

Full Length Research Article
Comparison of Genetic Diversity Indicators in Samples of Artificial and Natural Populations of Russian Sturgeon and Beluga at the Mouth of the Ural River
Nurbek Ginayatov1, Vadim Ulyanov1, Indira Beishova1*, Tatyana Ulyanova1*, Aziza Sidarova1, Alexandr Kovalchuk1, Bekbol Sariyev1, Ulbolsyn Kuzhebayeva1, Anna Bakhareva2, Kuantar Alikhanov3
Adv. life sci., vol. 11, no. 4, pp. 943-952, November 2024
*– Corresponding Author: Indira Beishova (indira.beishova@mail.ru)
Authors' Affiliations
2. K.G. Razumovsky Moscow State University of Technologies and Management (The First Cossack University), Moscow – Russian Federation
3. Kazakh National Agrarian Research University, Almaty – Republic of Kazakhstan
[Date Received: 01/07/2024; Date Revised: 15/08/2024; Date Published: 24/11/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Despite its ecological adaptability, sturgeon exhibits a low nuclear DNA evolution rate, enabling the use of the same primer sets for analyzing different Acipenseriformes species. The purpose of the paper was to analyze the indicators of genetic polymorphism of sturgeon populations in closed water supply systems compared with natural populations for conservation and restoration.
Methods: 147 sturgeon specimens, including Acipenser gueldenstaedtii and Huso huso, were selected from natural populations in the Ural River delta and artificial populations in aquaculture farms. DNA was extracted from fin tissues, and genetic diversity was assessed using seven short tandem repeat markers.
Result: The study showed significant genetic diversity in natural and artificial populations. Natural A. gueldenstaedtii populations demonstrated a higher level of genetic diversity (He=0.871, Ne=8.691) compared with artificial A. gueldenstaedtii populations (He=0.829, Ne=5.980). Similarly, artificial H. huso populations showed lower genetic diversity (He=0.554, Ne=2.704) than natural populations (He=0.663, Ne=3.238).
Conclusion: The analysis showed a deficiency of heterozygotes at many loci due to inbreeding, which highlights the importance of genetic management in aquaculture practice. The results highlight the need to implement genetic management strategies in sturgeon aquaculture to preserve genetic diversity and prevent inbreeding. Regular genetic monitoring and strategic breeding programs are recommended to ensure the sustainability and persistence of cultivated and natural sturgeon populations.
Keywords: Short tandem repeat; DNA polymorphism; Diversity, Huso huso; Acipenser gueldenstaedtii
Introduction![]()
Industrial aquaculture using recirculating aquaculture systems (RAS) is a developing industry globally due to the increased demand for commercial sturgeons and black caviar. Kazakhstan has a problem associated with low fish consumption per capita up to 4.3 kg/year, with a recommended rate of 14 kg/year. Therefore, the relevance of the development of the fish farming industry has increased significantly. As in many other countries, in Kazakhstan, traditional fishing is gradually giving way to artificial breeding and rearing of fish as a result of the anthropogenic impact of human activities on rivers and seas [1,2]. This method of fish farming is an effective tool for replenishing the natural sturgeon populations on the verge of extinction. The cultivation of sturgeon species in RAS allows for preserving pure populations (clean lines) without obtaining hybrids due to the control of fertilization processes, whereas in natural conditions, the number of pure populations of sturgeon decreases. In the natural environment, most sturgeon species have a low reproductive purity of the population due to changing environmental conditions (temperature, current, spawning sites, etc.) [3,4]. Despite the advantages of growing sturgeon in RAS, one of the main constraining problems is the limited choice of pairs for the reproduction of sturgeon, which determines the need to identify brood fish and breeding stock of fish to exclude closely related crosses, which will allow preserving the existing genetic structure and avoiding depletion of the gene pool [5,6].
When selecting brood fish pairs in fish farms, the risk of closely related crossing increases, inevitably leading to an increase in homozygosity in offspring and inbred depression, i.e., a decrease in vitality and fertility and an increase in harmful recessive mutations in young fish [7-10]. Yu.P. Altukhov, G.D. Ryabova, and E.I. Shishanova note that under artificial conditions, genetic diversity is depleted, and the reproduced population is increasingly less adapted to the environment and does not possess the full genetic diversity of the natural population [11-13]. Therefore, when carrying out measures aimed to maintain the number of sturgeons by releasing young fish into their natural habitat, it is necessary to apply a competent genetic approach allowing an assessment of the genetic diversity of the breeding stock and the degree of kinship of brood fish to exclude genetic degeneration of the population due to inbreeding.
In response to the transition to new conditions of existence (changes in the chemical composition of water and temperature [14], the appearance of toxic agents [15,16]), fish react with adaptations. When breeding fish in RAS, new factors (thermal waters, artificial feeds, high planting density, and industrial cultivation methods) create stressful conditions for fish [17].
In the conditions of constantly acting altered factors of existence, the role of stable genotypes ensuring selective success increases [18]. Therefore, there is a need to study the processes that form the genetic structure of rearing and breeding stock in these conditions. The formation of valuable rearing and breeding stock as an alternative to the removal of fish from natural populations for reproduction requires consideration of the impact of artificial reproduction conditions on changes in their genetic diversity.
A modern approach to studying the genetic diversity of fish populations and artificial fish stock and interspecific and species identification of specimens associated with the effective selection of brood fish pairs and the establishment of their geographical origin is the use of molecular markers [19]. Depending on the task, various approaches are used to study the genetic characteristics of sturgeons. Thus, studies have been conducted to evaluate the polymorphism of isoenzyme markers by electrophoresis of blood proteins in sturgeon [12,20] and the genetic variability of domesticated and natural sturgeon stock [21]. On the other hand, the identification of sturgeons, interspecific hybrids, and commercial products is based on the study of nucleotide sequences using such methods for detecting DNA polymorphism as RAPD (Randomly Amplified Polymorphic DNA). These markers were used in the work of K.V. Rozhkovan et al. [22] for the identification of four interspecific hybrids from the crossing of Acipenser schrenckii × Acipenser baerii × Acipenser ruthenus. Another of the main methods of nuclear DNA research is the analysis of the allelic composition of microsatellite loci. These DNA sections, which are not subjected to selective pressure and evolve at their own pace, gradually accumulate inherited differentiating features, the analysis of which allows for interspecific, intraspecific, and individual identification of sturgeons [23]. By analyzing the polymorphism of microsatellite loci, we established the species affiliation of sturgeon (Acipenseridae) and identified specimens of hybrid origin [23]. We established the geographical affiliation of specimens from fish farms [24] and assessed the genetic variability of aquacultural brood fish of Siberian sturgeon of various origins from 13 farms in the Russian Federation [25]. We studied the genetic diversity of natural populations of sterlet A. ruthenus, Acipenseridae, and two rearing and breeding stocks in the Perm Territory [26] and comparatively assessed the genetic structure of the sterlet population in the Irtysh River and the rearing and breeding stock in Tyumen [27] and indicators of genetic diversity and originality of three natural populations and three rearing and breeding stocks of sterlet (A. ruthenus, Acipenseridae) in the Volga Federal District [28]. We studied the geographical genetic structure of the lake sturgeon (Acipenser fulvescens) in its Canadian range [29] and the genetic structure of Atlantic sturgeon populations from 13 spawning rivers [30]. Based on the 18S rDNA sequencing data, we analyzed the phylogenetic relationships of the Amur sturgeon A. schrenckii [31]. We developed population genetic markers based on the variability of inter-satellite DNA [32]. Based on the literature data, the study of the genetic diversity of sturgeon is relevant to this day, which stimulates the continuation of scientific research to solve existing problems.
Despite its high ecological adaptability, sturgeon is characterized by a low rate of nuclear DNA evolution [33], which allows for using the same sets of primers for the analysis of different Acipenseriformes species [34].
Thus, the study aimed to analyze and compare the genetic polymorphism of sturgeon populations grown in closed water supply systems (CWSS) compared with natural populations.
Methods![]()
Sampling
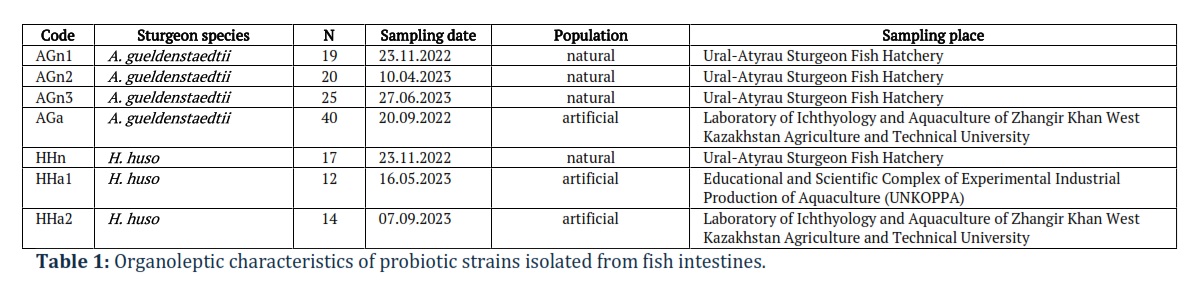
In total, 147 specimens belonging to the sturgeon family were analyzed in our study (104 Acipenser gueldenstaedtii representatives, 43 Huso huso representatives). All specimens, details of sampling and on the origins, population are indicated in Table 1.
Samples were taken from 40 A. gueldenstaedtii specimens and 26 H. huso specimens. Samples were also taken from the natural population (64 A. gueldenstaedtii specimens and 17 H. huso specimens) caught in the delta of the Ural River and contained in RAS at the Ural-Atyrau Sturgeon Fish Hatchery RSE, which is a complex of engineering structures designed to grow young sturgeon species to replenish the stocks in the basin of the Caspian Sea. The Laboratory of Ichthyology and Aquaculture of Zhangir Khan West Kazakhstan Agriculture and Technical University is a modern laboratory aimed at conducting scientific research at aquaculture facilities, including on sturgeon grown in RAS. UNKOPPA is the largest aquaculture complex in Central Asia, where about 35 tons of sturgeon are grown (breeding stock, rearing and breeding stock, commercial fish, and young fish) and which serves as a technical and research basis for the introduction of new developments in fish farming and industrial aquaculture.
DNA extraction
DNA extraction, genotyping, and processing of the results were performed in the Laboratory of Biotechnology and Diagnostics of Infectious Diseases of the Testing Center of the Zhangir Khan West Kazakhstan Agriculture and Technical University.
Fragments of the pectoral fin were selected as material for DNA extraction and short tandem repeat (STR) analysis. Each selected sample was assigned an identification laboratory number. DNA was extracted from fin tissues using a commercial DNA-Extran-2 kit (SYNTOL, Russia). The quality and quantity of the isolated DNA material was determined using a spectrophotometer.
Genetic analysis
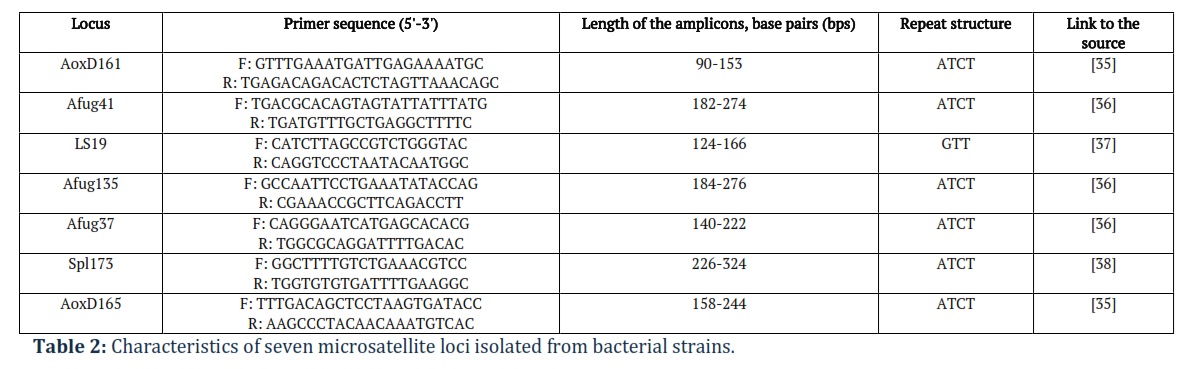
For genotyping, we used a GeneXpert-Sturgeon reagent kit (SYNTOL, Russia) designed for the genetic typing of sturgeon species. Genotyping was performed at seven STR loci. Characteristics of all seven loci are indicated in Table 2, including primer sequence. This kit allows the typing of most specimens of sturgeon species. The genotyping results according to these loci can be reproduced in other laboratories; all these loci are amplified in multiplex polymerase chain reaction (PCR).
The amplification reaction was carried out according to the manufacturer’s instructions to the genotyping kit on the ProFlex PCR system thermal cycler (Applied Computer Systems, USA).
The separation of the obtained DNA fragments was carried out by capillary electrophoresis in a polymer gel using the AB 3500 genetic analyzer (Applied Microsystems, USA). The interpretation of the analysis results was carried out using the GeneMapper 6.0 software (Applied Biosystems).
Data analysis
Genetic diversity was analyzed using a specialized add-in for Microsoft Excel, GenAlEx 6.51 [39]. We evaluated the following indicators: allele frequency, number of alleles per locus, effective number of alleles (Na), observed (Ho) and expected (He) heterozygosity, fixation index (Fis), and Nei genetic distance index [40]. Cluster analysis was performed using the Structure 2.3.4 software based on the Markov Chain Monte Carlo (MCMC) method [41]. The number of clusters (C) ranged from 1 to 10. To analyze the results of Structure and identify the most optimal C value, a portable version of the Structure Harvester V. 0.6 software was used, which uses Evanno methods to calculate the Delta K index and determine the optimal number of clusters by sequentially iterating through them [42]. The results of the Structure launches were summarized using the CLUMPP software [43]. The resulting Q-matrices were presented in the form of histograms.
Results![]()
Phenotypic traits
As a result of the study of DNA polymorphism at seven STR loci, 104 specimens from four studied A. gueldenstaedtii samples showed 16 alleles at the AoxD161 locus, 28 alleles at the Afug41 locus, 13 alleles at the LS19 and Afug135 loci, 23 at the Afug37 locus, 20 at the Spl173 locus, and 17 at the AoxD165 locus. Concerning the 43 specimens in three studied H. huso samples, 5 alleles were found at the AoxD161 locus, 12 at the Afug41 locus, seven alleles each at the LS19, Afug135, Afug37, and Spl173 loci, and eight alleles at the AoxD165 locus . Some of the identified alleles were detected with a low frequency (less than 0.1), which may indicate the risk of loss of these alleles in the future in the studied populations.
Among the belugas, allele 100 at the AoxD161 locus (more than 0.6 in each population), allele 198 at the Afug135 locus (frequency from 0.4 to 0.7 depending on the population), and allele 190 (AoxD165) with a frequency from 0.6 to 0.92 in the artificial population of UNKOPPA (HHa1) were distinguished with high frequency. As for the samples of Russian sturgeon, the maximum frequency of alleles did not exceed 0.4, and a decrease in the number of alleles and, accordingly, an increase in frequencies was more typical for the artificial population of A. gueldenstaedtii (AGa).
The I index characterizes genetic diversity; the higher this indicator, the higher the level of diversity in the sample. In our case, the values of I are high in the samples of Russian sturgeon from natural populations (more than 2) and slightly lower in the sample from an artificial population, decreasing up to 1.8 at the Afug135 locus. Samples from beluga populations are characterized by lower I index values from 0.345 to 1.691, while the lowest index values were observed in the HHa1 sample.
The Ho level in almost all loci in the Russian sturgeon samples was below the He level, which indicates a deficiency of heterozygotes, probably as a result of inbred crossing in the population. This is also indicated by the positive value of the F index. The situation differs among the three samples from the beluga populations. In the HHn and HHa1 samples, Ho is higher than He in most loci (the F index is negative), while in the HHa2 sample, only one locus (LS19) out of seven is characterized by a higher level of Ho (0.786) over He (0.673). This fact suggests that the HHn and HHa1 samples have a higher level of heterozygosity compared to the HHa2 sample.
There are not many works in scientific literature on the genetic structure of sturgeons, and even fewer used STR markers for genotyping; there is a wide variety of microsatellite loci suitable for study. Therefore, there is not much work to compare the data we obtained with those available in world databases.
In the samples of the Russian sturgeon, compared with the samples of the beluga, a significantly higher number of alleles is observed for each locus, which is associated with the tetraploidy of these sturgeon species. However, in this image, a decrease in the number of alleles in the AGa sample can be traced for almost every locus except AoxD161, where the number of alleles is comparable to samples from natural populations. The reverse situation is typical for samples from beluga populations: the number of alleles in the sample from the artificial HHa2 group differs favorably in almost all loci from the other two samples, while the situation is similar in the HHn and HHa1 samples. This fact indicates a lack of brood fish in the natural environment and RAS at UNKOPPA and a better selection of parent pairs in Laboratory of Ichthyology and Aquaculture of Zhangir Khan West Kazakhstan Agriculture and Technical University compared with the second RAS studied by us.
The He value was established for the study of genetic diversity. The He values are high, especially for the Russian sturgeon, as a polyploid species, while for the beluga, which is a diploid, it was significantly lower and ranged from 0.554 to 0.663. For the total sample of sturgeon, the He was 0.750. For the samples of Russian sturgeon, this indicator is the highest in the sample AgN3 (He=0.871), and the lowest in the sample AGa (He=0.829). Among the beluga, the highest He value was characterized by the HHa2 sample (He=0.663), and the lowest value was found in the HHa1 sample (He=0.554).
In all groups of Russian sturgeon, the possibility of inbreeding can be considered since they had a lack of heterozygotes for all the studied loci, the only exception was the Afug41 locus in the A. gueldenstaedtii AGa group. In the studied beluga groups in HHa2, six of the seven studied loci had a deficiency of heterozygotes, which indicates the possibility of inbreeding in this group.
The Russian sturgeon has the highest Na and Ne values in the AGn1 sample (Na=14,571; Ne=8,691), and the lowest in the AGa sample (Na=10,429; Ne=5,980). In beluga, the highest Na and Ne values were observed in the sample of artificial origin HHa2 (5,571 and 3,238, respectively), and the lowest in the sample of natural origin HHn (3,714 and 2,822, respectively). The low indicators of genetic diversity of the natural beluga sample can be explained by the fact that, first, the number of brood fish of this species is limited in natural conditions and, second, a small number of specimens were studied.
Results demonstrated that Na is the average number of alleles per locus, Na freq. ≥5% is the average observed number of alleles with a frequency of ≥5% per locus, Ne is the effective number of alleles, No. of private alleles is the average number of unique alleles in a population, and He is the expected heterozygosity. Thus, the average number of alleles with a frequency of ≥5% was ≈50% for samples from natural populations of A. gueldenstaedtii (AGn1, AGn2, AgN3) and more than 60% for the AGa sample. For samples from H. huso populations, the number Na freq. ≥5% was a significant 70% in the HHa1 sample, 79% in the HHa2 sample, and 88% in the natural HHn population sample. The effective number of alleles was more than 50% of the average number of alleles for each sample, while unique alleles were found in each group of samples (No. of private alleles) except a sample of H. huso from UNKOPPA (HHa1).
Analysis of genetic distances
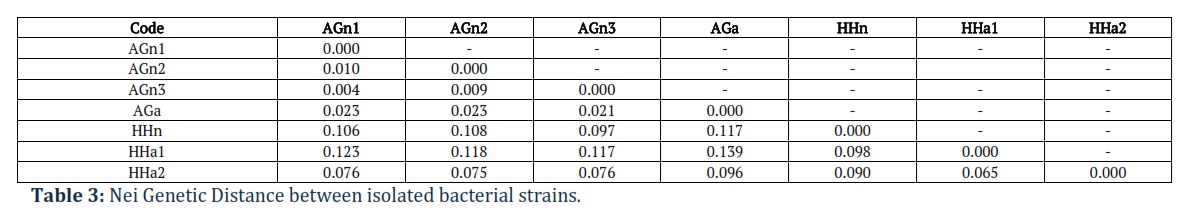
The average intra-group distances of the Russian sturgeon and beluga for each of the species were approximately the same, and the distances between natural and artificial groups were greater, as expected, as Nei Genetic Distance indicated in Table 3.
There is a clear division of natural and artificial samples in both A. gueldenstaedtii and H. huso. There is a high similarity of two samples caught from natural conditions at different times, namely, AGn1 selected on 23.11.2022 and AgN3 selected on 27.06.2023. Here, repeated sampling from the same specimen is excluded, since newly caught specimens were scanned for the presence of a chip and chipped in case of its absence; all obtained genotypes were checked for duplication.
Cluster analysis
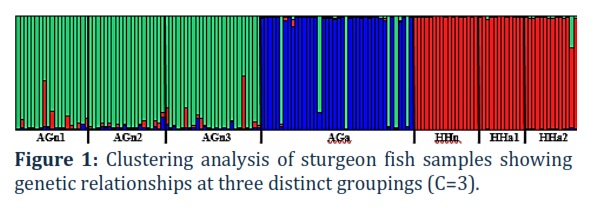
When conducting a cluster analysis of the genotyping results in the Structure program, the C value was set from 1 to 10. However, in all cases, starting from C =4, the clustering process became uninformative. The determination of Delta C values by the Evanno method showed that the most optimal number of clusters in the analysis of all samples (n= 147) of both fish species was three, with Delta C=727.544. Clustering results of the sturgeon fish at C=3 can be seen in Figure 1.
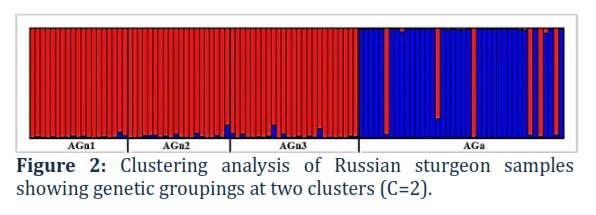
As can be seen from Figure 1, a cluster formed from 3 samples of A. gueldenstaedtii (green) of the natural population of the Ural-Atyrau Sturgeon Fish Hatchery is distinguished. A sample from the artificial population of Russian sturgeon of the Laboratory of Ichthyology and Aquaculture of Zhangir Khan West Kazakhstan Agriculture and Technical University is distinguished in a separate cluster (blue). A group of one natural and two artificial populations of beluga is formed in a separate cluster (red). As can be seen in the figure, even though all populations are kept in different conditions, there are cross-values from other species. This indicates the presence of common alleles, since genetic markers are preserved due to common origin. Nevertheless, with a separate analysis of the data for each species of studied sturgeon in the samples of belugas, there is also a division into two clusters (C=2 with Delta C=155.679), a sample of belugas from the natural population forms a separate cluster. In the case of Russian sturgeon samples, no changes are observed in a separate analysis, and two clusters are clearly distinguished separating samples from natural and artificial populations. Results of clustering of Russian sturgeon at C=2 are indicated in Figure 2.
As a result of the analysis of the structure of the studied groups of Russian sturgeon using modeling in STRUCTURE v.2.3.4 [25] based on the results of genotyping for seven STR loci, we showed two distinct clusters (C=2). In Figure 2, 2 clusters are distinguished. Formed from 3 samples of A. gueldenstaedtii (red) of the natural population of the Ural-Atyrau Sturgeon Fish Hatchery and a cluster of the artificial population of Russian sturgeon of the Laboratory of Ichthyology and Aquaculture of Zhangir Khan West Kazakhstan Agriculture and Technical University (blue).
Based on the data on the genetic polymorphism of the studied samples of Russian sturgeon and beluga, the following recommendations are given:
- To preserve the genetic resources of A. gueldenstaedtii, it is recommended to use a sample from the confluence of the Ural River with the Caspian Sea and contained in RAS at the Ural-Atyrau Sturgeon Fish Hatchery (AGn1 and AgN3) with the highest indicators of genetic diversity (He=0.867; Ne=8.691 and He=0.871; Ne=8.268);
- To compensate for the damage caused to reservoirs by fishing and human economic activity, it is recommended to release sturgeons from the rearing and breeding stock of the Ural-Atyrau Sturgeon Fish Hatchery, the Laboratory of Ichthyology and Aquaculture of Zhangir Khan West Kazakhstan Agriculture and Technical University, and UNKOPPA into the Ural River.
One of the main goals of genotyping and studying the genetic diversity of rare species of fish and sturgeons is to preserve the gene pool and prevent its depletion. The selection of parental pairs and the development of sturgeon crossing schemes becomes an important task. The data allows us to determine the degree of kinship of specimens based on microsatellite analysis data. The general sample of sturgeons is characterized by high He (He=0.750) and Ne values (Ne=5.676).
The indicators of genetic diversity in Russian sturgeon are higher in the groups AGn1 (He=0.867; Ne=8.691) and Agn3 (He=0.871; Ne=8.268), A. gueldenstaedtii from the Laboratory of Ichthyology and Aquaculture of Zhangir Khan West Kazakhstan Agriculture and Technical University caught at the confluence of the Ural River with the Caspian Sea, and lower values are observed in the sample from the Laboratory of Ichthyology and Aquaculture of Zhangir Khan West Kazakhstan Agriculture and Technical University AGa (He=0.829; Ne=5.980).
In beluga, the indicators of genetic diversity are higher in the group from the Laboratory of Ichthyology and Aquaculture of Zhangir Khan West Kazakhstan Agriculture and Technical University (He=0.663; Ne=3.238), and lower in the group from UNKOPPA (He=0.554; Ne=2.704).
In our case, with a maximum degree of kinship equal to 1, 100% of alleles in both specimens will be repeated and, conversely, if both specimens do not have the same alleles, the degree of kinship will be 0. Thus, when selecting parental pairs in RAS, it is recommended to identify (chip), as well as genotyping each specimen, and select pairs with the lowest total number of alleles. The maximum permissible value of the degree of kinship can be set at 0.25. However, it is necessary to separately consider the situation with the state of the gene pool and the number and type of genetic markers studied in each population and strive for lower values of the degree of kinship when developing crossing schemes.
Figures & Tables
The study of genetic polymorphism was carried out using microsatellite markers, the effectiveness of which for sturgeon has been shown in many works [23,44-47].
Our findings unveiled notable differences in the genetic diversity between natural and artificial populations of sturgeon species (Acipenser gueldenstaedtii and Huso huso). The data we have obtained are consistent with the opinion of the authors [52], the presence of hexaploid individuals in A. gueldenstaedtii, low I index value for natural beluga populations, lower Ho values in Russian sturgeon compared to Beluga specimens which showed higher Ho values. Our data displayed that AoxD161 locus, Afug135, and AoxD165 from the artificial population of Hh displayed a higher frequency, which is in direct contrast to the data from these authors [52]. As in our previous work, a hexasomal distribution of alleles was found at the LS19 locus in four A. gueldenstaedtii specimens caught from the natural environment [48].
The data we obtained were slightly consistent with the opinion of the authors [53], who studied the six microsatellite loci of which AoxD16, AoxD165, and AfuG41 aligned with ours and revealed that belugas showed higher He values at AoxD161 and AfuG41 loci than those observed. In our research, He values in AfuG41 loci were consistently higher in every Hh population but AoxD161 was significantly low.
The highest genetic distance was recorded in Hhn, which is consistent with the results of the authors [52]; the low genetic distance among AG specimens aligns with these authors [54], and our findings on the clustering of the conclusion that values of C or K > 3 were uninformative was confirmed by [52]. Inbreeding within artificial populations may occur due to the absence of heterozygotes in several loci, which agrees with the work reported by Roques et al. (4) on Acipenser sturio. Thus, annual monitoring and broadening the sturgeon species range are necessary for preventing the formation of such related pairs of parents. These actions are consistent with proposals made by previous studies [55].
A. Dudu et al. [49] conducted a study of 33 samples of belugas caught in the Danube River in different years (2001-2007). The analysis was carried out on seven microsatellite loci, of which only one coincides with the loci used by us (LS19). The number of alleles in this locus was nine, whereas in ours, in each of the three samples, it did not exceed five for each sample and seven for the total number of belugas studied. R. Matache [50] focuses on the genotyping of 29 beluga whales caught in the Caspian Sea, where 10 alleles were found at the LS19 locus.
N.A. Nebesikhina et al. [51] analyzed 439 brood fish of Russian sturgeon from the repair and breeding stock of the Azdonrybvod and 167 specimens caught during the spawning period in the Sea of Azov. The study was conducted on five STR markers (An20, Afug41, Afug51, AoxD161, AoxD165), the number of alleles at matching loci was not much lower than the values we obtained, except the AGa sample; the Ho and He indicators were significantly higher than ours and approached 1. According to Nebesikhina et al., the indicators of genetic diversity of samples from the natural environment were lower than the corresponding values for samples from artificial populations, although the opposite was observed in our work.
From our study, we obtained data that suggest species with high mean Ne and He and low genetic distance (AGn1, AGn3) should be implemented in RAS to increase genetic diversity, although, It is believed that RAS systems might mitigate inbreeding; the data received from our study suggest otherwise, and this may result from inadequate inbreeding or mismanagement. This finding calls for further research, a reassessment of the current RAS system, and more stringent monitoring
The obtained data on the genetic polymorphism of natural groups and those grown in RAS can be used to preserve the gene pools of Russian sturgeon and beluga typical for a region. The Ho and F values indicate a deficiency of heterozygotes in most of the studied samples as a result of closely related interbreeding within populations, which indicates a decrease in genetic diversity and a shortage of brood fish. This work should be carried out annually, covering a larger number of specimens and species of sturgeon, to monitor the state of indicators of genetic diversity, develop crossing schemes, and prevent the formation of parental pairs with a high level of genetic kinship.
Funding
The work was carried out within the framework of a grant funding project for scientific and (or) scientific and technical projects of the Ministry of Science and Higher Education of the Republic of Kazakhstan for 2022-2024 “Study of the specific features of the genetic structure of sturgeon fish and their hybrids grown in closed water supply installations” IRN AP14870980, state registration No. 0122RK00243.
Author Contributions
Nurbek Ginayatov: Conceptualization, Methodology, Data Analysis, Writing – Review & Editing. Vadim Ulyanov: Methodology, Investigation, Writing – Original Draft. Indira Beishova: Project Administration, Supervision, Writing – Review & Editing. Tatyana Ulyanova: Data Curation, Investigation, Writing – Original Draft. Aziza Sidarova: Investigation, Data Analysis, Writing – Review & Editing. Alexandr Kovalchuk: Software, Validation, Data Visualization. Bekbol Sariyev: Formal Analysis, Writing – Review & Editing. Ulbolsyn Kuzhebayeva: Investigation, Data Curation, Writing – Original Draft. Anna Bakhareva: Investigation, Resources, Writing – Review & Editing. Kuantar Alikhanov: Supervision, Funding Acquisition, Writing – Review & Editing.
The author declare that there is no conflict of interest regarding the publication of this paper.
![]() References
References
- Ambreen F, Hashmi MA, Abbas S, Kouser S, Latif F, Javed M. Genotoxic Response of Oreochromis niloticus Exposed to Tertiary Mixture of Pesticides. Advancements in Life Sciences, (2023); 9(4): 384-90.
- Ambreen HS, Arshad N, Shahzad MM, Javed GA, Shazadi K, Chaudhury FA. Animal Blood supplemented diet can improve growth performance, body composition and blood profile of Genetically Improved Farm Tilapia (Oreochromis niloticus). Advancements in Life Sciences, (2023); 9(4): 567-73.
- Kamelov AK, Sokolskii AF, Alpeisov ShA. Sovremennoe sostoyanie i podkhody k vosstanovleniyu chislennosti osetrovykh Uralo-Kaspiiskogo basseina [Current state and approaches to restoring the number of sturgeon in the Ural-Caspian basin]. 2005; 135-140. Bastau, Almaty.
- Matishov GG, Matishov DG, Ponomareva EN, Sorokina MN, Kazarnikova AV, Kovalenko MV. Osnovy osetrovodstva v usloviyakh zamknutogo vodoobespecheniya dlya fermerskikh khozyaistv [Basics of sturgeon farming in closed water supply conditions for farms]. 2008; 112. YuNTs RAN, Rostov-on-Don.
- Roques S, Berrebi P, Rochard E, Accolas M. Genetic monitoring for the successful restocking of species with low diversity: The case of the critically endangered European sturgeon, Acipenser sturio. Biological Conservation, (2018); 221: 91-102.
- Wirgin I, Roy NK, Maceda L, Mattson M. DPS and population origin of subadult Atlantic sturgeon in the Hudson River. Fisheries Research, (2018); 207: 165-170.
- Sytova M. Razrabotka nauchnykh podkhodov razvitiya osetrovogo khozyaistva na osnove proslezhivaemosti produktsii iz osetrovykh ryb [Development of scientific approaches to the development of sturgeon farming based on the traceability of sturgeon products]. Trudy VNIRO, (2016); 159: 143-150.
- Chebanov MS, Galich EV. Rukovodstvo po iskusstvennomu vosproizvodstvu osetrovykh ryb [Guidelines for artificial reproduction of sturgeon]. Tekhnicheskie doklady FAO po rybnomu khozyaistvu i akvakulture [The State of World Fisheries and Aquaculture] No. 558. 2013; 325. FAO, Ankara.
- Chebanov MS. Formirovanie geneticheskoi kollektsii osetrovykh v yuzhnom filiale FGUP FSGTsR [Formation of a genetic collection of sturgeon in the southern branch of the Federal State Unitary Enterprise Federal Selection and Genetic Center for Fish Breeding]: Genetika, selektsiya i vosproizvodstvo ryb: doklady Pervoy Vserossiyskoy konferentsii. 2002; 73-80. St. Petersburg.
- Food and Agriculture Organization of the United Nations. Razvitie akvakultury. 3. Upravlenie geneticheskimi resursami. Tekhnicheskoe rukovodstvo FAO po otvetstvennomu rybnomu khozyaistvu [Aquaculture development. 3. Genetic resource management. FAO Technical Guidance on Responsible Fisheries]. No. 5, Annex 3. 2010; 154. FAO, Rome.
- Altukhov YuP. Geneticheskie posledstviya selektivnogo rybolovstva i rybovodstva [Genetic consequences of selective fishing and fish farming]. Voprosy rybolovstva, (2000); 4(8): 562-603.
- Ryabova GD, Klimonov VO, Shishanova EI. Geneticheskaya izmenchivost v prirodnykh populyatsiyakh i dometifitsirovannykh stadakh osetrovykh ryb Rossii [Genetic variability in natural populations and domesticated sturgeon stocks in Russia]. Atlas allozimov. 2008; 96. Rosselkhozakademiya, Moscow.
- Shishanova EI. Problemy sokhraneniya i ekspluatatsii populyatsii osetrovykh ryb Kaspiiskogo basseina [Problems of conservation and exploitation of sturgeon populations in the Caspian Basin]. Izvestiya Samarskogo nauchnogo tsentra Rossiiskoi akademii nauk, (2009); 1(2): 188-192.
- Hauser L, Hemingway KL, Wedderbern J, Lawrence AJ. Mollecular/cellular processes and population genetics of a species: molecular and population response: effects of pollution on fish: molecular effects and population responses. 2003; 256-288. Blackwell Science Ltd., New York.
- Cuvin-Aralar ML, Aralar EV. Resistance to a heavy-metal mixture in Oreochromis niloticus progenies from parents chronically exposed to the same metals. Chemosphere, (1995); 30: 953-963.
- Kozlova NV. Vliyanie nekotorykh toksikantov na organizm segoletkov sterlyadi [The influence of some toxic agents on the bodies of sterlet young]. Vestnik Astrakhanskogo gosudarstvennogo tekhnicheskogo universiteta, (2007); 5(40): 252-257.
- Abrosimov SS. Stress-faktory i ikh vliyanie na fiziologo-biokhimicheskii status molodi osetrovykh [Stress factors and their influence on the physiological and biochemical status of sturgeon young]. Trudy Kubanskogo gosudarstvennogo agrarnogo universiteta, (2008); 12: 93-98.
- Bolshakov VN, Moiseenko TI. Antropogennaya evolyutsiya zhivotnykh: fakty i ikh interpretatsiya [Anthropogenic evolution of animals: facts and their interpretation]. Ekologiya, (2009); 5: 323-332.
- Kozlova NV, Bazelyuk NN, Faizulina DR, Stonogina EV. Primenenie molekulyarno-geneticheskikh issledovanii v akvakulture osetrovykh ryb [Application of molecular genetic research in sturgeon aquaculture]. Vestnik Astrakhanskogo gosudarstvennogo tekhnicheskogo universiteta, (2013); 3: 113-117.
- Kuzmin EV, Kuzmina OYu. Polimorfizm lokusa miogenov u nekotorykh predstavitelei semeistva osetrovykh (Acipenseridae) [Polymorphism of the myogen locus in some representatives of the sturgeon family (Acipenseridae)]. Genetika, (2014); 50(9): 1089-1097.
- Mamonova AS, Shishanova EI. Geneticheskaya izmenchivost odomashnennykh stad russkogo osetra (Acipenser gueldenstaedtii, Brandt) [Genetic variability of domesticated stocks of Russian sturgeon (Acipenser gueldenstaedtii, Brandt)]. Vestnik Astrakhanskogo gosudarstvennogo tekhnicheskogo universiteta, (2016); 4: 83-92.
- Rozhkovan KV, Chelomina GN, Rachek EI. Molekulyarnaya identifikatsiya i osobennosti geneticheskogo raznoobraziya mezhvidovykh gibridov amurskogo osetra (Acipenser schrenckii × A. baerii, A. baerii × A. schrenckii, A. schrenckii × A. ruthenus i A. ruthenus × A. schrenckii) po dannym izmenchivosti multilokusnykh RAPD-markerov [Molecular identification and features of genetic diversity of interspecific hybrids of Amur sturgeon (Acipenser schrenckii × A. baerii, A. baerii × A. schrenckii, A. schrenckii × A. ruthenus and A. ruthenus × A. schrenckii) according to the variability of multilocus RAPD markers]. Genetika, (2008); 44(11): 1453-1460.
- Barmintseva AE, Myuge NS. Ispolzovanie mikrosatellitnykh lokusov dlya ustanovleniya vidovoi prinadlezhnosti osetrovykh i vyyavleniya osobei gibridnogo proiskhozhdeniya [Use of microsatellite loci to establish the species identity of sturgeon and identify specimens of hybrid origin]. Genetika, (2013); 49(9): 1093-1105.
- Slukvin AM, Koneva OYu, Lesyuk MI. Geneticheskaya identifikatsiya sterlyadi (Acipenser ruthenus L.), vyrashchennoi v OAO “Rybkhoz “Polese” Pinskogo raiona Brestskoi oblasti, po mikrosatellitnym markeram [Genetic identification of sterlet (Acipenser ruthenus L.) grown in Rybkhoz Polesie OJSC, Pinsk district, Brest region, using microsatellite markers]. Molekulyarnaya i prikladnaya genetika, (2009); 9: 146-152.
- Barmintseva A, Mugue N. Genetic Variation of the Siberian Sturgeon (Acipenser baerii Brandt, 1869) in Aquaculture. Russian Journal of Genetics, (2018), 54: 210-217.
- Peleeva AR, Komarova LV, Vasileva YuS. Analiz geneticheskogo raznoobraziya estestvennykh populyatsii i remontno-matochnykh stad sterlyadi na osnovanii polimorfizma mezhmikrosatellitnykh markerov [Analysis of genetic diversity of natural populations and rearing and breeding stocks of sterlet based on polymorphism of intermicrosatellite markers]. Byulleten nauki i praktiki, (2018); 4(4): 20-29.
- Kotov IA, Trofimov OV, Pak IV, Shanskikh AI. Geneticheskii polimorfizm sterlyadi (Acipenser ruthenus), obitayushchei v estestvennykh usloviyakh i vyrashchennoi v usloviyakh zavodskogo vosproizvodstva [Genetic polymorphism of sterlet (Acipenser ruthenus) living in natural conditions and reared under conditions of breeding farm reproduction]. Vodnye bioresursy i sreda obitaniya, (2022); 5(2): 75-82.
- Komarova LV, Peleeva AR, Kostitsyna NV, Melnikova AG, Boronnikova SV. Polimorfizm DNK, geneticheskaya originalnost i identifikatsiya populyatsii i remontno-matochnykh stad sterlyadi (Acipenser ruthenus) [DNA polymorphism, genetic originality and identification of populations and rearingand breeding stocks of sterlet (Acipenser ruthenus)]. Vestnik Permskogo universiteta, (2021); 1: 53-60.
- Kjartanson SL, Haxton T, Wozney K, Lovejoy NR, Wilson CC. Conservation genetics of lake sturgeon (Acipenser fulvescens): nuclear phylogeography drives contemporary patterns of genetic structure and diversity. Diversity, (2023); 15(3): e.385.
- Wirgin I, Fox AG, Maceda L, Waldman J. Two distinct life history strategies of atlantic sturgeon in the Ogeechee river, Georgia. Diversity, (2023); 15(3): 325.
- Rozhkovan KV, Chelomina GN, Ivanov SA. Filogeneticheskie svyazi amurskogo osetra Acipenser shrenskii Brandt, 1869 po dannym sekvenirovaniya 18S rDNK [Phylogenetic relationships of the Amur sturgeon Acipenser shrenskii Brandt, 1869 according to 18S rDNA sequencing data]. Tsitologiya, (2009); 51(3): 265-270.
- Melnikova MN, Senchukova AL, Pavlov SD. Razrabotka novykh populyatsionno-geneticheskikh markerov dlya vida Parasalmo (Oncorhynchus) mykiss na osnove variabelnosti mezhsatellitnoi DNK [Development of new population genetic markers for the species Parasalmo (Oncorhynchus) mykiss based on intersatellite DNA variability]. Doklady Akademii nauk, (2010); 435(1): 138-141.
- Krieger J, Fuerst PA. Evidence for a slowed rate of molecular evolution in the order Acipenseriformes. Molecular Biology and Evolution, (2002); 19(6): 891-897.
- Congiu L, Pujolar JM, Foriani A, Cenadelli S, Dupanloup I, Barbisan F, Galli A, Fontana F. Managing polyploidy in ex situ conservation genetics: the case of the critically endangered Adriatic sturgeon (Acipenser naccarii). PLoS One, (2011); 6(3): e18249-1-e18249-10.
- Henderson-Arzapalo A, King TL. Novel microsatellite markers for Atlantic sturgeon (Acipenser oxyrinchus) population delineation and broodstock management. Molecular Ecology Notes, (2002); 2: 437-439.
- Welsh AB, Blumberg M, May B. Identification of microsatellite loci in lake sturgeon, Acipenser fulvescens, and their variability in green sturgeon, A. medirostris. Molecular Ecology Notes, (2003); 3(1): 47-55.
- McQuown EC, Sloss BL, Sheehan RJ, Rodzen J, Tranah G, May B. Microsatellite analysis of genetic variation in sturgeon: new primers sequences for Scaphirhynchus and Acipenser. Transactions of the American Fisheries Society, (2000); 129: 1380-1388.
- May B, Krueger CC, Kincaid HL. Genetic variation at microsatellite loci in sturgeon: primer sequence homology in Acipenser and Scaphirhynchus. Canadian Journal of Fisheries and Aquatic Sciences, (1997); 54(7): 1542-1547.
- Peakall R, Smouse PE. GENALEX 6: genetic analysis in Excel, population genetic software for teaching and research. Molecular Ecology Notes, (2006); 6(1): 288-295.
- Nei M. Genetic distance between populations. The American Naturalist, (1972); 106(949): 283-292.
- Pritchard JK, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data. Genetics, (2000); 155(2): 945-959.
- Earl DA, vonHoldt BM. Structure harvester: a website and program for visualizing structure output and implementing the Evanno method. Conservation Genetics Resources, (2012); 4: 359-361.
- Jakobsson M, Rosenberg NA. CLUMPP: a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics, (2007); 23(14): 1801-1806.
- Georgescu S, Canareica O, Dudu A, Costache M. Analysis of the microsatellite variation in the common hybrid between Russian Sturgeon (Acipenser Gueldenstaedtii Brandt and Ratzeburg, 1833) and Siberian Sturgeon (Acipenser Baerii Brandt, 1869) from aquaculture. Transylvanian Review of Systematical and Ecological Research, (2014); 15(2): 117-124.
- Chassaing O, Hänni C, Berrebi P. Distinguishing species of European sturgeons Acipenser spp. using microsatellite allele sequences. Journal of Fish Biology, (2011); 78(1): 208-226.
- Timoshkina NN, Barmintseva AE, Usatov AV, Miuge NS. Intraspecific genetic polymorphism of Russian sturgeon Acipencer gueldenstaedtii. Russian Journal of Genetics, (2009); 45(9): 1250-1259.
- Selyukov A, Zhigileva O, Shuman L, Selyukova S, Bogdanova V. Cytomorphological and genetic indicators in the early ontogenesis of the wild and farmed broad white fish (Coregonus nasus). Aquaculture and Fisheries, (2022); 7(2): 211-222.
- Study of the genetic diversity of sturgeon fish from the natural environment and grown in aquaculture in the Western region of Kazakhstan.
- Dudu A, Georgescu SE, Burcea A, Florescu I, Costache M. Analysis of genetic diversity in Beluga Sturgeon, Huso huso (Linnaeus, 1758) from the Lower Danube river using DNA markers. Scientific Papers: Animal Science and Biotechnologies, (2014); 47(1): 64-68.
- Matache R, Holban E, Gyorgy D, Prangate R, Matei M, Boboc M, Popescu R, Nuraiti T. Determination of the hybridization rate of Acipenser stellatus and Huso huso based on microsatellite analysis. IOP Conference Series: Earth and Environmental Science, (2023); 1216: 012011.
- Nebesikhina NA, Lepeshkov AG, Ivanova EA, Timoshkina NN. Evaluation of genetic diversity in Russian sturgeon breeders kept at the Don sturgeon farm “Azdonrybvod”. AzNIIRKH Proceedings, (2017); 1: 244-249.
- Ulyanov V, Beishova I, Ulyanova T, Sidarova A, Ginayatov N, Kovalchuk A, Chuzhebaeva G, Alikhanov K, Sariyev B, Kuzhebayeva U, Nurzhanova F, Beishov R, Sabyrzhanov A, Bexultan A. Genetic health and diversity assessment of Sturgeon species in Kazakhstan’s aquaculture and natural habitats. German Journal of Veterinary Research, (2024); 4(2): 127-138.
- Slukvin AM, Dromashko SE, Balashenko NA, Barulin NV, Barmintseva AE. Study results on the molecular genetic, morphometric and sexual characteristics of the Beluga (Huso huso L., 1758) grown in the aquaculture of the Republic of Belarus. Azerbaijan Journal of Physiology, (2022); (1).
- Nebesikhina NA, Lepeshkov AG, Ivanova EA, Timoshkina NN. Assessment of the genetic diversity of Russian sturgeon producers from the broodstock of the federal state budgetary institution "Azdonrybvod" Don Sturgeon Plant, (2017); 244-249.
- Roques S, Chancerel E, Boury C, Maud P, Acolas M. From microsatellites to single nucleotide polymorphisms for the genetic monitoring of a critically endangered sturgeon. Ecology and Evolution, (2019); 9(12): 7017–7029.
- Karmaliyev R, Nurzhanova F, Sidikhov B, Murzabaev K, Sariyev N, Satybayev B, Abirova I. Epizootiology of Opisthorchiasis in carnivores, fish, and mollusks in the West Kazakhstan Region. American Journal of Animal and Veterinary Sciences, (2023); 18(2): 147-155.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()