism 10 software to determine whether differences were remarkable statistically significant. Statistical significance was considered when the p-value was below 0.05 (p < 0.05), while p-values greater than or equal to 0.05 (p ≥ 0.05) indicated no significant difference.
Results![]()
Morphological characteristics of yeast S. cerevisiae cells on YEPD agar medium
For the morphological study, the wild-type strain of S. cerevisiae (BY4742-WT) and the mutant strain met15Δ(BY4741) were prepared and tested for growth in YEPD medium firstly. Morphological description of yeast colonies showed that the shape of colonies of both yeast cells of S. cerevisiae (BY4742-WT) and the mutant strain met15Δ(BY4741) were round, creamy in color, and had smooth surfaces. However, the sizes of both cells types vary considerably Figure (1).
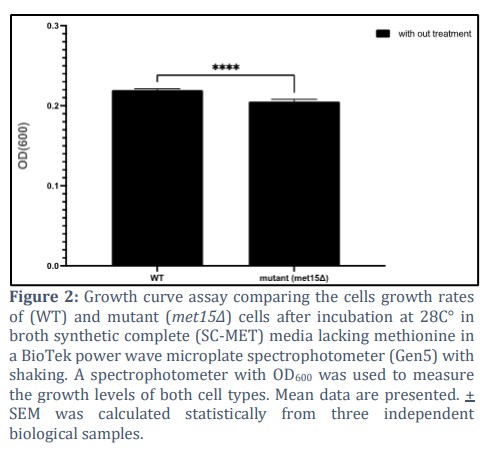
Absence of methionine decreases the growth rates of the deletion mutant cells (met15Δ) compared to the wild type cells
In order to compare the growth level of the BY4742 wild-type cells and deletion-mutant cells (met15Δ), spectrophotometer at OD600 was used to measure the growth level of the WT-BY4742 and the mutant cells (met15Δ) in the broth synthetic complete (SC) media, which is YNB medium supplemented with all amino acids except methionine (YNB-MET), for 2hr. The results indicated a significant increase in wild-type cells growth compared to the mutant strain met15Δ as shown Figure (2), and such differences were found to be remarkable statistically significant (p < 0.0072) according to the two-way ANOVA statistical analysis test. This rendered these cells suitable for testing whether the expression level of MET15 influences yeast cell growth.
Garlic extract enhances the growth rates of both wild type and mutant met15 Δ cells
The powerful biological advantages of garlic cloves are related mainly to its organosulfur components which act as a source of sulfur, which is an essential source for growth and used primarily to synthesis methionine. For this reason, the organosulfur molecules are the leading candidate for the garlic extract being investigated [7,17]. Therefore, the same assay that was mentioned above but with adding100 μl/ml of garlic extract in the SC (YNB-MET) media lacking methionine was used for both BY4742 wild-type cells and the deletion-mutant cells (met15Δ- BY4741) to test whether garlic extract elevate cells growth of both types. A present result showed that after the exposure to 100 μl/ml of garlic extract, an obvious improvement of cells growth rate was observed for both types of cells compared to the untreated strains (control) Figure (3). Furthermore, this increase in cells growth level of a treated wild type and met15 Δ cells was significant compared to the control according to the two-way ANOVA statistical analysis test where (p < 0.0001) figure (3 A). Moreover, there was a slight increase in growth levels of the treated met15 Δ cells compared to the untreated wild type cells but these differences are not significant statistically figure (3 B).
These data may elucidate the role of organosulfur components of garlic extract in compensating the lack of methionine or may act as a synergistic factor for methionine biosynthesis, which is an essential amino acid for cells growth, therefore, quantification of MET 6 gene expression level in both strains types (treated-untreated) and under the same conditions was required to verify whether garlic extract upregulate methionine expression levels or may act as synergistic for methionine biosynthesis, which may elucidate the cause of observed rise in growth level of both cell types.
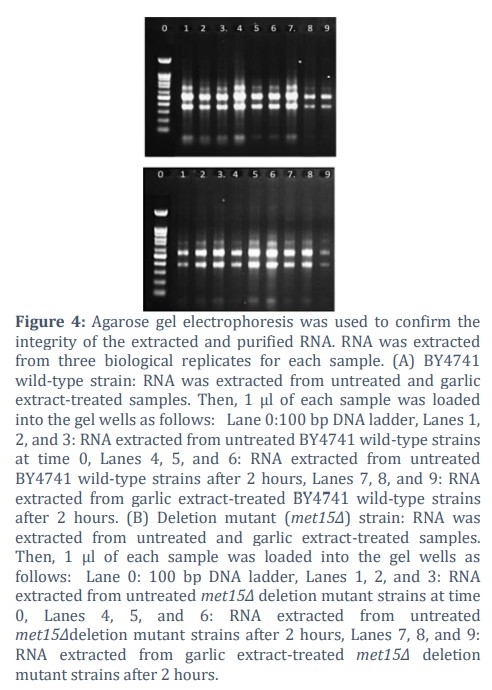
Quantification of purified RNA concentrations using a Nano-Drop spectrophotometer and gel electrophoresis.
The extraction of a high-quality RNA from both wild type and the deletion mutant met15Δ cells is a critical step in further gene expression analysis. The consistent RNA yield and concentration across all samples indicate the reliability of the extraction method used. All extracted and purified RNA samples were quantified using a NanoDrop spectrophotometer (NanoDrop Technologies), results showed that the average of RNA concentration in both BY4742 (WT) and the (met15Δ) cells untreated (C) and treated with garlic (G) was approximately range between 100 to 200 ng/µl, with a minimum variation between the biological replicates. Furthermore, agarose gel electrophoresis was used to confirm these extracted and purified RNA concentrations that were determined by Nano-drop. All samples untreated and treated with garlic extracts were analyzed by agarose gel electrophoresis and results showed the integrity of the extracted RNA in all samples, as evidenced by the presence of clear RNA bands and no degradation of RNA was observed, which indicating that a high-quality RNA extraction (Figure 4). This yielded RNA is qualified now to be used as a template to synthesis the first-strand cDNA and subsequently used for (q RT-PCR).
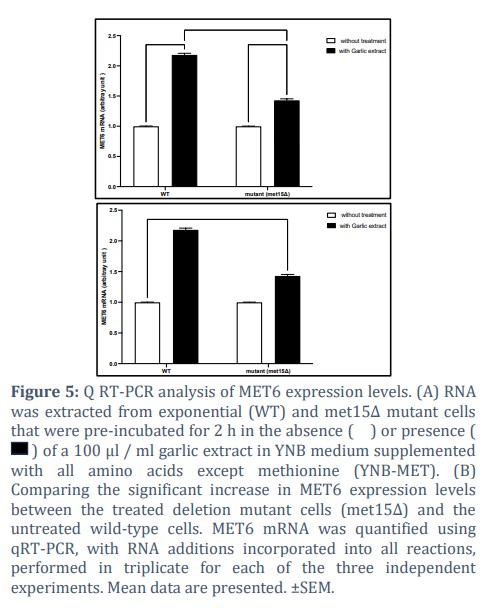
The plant extract alters the MET6 expression levels of the (WT) and (m. et15Δ) cells
Quantification analysis of MET6 gene expression levels in both the untreated and treated WT (BY4742) and (met15Δ) cells using q RT-PCR was performed in triplicate reactions, each comprising 100 ng purified cDNA to verify whether garlic extract affect MET6 expression level, and consequently may elucidate the cause of raising treated cells growth rates that showed previously.
Garlic extract upregulates MET6 expression level of both the wild type and the mutant met15Δ cells.
As mentioned previously methionine is an essential amino acid for cells growth [18].
Moreover, the powerful biological advantages of garlic cloves are related mainly to its organosulfur components which act as a source of sulfur, which is an essential source for growth and used primarily to biosynthesis methionine [7,17].
Therefore, to test whether garlic extract may elevate Met6 expression level and thereby may elucidate the cause of increasing cells growth rates of the treated wild-type (BY4742) and the deletion-mutant cells (met15Δ) that showed previously (figure 3) in media lacking methionine (YNB-MET), quantification analysis of MET6 gene expression levels of both the untreated and treated wild-type (BY4742) and the deletion-mutant cells (met15Δ) using q RT-PCR was performed in triplicate reactions, each comprising 100 ng purified cDNA.
Results showed that the addition of garlic extract led to elevating MET6 gene expression levels considerably of both treated wild-type (BY4742) and the deletion-mutant cells (met15Δ) compared to their untreated cells (control), and these differences were statistically significant according to the two-way ANOVA test where p <0.0001. Figure (5A).
Furthermore, the MET6 expression level was upregulated substantially in the treated deletion mutant cells (met15Δ) compared to the untreated (WT) cells, and these observed differences were found to be statistically significant according to the two-way ANOVA test where p<0.0001. Figure (5 B).
According to these results, the reason of increasing cells growth levels that were observed previously of both treated strains in media lacking methionine may owing to the importance of organosulfur components of garlic extract, which act as a source of sulphur that may act as a synergistic factor for methionine biosynthesis and thereby compensating its deficiency.
Figures & Tables


biosynthesis of almost all eukaryotic proteins. Saccharomyces cerevisiae Meth6 is described as cobalamin-independent methionine synthase, which involved in methionine regeneration and biosynthesis. In fungi, the ultimate step of methionine biosynthesis is motivated by Meth6 [19]. Chemically produced methionine is widely used in various applications. However, an issue is the sulfur source, which is typically supplied as inorganic sulfate, which needs to be reduced to create methionine. Therefore, stimulating the production of cobalamin-independent methionine synthase (Meth6) using natural sources rich with organosulfur-and nitrogen-containing compounds, such as garlic (Allium sativum), was the aim of this study. Recent study showed that absence of MET15 gene in BY4741strain affected specifically sulphur metabolism, as well as sulphur absorption pathways of the mutant (met15Δ-BY4741) strain was altered and led to differences in cells growth rates between these strains of wild type and its isogenic mutant (met15Δ-BY4741) [20].
In addition, further study demonstrated that MET6 gene plays an important role in sulphur metabolism pathways next MET15. Its expressed methionine acts as precursor for all proteins biosynthesis where is reported as the first amino acid starts translation process [21,22]. As a result of all previous mentioned studies, wild type of S. cerevisiae (BY4742) and the deletion mutant (met15Δ- BY4741) strains were employed in this study to quantify MET6 expression levels in the absence and presence of natural sources such as garlic (Allium sativum), which are rich in organosulfur-and nitrogen-containing compounds, to verify their role in enhancing the methionine biosynthesis and cellular viability. The results of this study demonstrated a significant increase in the growth rate of untreated wild-type cells compared to the met15Δ deletion mutant strain. These differences were remarkable statistically significant, as determined by two-way ANOVA analysis, which is consistent with previous study that showed absence of methionine led to impaired metabolic processes, reduced glutathione synthesis, which acts as an antioxidant factor, and thereby lower cellular viability was observed, since cells became more vulnerable to oxidative stress [23]. Interestingly, our quantities analysis results of MET6 gene expression level using q RT-PCR showed that the addition of garlic extract led to elevating MET6 gene expression level considerably of both treated wild-type (BY4742) and the deletion-mutant cells (met15Δ) compared to the untreated both cells types (control), and these differences were statistically significant, following that an obvious improvement of cells growth rates was observed for both cells types compared to the untreated cells (control). These findings are consistent with Yu et al [24] study that used microarray analysis to demonstrate that allicin is the main active compound of garlic that led to significant expression levels increase of several methionine genes (MET3, MET10, MET14, MET16, MET17, and MET22) of Saccharomyces cerevisiae involved in sulphur assimilation and methionine biosynthesis pathways, but no earlier studies about MET6 gene expression levels in the absence and presence of aqueous garlic extracts have been founded using q RT-PCR. In addition, sulphur compounds of garlic extract are thought to compensate methionine deficiency. Moreover, [25] demonstrated that allicin treatment triggers the oxidative stress defence. Because it affects the expression level of genes associated with antioxidant activity, such as HMOX1, GCLC, and SLC6A6 in humans and thereby help improve metabolic regulation. The anticancer potential of aqueous garlic extracts was studied against human breast cancer cells (CAL-51) and mouse mammary adenocarcinoma cells (AMN-3) which showed that garlic's organosulfur compounds such as diallyl trisulfide and ajoene are thought to have anticancer properties since they affect signalling pathways linked to cell cycle progression, death, and cells proliferation. These compounds able to reduce CDK1 expression level in cancer cells, which results in cell cycle arrest and cell death reduction [26].
Author Contributions
Alawiah M. Alhebshi designed the study, performed the experimental analyses, wrote, revised, and edited the manuscript and gene expression testing. NOUF K. ALQAHTANI performed the experiments, participated in manuscript editing and performed the experimental analyses.
The authors declare that there is no conflict of interest regarding the publication of this paper.![]()
References
- Ravanel S, Gakiere B, Job D, Douce R. The specific features of methionine biosynthesis and metabolism in plants. Proceedings of the National Academy of Sciences, (1998); 95(13): 7805-7812.
- Walvekar AS, Srinivasan R, Gupta R, Laxman S. Methionine coordinates a hierarchically organized anabolic program enabling proliferation. Molecular Biology of the Cell, (2018); 29(26): 3183-3200.
- Lemme A, Naranjo V, de Paula Dorigam JC. Utilization of methionine sources for growth and Met+ Cys deposition in broilers. Animals, (2020); 10(12): 2240.
- Becker J, Wittmann C. Systems and synthetic metabolic engineering for amino acid production–the heartbeat of industrial strain development. Current Opinion in Biotechnology, (2012); 23(5): 718-726.
- Stipanuk MH, Ueki I. Dealing with methionine/homocysteine sulfur: cysteine metabolism to taurine and inorganic sulfur. Journal of Inherited Metabolic Disease, (2011); 34: 17-32.
- Sharma J, Kumar V, Prasad R, Gaur NA. Engineering of Saccharomyces cerevisiae as a consolidated bioprocessing host to produce cellulosic ethanol: Recent advancements and current challenges. Biotechnology Advances, (2022); 56: 107925.
- Omar SH, Al-Wabel NA. Organosulfur compounds and possible mechanism of garlic in cancer. Saudi Pharmaceutical Journal, (2010); 18(1): 51-58.
- Butt MS, Sultan MT, Butt MS, Iqbal J. Garlic: nature's protection against physiological threats. Critical Reviews in Food Science and Nutrition, (2009); 49(6): 538-551.
- Yousif HM, El Mahdy AM, Hassan MF, Mansour MK. Overview on Antioxidant and Oxidative Stress Markers after Garlic Oil Supplement in Suckling Buffalo Calves. Kafkas Universitesi Veteriner Fakultesi Dergisi. (2024); 30: 73-80.
- Baker Brachmann C, Davies A, Cost GJ, Caputo E, Li J, et al. Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR‐mediated gene disruption and other applications. Yeast, (1998); 14(2): 115-132.
- Yadav S, Trivedi NA, Bhatt JD. Antimicrobial activity of fresh garlic juice: An: in vitro: study. AYU (An International Quarterly Journal of Research in Ayurveda), (2015); 36(2): 203-207.
- Holland SL, Avery SV. Actin-mediated endocytosis limits intracellular Cr accumulation and Cr toxicity during chromate stress. Toxicological Sciences, (2009); 111(2): 437-446.
- Sherman F. Getting Started with Yeast. Methods in Enzymology, (2002); 350: 3-41.
- Khozoie C, Pleass RJ, Avery SV. The antimalarial drug quinine disrupts Tat2p-mediated tryptophan transport and causes tryptophan starvation. Journal of Biological Chemistry, (2009); 284(27): 17968-17974.
- Vomelova I, Vanickova Z, Sedo A. Technical note methods of RNA purification. All ways (should) lead to Rome. Folia Biologica (Praha), (2009); 55: 243-251.
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods, (2001); 25(4): 402-408.
- Amagase H. Clarifying the real bioactive constituents of garlic. The Journal of Nutrition, (2006); 136(3): 716S-725S.
- Ghazi T, Arumugam T, Foolchand A, Chuturgoon AA. The impact of natural dietary compounds and food-borne mycotoxins on DNA methylation and cancer. Cells, (2020); 9(9): 2004.
- Sadhu MJ, Moresco JJ, Zimmer AD, Yates III JR, Rine J. Multiple inputs control sulfur-containing amino acid synthesis in Saccharomyces cerevisiae. Molecular Biology of the Cell, (2014); 25(10): 1653-1655.
- Van Oss SB, Parikh SB, Coelho NC, Wacholder A, Belashov I, et al. On the illusion of auxotrophy: met15Δ yeast cells can grow on inorganic sulfur, thanks to the previously uncharacterized homocysteine synthase Yll058w. Journal of Biological Chemistry, (2022); 298(12): 102697.
- Stipanuk MH. Metabolism of sulfur-containing amino acids: how the body copes with excess methionine, cysteine, and sulfide. The Journal of Nutrition, (2020); 150: 2494S-2505S.
- Lauinger L, Kaiser P. Sensing and signaling of methionine metabolism. Metabolites, (2021); 11(2): 83.
- Walvekar AS, Laxman S. Methionine at the heart of anabolism and signaling: perspectives from budding yeast. Frontiers in Microbiology, (2019); 10: 2624.
- Yu L, Guo N, Meng R, Liu B, Tang X, et al. Allicin-induced global gene expression profile of Saccharomyces cerevisiae. Applied Microbiology and Biotechnology, (2010); 88: 219-229.
- Lv C, Wang C, Li P, Huang Y, Lu X, et al. Effect of Garlic Organic Sulfides on Gene Expression Profiling in HepG2 Cells and Its Biological Function Analysis by Ingenuity Pathway Analysis System and Bio‐Plex‐Based Assays. Mediators of Inflammation, (2021); 2021(1): 7681252.
- Mohammed NT. Antiproliferative effect of garlic extract on cancer cell lines. Journal of Animal Health and Production, (2024); 12(3): 444-449.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


