

Full Length Research Article
Anti-atherosclerotic potential of fatty acids in Chlorella vulgaris via inhibiting the foam cell formation: An in silico study
Muhammad Hermawan Widyananda1,2, Rahmat Grahadi1,3, Ichda Arini Dinana1, Arif Nur Muhammad Ansori2,4,5, Viol Dhea Kharisma2, Vikash Jakhmola4, Maksim Rebezov6,7, Marina Derkho8, Pavel Burkov9, Pavel Scherbakov10, Rahadian Zainul11,12*
Adv. life sci., vol. 12, no. 2, pp. 296-303, May 2025
*- Corresponding Author: Rahadian Zainul (Email: rahadianzmsiphd@fmipa.unp.ac.id)
Authors' Affiliations
2. Drug and Vaccine Innovation Research Group, Virtual Research Center for Bioinformatics and Biotechnology, Surabaya – Indonesia.
3. Research Center of Smart Molecule of Natural Genetics Resources, Brawijaya University, Malang – Indonesia.
4. Uttaranchal Institute of Pharmaceutical Sciences, Uttaranchal University, Dehradun – India.
5. Division of Research and Development, Jalan Tengah, Surabaya – Indonesia.
6. Faculty of Biotechnology and Food Engineering, Ural State Agrarian University, Yekaterinburg – Russia.
7. K.G. Razumovsky Moscow State University of Technologies and Management (The First Cossack University), Moscow – Russia.
8. Department of Natural Sciences, South Ural State Agrarian University, Troitsk – Russia.
9. Center for Biotechnology of Animal Reproduction, South Ural State Agrarian University, Troitsk – Russia.
10. Department of Infectious Diseases and Veterinary, South Ural State Agrarian University, Troitsk – Russia.
11. Department of Chemistry, Faculty of Mathematics and Natural Sciences, Universitas Negeri Padang, Padang – Indonesia.
12. Center for Advanced Material Processing, Artificial Intelligence, and Biophysic Informatics (CAMPBIOTICS), Universitas Negeri Padang, Padang – Indonesia
[Date Received: 20/10/2024; Date Revised: 05/01/2025; Date Published: 22/03/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: WHO in 2019 announced that atherosclerosis is included in the top leading cause of mortality, which is more than 32% of death worldwide. The most common early symptom of atherosclerosis is the formation of foam cells inside blood vessel wall. Fatty acids from Chlorella vulgaris have the potential to inhibit foam cell formation. The purpose of this study is to analyze the inhibitory activity of fatty acids from C. vulgaris towards various proteins involved in foam cell formation.
Methods: The fatty acids content of C. vulgaris was determined based on previous research. The molecular samples of fatty acids and proteins including CETP, LOX1, ACAT1, and CD36 were obtained from the PubChem and RCSB PDB databases. Drug-likeness and probable activity screening were conducted using the SWISS ADME and PASS Online web servers. Molecular docking is performed using AutoDock Vina, which is integrated into the PyRx software.
Results: All fatty acids comply with the Lipinski rule of five. Thirteen fatty acids were predicted to have anti-atherosclerosis activity based on PASS online screening: myristoleic acid, hexadecadienoic acid, linolenic acid, palmitoleic acid, linoleic acid, heptadecenoic acid, oleic acid eicosadienoic acid, nonadecenoic acid, gadoleic acid, heneicosanoic acid, brassidic acid, and nervonic acid. Molecular docking simulation results showed that Nervonic acid bind to CETP, ALOX1, and ACAT1 in the same site as an inhibitor with the lowest binding affinity value (-5.9, -7, and -7.8 kcal/mol). Hexadecadienoic acid binds to CD36 with the lowest binding affinity value (-4.9 kcal/mol) and binds in the same site with inhibitor.
Conclusion: Therefore, nervonic acid and hexadecadienoic acid have a high potential as a foam cell formation inhibitor.
Keywords: Atherosclerosis; Fatty acid; Foam cell; Molecular docking
Introduction![]()
Atherosclerosis is considered a chronic cardiovascular disease (CVD) characterized by the retention of low-density lipoprotein (LDL) and inflammation that narrows and triggers damage to the inner walls of blood vessels [1,2]. In 2019, WHO included atherosclerosis in the top leading causes of mortality, accounting for more than 32% of deaths worldwide. The most common early symptom of atherosclerosis is the foam cells formation from macrophages in the atherosclerotic lesion of blood vessels walls [2,3]. Excessive consumption of high-fat-diet triggers high LDL content in cells. Reactive Oxygen Species (ROS) then modifies LDL to oxidized LDL (ox-LDL) [4]. The ox-LDL is captured by scavenger receptors (SRs) on the surface of cell membranes namely CD36 [1,2,5]. Ox-LDL will then enter the late endosome lysosome and degraded by lysosomal acid lipase (LAL) into free cholesterol (FC). FC can be re-esterified by acyl-coenzyme A: cholesterol acyltransferase-1 (ACAT1) into cholesterol esters (CE) [2,6]. This process triggers the CE build-up and the formation of droplets within the macrophage which initiates the formation of foam cells promoting the atherosclerotic lesion. FC-added HDL can be converted into LDL by Cholesterol Ester Transfer Protein (CETP), unlocking a new risk of ox-LDL formation, an endless cycle of atherosclerosis [2]. Despite the advancement in medical research, atherosclerosis remains a serious social and medical issue with limited drug options. Synthetic drug usage for atherosclerosis prevention is not a preferred treatment since it is considered expensive and high risk due to the finite number of indications for severe side effects [7]. Hence, anti-atherosclerotic treatment based on the natural product will be the best alternative.
The fatty acid has long been studied for its ability to inhibit foam cell formation. The green algae from the genus Chlorella have long been renowned as an essential fatty acid source, specifically C. vulgaris. Many researchers even considered it as a potent biofactory for fatty acid and other beneficial nutrients [8–10]. C. vulgaris can easily be found anywhere. It has high adaptability and photosynthetic capability, promoting rapid growth under various environmental conditions [11]. C. vulgaris fatty acid comprises of 15 saturated fatty acids (SFAs), 8 monounsaturated FAs (MUFAs), and 4 polyunsaturated fatty acids (PUFAs), providing it with a high lipid profile variability [12]. According to [13], fatty acid intake can reduce the ability of LDL to bind to LDL receptors expressed on the macrophage cell surface, thus preventing cholesterol accumulation. In addition, [14] specifically describe the role of omega-3 fatty acid as an anti-atherosclerotic compound that alters monocyte subsets and restricts monocyte intake to the aortic lesion. In the former research, C. vulgaris supplementation resulted in decreasing the rate of cholesterol absorption by promoting bile acid turnover serving anti-atherosclerosis activity [15]. Regardless of all research proposing anti-atherosclerosis properties of C. vulgaris, the mechanism underlying inhibition foam cell development prevention by this organism is complex and not well defined, though, it is known that foam cell is an atherosclerotic hallmark.
Advancement in technology eases the access for researchers to predict specific biological mechanisms through software only. This method is known as in silico procedure, which help to make decision or virtually simulate every bit of drug discovery and development [16]. This method is inexpensive and time effective. Furthermore, drug designing tool provides a high accuracy in decreasing the fatal risk of drug administration [17]. Currently, there are no direct methods to prevent atherosclerosis. Even though an anti-atherosclerotic treatment does exist, those approach are still under development [18]. Even Statins, the first-line treatment for atherosclerotic patients are not free from adverse effects [7]. For those reasons, exploring the fatty acid potential as anti-atherosclerotic compound by preventing foam cell formation is crucial. This present work aimed to identify fatty acid from C. vulgaris and its potential inhibitory interaction with various proteins involved in foam cell formation of atherosclerosis pathogenesis using in silico approach.
Methods![]()
Sample retrieval
The total of 28 fatty acids content of C. vulgaris were retrieved from [19]. The 3D models of the fatty acids were obtained from PubChem (https://pubchem.ncbi.nlm.nih.gov/). The 4 proteins that responsible for forming foam cells such as CETP, LOX1, CD36, and ACAT1 were downloaded from protein data bank (https://www.rcsb.org/).
Druglikeness prediction
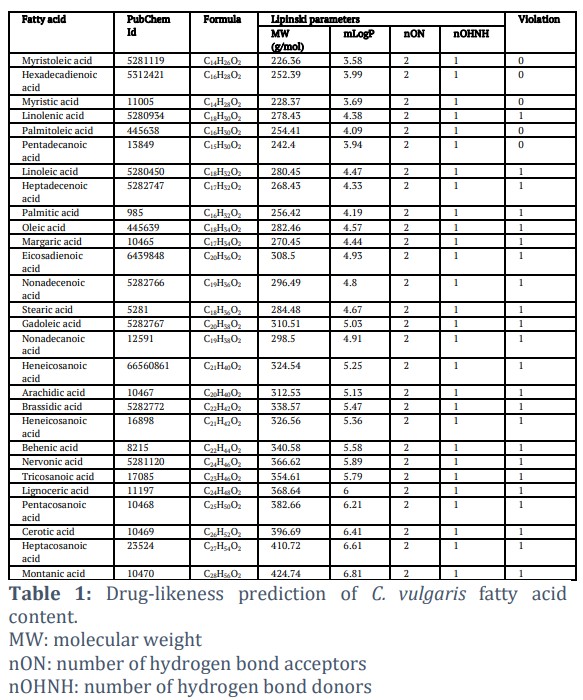
Druglikeness prediction was conducted using SwissADME (http://www.swissadme.ch/), which predicts the target compounds biological activity and toxicity in ADME (absorption, distribution, metabolism, and excretion) mechanisms. We use Lipinski’s parameter, which examines the physiochemistry of active compound when taken orally [20]. The violation, bioavailability, and toxicity of the compounds were also considered.
Bioactivity prediction
We predict the bioactivity of 28 fatty acids based on their structure formula using PASS online (http://way2drug.com/passonline/predict.php) [21]. PASS online determines the resemblance of the bioactivity by comparing the compounds’ structure-activity relationships with other identical compounds that the bioactivity and the toxicity are known. All listed fatty acids that have anti-inflammation, cholesterol antagonist, and anti-cholesterol parameters with Pa value of 0.6 were analyzed afterwards. The Pa value indicates the similarity of the compound tested to other known compounds regarding its structure and activity. We chose the Pa value of 0.6 because the higher Pa value tends to be analogous to the known drugs, while the lower Pa value the bioactivity of the compound is also lower.
Molecular docking and dynamic simulation
The 3D model of fatty acids and controls were prepared for ligands by minimizing energy using PyRx v0.8 [22] while the proteins (CETP, LOX1, CD36 and ACAT1) were prepared by removing the water molecules and natural ligands using Discovery Studio 2019 [23][24]. The proteins and ligands docking were conducted using Autodock Vina in PyRx v.0.8, a software with specific docking method. The grid position for the macromolecules CETP, LOX1, ACAT1, and CD36 were characterized by specific dimensions in the X, Y, and Z axes. For CETP, the coordinates were measured at X: 10.6905 (15.7386 Å), Y: 1.1872 (16.3630 Å), and Z: 37.3356 (15.6031 Å). LOX1 had search spaces defined as X: 25.8932 (44.7324 Å), Y: -30.4209 (47.3701 Å), and Z: -44.1985 (45.9076 Å). The macromolecule ACAT1 exhibited larger search spaces, with X: 100.4097 (21.9748 Å), Y: 163.9852 (21.9512 Å), and Z: 149.3645 (17.3379 Å). Lastly, the search space for CD36 was observed as X: -28.9857 (27.6564 Å), Y: -38.7696 (28.1075 Å), and Z: 54.6805 (24.8583 Å). Autodock vina uses stochastic global optimization to predict the free binding energy of the macromolecules and ligands, and the specific docking used to speed up the processes [25].
Molecular dynamics simulations were performed online using the CABS-flex 2.0 webserver (http://biocomp.chem.uw.edu.pl/CABSflex2). Determination of molecular dynamics parameters follows the default settings. This simulation was used to predict the stability of protein-ligand complex, RMSF value between 1-3 indicates stable amino acid during simulation [26].
Results![]()
Druglikeness Prediction Analysis
Druglikeness prediction was performed to eliminate fatty acids that do not have drug-like characteristics. Compounds with drug-like characteristics will be easily absorbed by the body, circulated by the blood, and reached the target protein [27]. Druglikeness prediction results showed that all fatty acids contained in C. vulgaris have drug-like characteristics because they meet the Lipinski rule of five (Table 1). According to Lipinski's rule, no more than two vibrations indicate the compound's drug-like character [28]. From these results, there are even fatty acids that do not have violation, namely Myristoleic acid, Hexadecadienoic acid, Myristic acid, Palmitoleic acid, and Pentadecanoic acid.
Bioactivity Prediction Analysis
Bioactivity prediction was carried out to analyze the possible bioactivity of fatty acid activity from C. vulgaris. This prediction used the PASS Online web server, which uses the QSAR principle, which predicts the compounds activity based on the structure of other known compounds [29]. The results showed that 13 fatty acids had a Pa value above 0.6 in the three parameters. Therefore, these thirteen compounds are predicted to have high anti-foam cell formation activity (Figure 1A). The thirteen compounds are Myristoleic acid, Hexadecadienoic acid, Linolenic acid, Palmitoleic acid, Linoleic acid, Heptadecenoic acid, Oleic acid Eicosadienoic acid, Nonadecenoic acid, Gadoleic acid, Heneicosanoic acid, Brassidic acid, and Nervonic acid (Figure 1B).
Molecular Docking and Simulation
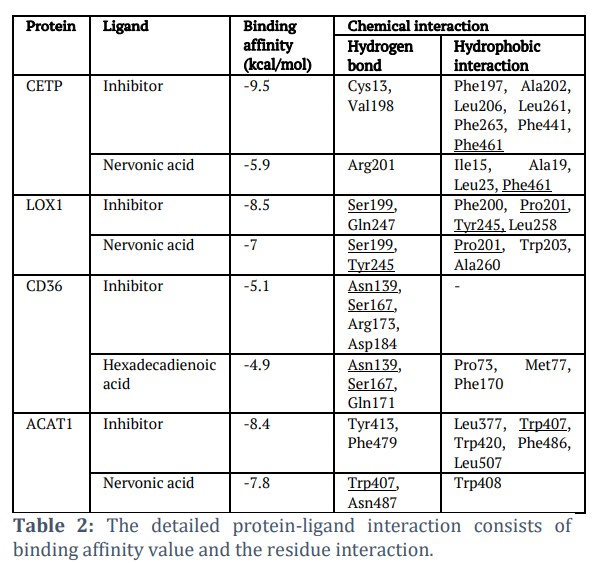
Molecular docking was carried out to analyze the potential of thirteen fatty acids from C. vulgaris in inhibiting four proteins related to foam cell formation, specifically CETP, LOX1, CD36, and ACAT1. The docking results show that thirteen fatty acids bind to the active site of those proteins (Figure 2A), indicating the potential inhibitory activity of those thirteen compounds. Out of the thirteen analyzed compounds, one compound has the lowest binding affinity value for each protein, which is predicted to have the best inhibitory ability to these proteins. The fatty acid with the lowest binding affinity value on the CETP, LOX, and ACAT1 proteins is Nervonic acid (-5.9, -7, dan -7.8 kcal/mol), while the fatty acid with the lowest binding affinity value on CD36 protein is Hexadecadienoic acid (-4.9 kcal/mol) (Table 2).
Molecular Interaction Analysis
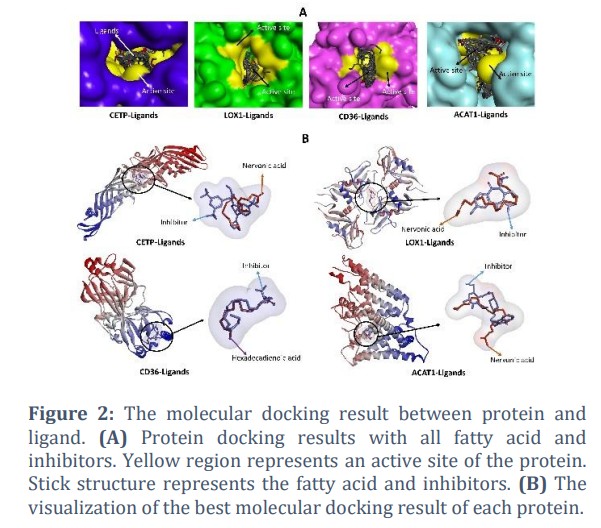
All fatty acids with the highest binding affinity value are bind in the same cavity as the inhibitor (Figure 2B). The details of the interactions are shown in Table 2. Nervonic acid binds CETP by forming one hydrogen bond (Arg201) and four hydrophobic interactions (Ile15, Ala19. Leu23, and Phe461). Nervonic acid is bound to the same residue as the inhibitor, namely Phe461. Nervonic acid is bound to LOX1 by forming two hydrogen bonds at Ser199, Tyr245 and three hydrophobic interactions at residues Pro201, Trp203, and Ala260. Nervonic acid is bound to the same three residues as the inhibitor, namely Ser199, Tyr245, and Pro201. Hexadecadienoic acid interacted with CD36 by generating three hydrogen bonds at Asn139, Ser167, and Gln171 and three hydrophobic interactions at Pro73, Met77, and Phe170. Hexadecadienoic acid is also bound to the same residue as the inhibitors of Asn139 and Ser167. Nervonic acid is bound to ACAT1 by forming two hydrogen bonds (Trp407 and Asn487) and one hydrophobic interaction (Trp408). Nervonic acid is bound to the same residue as the inhibitor at Trp407.
Molecular dynamic simulation
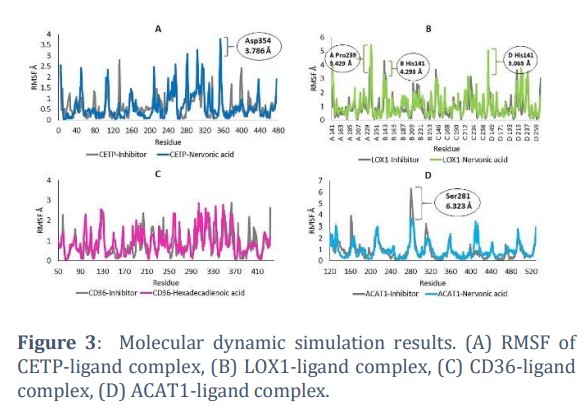
Molecular dynamics simulation results showed that there was a slight difference in conformational stability between protein-inhibitors and protein-fatty acids complex. However, most of the amino acids in each protein were still relatively stable. There is one unstable amino acid in the CETP-Nervonic acid complex, namely Asp354 with the fluctuations of 3,786 Å (Figure 3A). In the LOX1-Nervonic acid complex, there are two unstable amino acids, namely A Pro239 and D His141 with the fluctuations of 5.429 Å and 5.065 Å, respectively (Figure 3B). The conformation of the CD36-ligand protein complex showed RMSF values below 3 Å for all amino acids, which indicated that the protein conformation remained stable when interacting with Hexdecadienoic acid (Figure 3C). The conformation of ACAT1 also tends to be stable when interacting with Nervonic acid, even more stable than when ACAT interacts with inhibitors, which is characterized by high fluctuations in the Ser281 residue (6.323 Å) (Figure 3D). These unstable residues were not the residues on the protein active site, as the residues on the active site remain in a stable state. Therefore, based on RMSF it can be predicted that the interaction between the four proteins and fatty acids was stable.
Figures & Tables

The screening of fatty acids which are predicted to have anti-foam cell formation activity was carried out using drug-likeness prediction and bioactivity prediction. The screening results showed that 13 compounds had the potential to inhibit the formation of foam cells. Based on the previous research, some of these compounds have anti-atherosclerosis effects. Linolenic acid could reduce atherosclerotic plaque in blood vessels [30]. Former study revealed the ability of palmitoleic acid to delay atherosclerosis development in LDLR-KO mice [31]. Linoleic acid has anti-atherogenic activity in apoE-deficient mice [32].
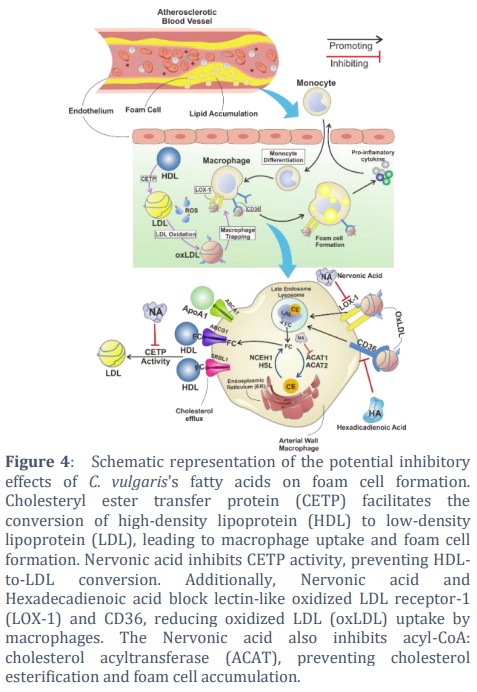
Cholesterol ester transfer protein (CETP) is a ~53 kDa protein consisting of 476 amino acids. Its banana-like structure is composed of four structural components: N-terminal domain, C-terminal domain, central sheet, and C-terminal extension [33]. The role of CETP is to mediate the transfer of CE and triglycerides between HDL, LDL, and VLDL so that HDL will turn into LDL or VLDL [34]. CETP activity results in a decrease in HDL and an increase in LDL [35]. The LDL will react with ROS to form ox-LDL, which then will enter the macrophages and causes the formation of foam cells [36]. Thus, the inhibition of CETP activity hinders the foam cell formation. Molecular docking results indicate the nervonic acid's ability to inhibit CETP activity by binding in the same area as the known inhibitor (Figure 4).
LOX1 (lectin-like oxidized low-density lipoprotein receptor 1) is a transmembrane glycoprotein that binds to ox-LDL and mediates its entry into macrophages [37]. This protein in the heterodimer structure has a heart-like shape with a tunnel along the center of the molecule [38]. After binding to LOX-1, ox-LDL will enter macrophages and stimulate the formation of foam cells [39]. Therefore, preventing the interaction between LOX-1 and ox-LDL can stop the formation of the foam cell, thus, inhibiting atherosclerosis. The inhibitor molecule used in this study was BI-0115, which was able to block ox-LDL binding [40]. The docking results in this study showed that Nervonic acid binds in the same place as the inhibitor (BI-0115). From this result, it is predicted that Nervonic acid has the ability to block ox-LDL and LOX-1binding (Figure 4).
CD36 plays an important role in mediating the entry of ox-LDL into macrophages for foam cell formation [41]. CD36 is a highly glycosylated 80kDa integral protein with a large extracellular domain containing a hydrophobic sequence domain where ligands bind [42]. The entry of ox-LDL into macrophages can be inhibited by blocking the binding of ox-LDL with CD36. The small molecule in previous studies that were able to block ox-LDL binding was nitro-oleic acid used as a control in this study [43]. The docking results showed Hexadecadienoic acid binds to CD36 in the same place as Nitro-oleic acid. Therefore, Hexadecadienoic acid is predicted to have acted as a CD36 inhibitor (Figure 4).
ACAT1 (Acyl-CoA: cholesterol O-acyltransferase 1) is an integral protein with the molecular weight of ~50 kDa, converting free cholesterol into a storage form of cholesterol ester [44,45]. This protein transfers fatty acid groups from acyl-coenzyme A to the -hydroxyl part of cholesterol to form cholesterol esters. Cholesterol esters then coalesce to form cytoplasmic lipid droplets on macrophages [46]. Inhibition of ACAT1 activity can prevent foam cell formation [47]. This study used a small molecule ACAT1 inhibitor (K604), which has been shown to inhibit ACAT1 activity by an in vitro study [48]. The docking results showed that Nervonic acid bound ACAT1 in the same site as the inhibitor (Figure 2B). Therefore, nervonic acid has a high potential as an ACAT1 inhibitor that can inhibit the formation of foam cells.
The formation of foam cells is played by several proteins such as CETP, LOX1, CD36, and ACAT1. CETP converts HDL to LDL in hepatocytes. LDL will react with oxygen to form ox-LDL. Ox-LDL is then bound to LOX1 and CD36 on macrophages and then inserts into macrophages. In macrophages, LDL is converted by ACAT1 to cholesterol esters. Cholesterol esters then precipitate and ultimately convert macrophages into foam cells. To inhibit the formation of foam cells, compounds that can inhibit the activity of these proteins are needed. In this study, it can be seen that nervonic acid and hexadecadienoic acid have the potential to inhibit proteins related to foam cell formation. Nervonic acid is predicted to inhibit CETP, LOX1, and ACAT1 activities. Meanwhile, Hexadecadienoic is predicted to inhibit CD36 protein. However, it is necessary to conduct further studies using in vitro and in vivo approaches to prove the anti-foam cell formation activity of fatty acids of C. vulgaris.
All of the fatty acids contained in Chlorella vulgaris have medicinal properties. There are thirteen compounds predicted to inhibit the formation of foam cells, namely myristoleic acid, hexadecadienoic acid, linolenic acid, palmitoleic acid, linoleic acid, heptadecenoic acid, oleic acid eicosadienoic acid, nonadecenoic acid, gadoleic acid, heneicosanoic acid, brassidic acid, and nervonic acid. Among these thirteen compounds, Nervonic acid is predicted to have the highest potential in inhibiting the activity of proteins related to foam cell formation, such as CETP, LOX1, and ACAT1. Meanwhile, hexadecadienoic acid has the best potential as a CD36 protein inhibitor.
Author Contributions
Muhammad Hermawan Widyananda contributed to the conceptualization, methodology, and writing of the original draft. Rahmat Grahadi was responsible for data curation, investigation, and validation of the results. Ichda Arini Dinana contributed to the formal analysis and visualization of the data. Arif Nur Muhammad Ansori played a key role in the writing, review, and editing of the manuscript. Viol Dhea Kharisma assisted in the literature review and data interpretation. Vikash Jakhmola provided expertise in experimental design and critical revisions. Maksim Rebezov contributed to the supervision and validation of findings. Marina Derkho was involved in reviewing and improving the manuscript. Pavel Burkov contributed to data analysis and result interpretation. Pavel Scherbakov assisted in validation. Rahadian Zainul contributed to project administration and funding acquisition.
We thank Jalan Tengah, Indonesia (https://jalantengah.site) for editing the manuscript.![]()
References
- Chan J, Karere GM, Cox LA, VandeBerg JL, Iughetti BPL, et al. Animal Models of Diet-induced Hypercholesterolemia. Hypercholesterolemia. InTech, (2015).
- Wang D, Yang Y, Lei Y, Tzvetkov NT, Liu X, et al. Targeting Foam Cell Formation in Atherosclerosis: Therapeutic Potential of Natural Products. Pharmacological Reviews, (2019); 71(4): 596-670.
- Gao S, Liu J. Association between circulating oxidized low-density lipoprotein and atherosclerotic cardiovascular disease. Chronic Diseases and Translational Medicine, (2017); 3(2): 89-94.
- Jürgens G, Hoff HF, Chisolm GM, Esterbauer H. Modification of human serum low density lipoprotein by oxidation – Characterization and pathophysiological implications. Chemistry
- Xu S, Ogura S, Chen J, Little PJ, Moss J, Liu P. LOX-1 in atherosclerosis: biological functions and pharmacological modifiers. Cellular and Molecular Life Sciences, (2013); 70(16): 2859-72.
- Yu X-H, Fu Y-C, Zhang D-W, Yin K, Tang C-K. Foam cells in atherosclerosis. Clinica Chimica Acta, (2013); 424: 245-52.
- Gupta KK, Ali S, Sanghera RS. Pharmacological Options in Atherosclerosis: A Review of the Existing Evidence. Cardiology and Therapy, (2019); 8(1): 5-20.
- Adarme-Vega TC, Lim DKY, Timmins M, Vernen F, Li Y, Schenk PM. Microalgal biofactories: a promising approach towards sustainable omega-3 fatty acid production. Microbial Cell Factories, (2012); 11(1): 96.
- Freitas HR. Chlorella vulgaris as a Source of Essential Fatty Acids and Micronutrients: A Brief Commentary. Open Plant Science Journal, (2017); 10(1): 92-9.
- Richmond A. Handbook of microalgal culture: biotechnology and applied phycology. Oxford Ames: Blackwell Science, (2004).
- Ru ITK, Sung YY, Jusoh M, Wahid MEA, Nagappan T. Chlorella vulgaris?: a perspective on its potential for combining high biomass with high value bioproducts. Applied Phycology, (2020); 1(1): 2-11.
- Montone CM, Aita SE, Catani M, Cavaliere C, Cerrato A, Piovesana S, et al. Profiling and quantitative analysis of underivatized fatty acids in Chlorella vulgaris microalgae by liquid chromatography?high resolution mass spectrometry. Journal of Separation Science, (2021); 44(16): 3041-51.
- Steinbrecher UP. Oxidation of human low density lipoprotein results in derivatization of lysine residues of apolipoprotein B by lipid peroxide decomposition products. Journal of Biological Chemistry, (1987); 262(8): 3603-8.
- Chang CL, Deckelbaum RJ. Omega-3 fatty acids: mechanisms underlying 'protective effects' in atherosclerosis. Current Opinion in Lipidology, (2013); 24(4): 345-50.
- Chovan?íková M, Šimek V. Effects of high-fat and Chlorella vulgaris feeding on changes in lipid metabolism in mice. Biologia Bratislava, (2001); 56(6): 661-666.
- Ekins S, Mestres J, Testa B. In silico pharmacology for drug discovery: methods for virtual ligand screening and profiling: In silico pharmacology for drug discovery. British
- Zloh M, Kirton SB. The benefits of in silico modeling to identify possible small-molecule drugs and their off-target interactions. Future Medicinal Chemistry, (2018); 10(4): 423-32.
- Orekhov AN, Sobenin IA, Revin VV, Bobryshev YV. Development of Antiatherosclerotic Drugs on the basis of Natural Products Using Cell Model Approach. Oxidative Medicine and Cellular Longevity, (2015); 2015: 1-11.
- Montone CM, Aita SE, Catani M, Cavaliere C, Cerrato A, Piovesana S, et al. Profiling and quantitative analysis of underivatized fatty acids in Chlorella vulgaris microalgae by liquid chromatography-high resolution mass spectrometry. Journal of Separation Science, (2021); 44(16): 3041-51.
- Daina A, Michielin O, Zoete V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Scientific Reports, (2017); 7: 1-13.
- Widyananda MH, Muflikhah L, Ulfa SM, Widodo N. Unveiling the antibreast cancer mechanism of Euphorbia hirta ethanol extract: computational and experimental study. Journal of Biologically Active Products from Nature, (2024); 14(3): 359-82.
- Dallakyan S, Olson AJ. Small Molecule Library Screening by Docking with PyRx, (2015).
- Widyananda MH, Wicaksono ST, Rahmawati K, Puspitarini S, Ulfa SM, Jatmiko YD, et al. A Potential Anticancer Mechanism of Finger Root (Boesenbergia rotunda) Extracts against a Breast Cancer Cell Line. Scientifica, (2022); 2022: 1-17.
- Widyananda MH, Pratama SK, Samoedra RS, Sari FN, Ansori ANM, Antonius Y. Molecular docking study of sea urchin (Arbacia lixula) peptides as multi-target inhibitor for non-small cell lung cancer (NSCLC) associated proteins. Journal of Pharmacy & Pharmacognosy Research, (2021); 9(4): 484-96.
- Trott O, Olson AJ. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. Journal of Computational Chemistry, (2009); 31(2): 455-461
- Kumar N, Sood D, Tomar R, Chandra R. Antimicrobial Peptide Designing and Optimization Employing Large-Scale Flexibility Analysis of Protein-Peptide Fragments. ACS Omega, (2019); 4(25): 21370-80.
- Hammoudi N-E-H, Sobhi W, Attoui A, Lemaoui T, Erto A, Benguerba Y. In silico drug discovery of Acetylcholinesterase and Butyrylcholinesterase enzymes inhibitors based on Quantitative Structure-Activity Relationship (QSAR) and drug-likeness evaluation. Journal of Molecular Structure, (2021); 1229: 129845.
- Benet LZ, Hosey CM, Ursu O, Oprea TI, Sciences T, Division I. HHS Public Access. Advanced Drug Delivery Reviews, 2017; 101: 89-98.
- Filimonov DA, Lagunin AA, Gloriozova TA, Rudik AV, Druzhilovskii DS, Pogodin PV, et al. Prediction of the Biological Activity Spectra of Organic Compounds Using the Pass Online Web Resource. Chemistry of Heterocyclic Compounds, (2014); 50(3): 444-57.
- Djoussé L, Folsom AR, Province MA, Hunt SC, Ellison RC. Dietary linolenic acid and carotid atherosclerosis: the National Heart, Lung, and Blood Institute Family Heart Study. The American Journal of Clinical Nutrition, (2003); 77(4): 819-25.
- Rencüzoğullari I, Karabağ Y, Cağdaş M, Adali Y, Karakoyun S, et al. Impact of supplementation with branched chain amino acids on myocardium and coronary in regularly and intensively exercising rats. Kafkas Universitesi Veteriner Fakultesi Dergisi. (2018); 24(3): 459-466 .
- Sato M, Shibata K, Nomura R, Kawamoto D, Nagamine R, Imaizumi K. Linoleic acid-rich fats reduce atherosclerosis development beyond its oxidative and inflammatory stress-increasing effect in apolipoprotein E-deficient mice in comparison with saturated fatty acid-rich fats. British Journal of Nutrition, (2005); 94(6): 896-901.
- Zhang M, Charles R, Tong H, Zhang L, Patel M, Wang F, et al. HDL surface lipids mediate CETP binding as revealed by electron microscopy and molecular dynamics simulation. Scientific Report, (2015); 5(1): 8741.
- Barter PJ, Rye K-A. Cholesteryl Ester Transfer Protein Inhibition Is Not Yet Dead-Pro. Arteriosclerosis, thrombosis, and vascular biology, (2016); 36(3): 439-41.
- Chapman MJ, Orsoni A, Robillard P, Therond P, Giral P. Duality of statin action on lipoprotein subpopulations in the mixed dyslipidemia of metabolic syndrome: Quantity vs quality over time and implication of CETP. Journal of Clinical Lipidology, (2018); 12(3): 784-800.e4.
- Ganesan R, Henkels KM, Wrenshall LE, Kanaho Y, Di Paolo G, Frohman MA, et al. Oxidized LDL phagocytosis during foam cell formation in atherosclerotic plaques relies on a PLD2-CD36 functional interdependence. Journal of Leukocyte Biology, (2018); 103(5): 867-83.
- Kattoor AJ, Goel A, Mehta JL. LOX-1: Regulation, Signaling and Its Role in Atherosclerosis. Antioxidants, (2019); 8(7): 218.
- Park H, Adsit FG, Boyington JC. The 14 Å Crystal Structure of the Human Oxidized Low Density Lipoprotein Receptor Lox-1. Journal of Biological Chemistry, (2005); 280(14): 13593-9.
- Stein S, Lohmann C, Schäfer N, Hofmann J, Rohrer L, Besler C, et al. SIRT1 decreases Lox-1-mediated foam cell formation in atherogenesis. European Heart Journal, (2010); 31(18): 2301-9.
- Schnapp G, Neubauer H, Büttner FH, Handschuh S, Lingard I, Heilker R, et al. A small-molecule inhibitor of lectin-like oxidized LDL receptor-1 acts by stabilizing an inactive receptor tetramer state. Communications Chemistry, (2020); 3(1): 75.
- Rahaman SO, Lennon DJ, Febbraio M, Podrez EA, Hazen SL, Silverstein RL. A CD36-dependent signaling cascade is necessary for macrophage foam cell formation. Cell Metabolism, (2006); 4(3): 211-21.
- Pepino MY, Kuda O, Samovski D, Abumrad NA. Structure-Function of CD36 and Importance of Fatty Acid Signal Transduction in Fat Metabolism. Annual Review of Nutrition, (2014); 34(1): 281-303.
- Vazquez MM, Gutierrez MV, Salvatore SR, Puiatti M, Dato VA, Chiabrando GA, et al. Nitro-oleic acid, a ligand of CD36, reduces cholesterol accumulation by modulating oxidized-LDL uptake and cholesterol efflux in RAW2647 macrophages. Redox Biology, (2020); 36: 101591.
- Cho KH, An S, Lee WS, Paik YK, Kim YK, Jeong TS. Mass-production of human ACAT-1 and ACAT-2 to screen isoform-specific inhibitor: A different substrate specificity and inhibitory regulation. Biochemical and Biophysical Research Communications, (2003); 309(4): 864-72.
- Dove DE, Su YR, Swift LL, Linton MF, Fazio S. ACAT1 deficiency increases cholesterol synthesis in mouse peritoneal macrophages. Atherosclerosis, (2006); 186(2): 267-74.
- Rogers MA, Liu J, Song BL, Li BL, Chang CCY, Chang TY. Acyl-CoA:cholesterol acyltransferases (ACATs/SOATs): Enzymes with multiple sterols as substrates and as activators. The Journal of Steroid Biochemistry and Molecular Biology, (2015); 151: 102-7.
- Yang L, Yang JB, Chen J, Yu GY, Zhou P, Lei L, et al. Enhancement of human ACAT1 gene expression to promote the macrophage-derived foam cell formation by dexamethasone. Cell Research, (2004); 14(4): 315-23.
- Ohmoto T, Nishitsuji K, Yoshitani N, Mizuguchi M, Yanagisawa Y, Saito H, et al. K604, a specific acyl-CoA:cholesterol acyltransferase 1 inhibitor, suppresses proliferation of U251-MG glioblastoma cells. Molecular Medicine Reports, (2015); 12(4): 6037-42.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


