

Review Article
Antifungal Therapy and Probiotics in Women's Health – A Comprehensive Review
Amankeldi Sadanov1, Baiken Baimakhanova1*, Asya Balgimbayeva1, Lyudmila Trenozhnikova1, Vladimir Berezin1, Olga Lakh1, Saltanat Orazymbet1, Andrey Bogoyavlenskiy1, Erik Shorabaev1, Gul Baimakhanova1, Zere Turlybaeva1, Alma Amangeldi1, Anel Omirbekova2, Aigerim Mamirova2, Aida Kistaubayeva2
Adv. life sci., vol. 11, no. 4, pp. 927-942, November 2024
*– Corresponding Authors: Baiken Baimakhanova (bbbayken@mail.ru)
Authors' Affiliations
2. Al- Farabi Kazakh National University – Almaty – Kazakhstan
[Date Received: 15/07/2024; Date Revised: 09/09/2024; Date Available Online: 24/11/2024]
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
Candida-induced infections affect millions of women around the world. Therefore, the purpose of this review is to provide a comprehensive analysis of the role of antifungal therapy and probiotics in managing one such infection, vulvovaginal candidiasis (VVC). The study employed a systematic review method to retrieve relevant articles from databases such as PubMed, MDPI, Scopus, and ResearchGate, using specific search criteria. The findings highlight the effectiveness and limitations of traditional antifungal treatment, the emergence of novel treatments targeting new biological pathways, and the potential of probiotics as adjunctive therapy. It is concluded that while traditional antifungals remain fundamental, resistance issues necessitate ongoing development of novel agents. Additionally, alternative therapies hold promise in enhancing treatment outcomes and maintaining vaginal health. The multifaceted approach in VVC treatment highlights the importance of integrating various infection control and treatment strategies into a more effective and holistic approach.
Keywords: Candida sp.; Vulvovaginal candidiasis; Antifungal agents; Probiotics
Introduction![]()
Candida-induced infections are among the most common bloodstream infections worldwide. Being a part of human microflora colonising mucosal surfaces, Candida sp. can easily switch from ‘normal’ colonisation onto the ‘pathogen’ phase [1]. Candida albicans, a dimorphic fungus, is the most known species among >160 representatives of the genus Candida, responsible for 40-70% of invasive candidiasis cases and over 90% of vulvovaginal candidiasis (VVC) [2]. Only ~20 Candida species classified as non-albicans Candida (NAC), such as C. glabrata, C. krusei, C. parapsilosis, C. tropicalis, C. dubliniensis, C. lusitaniae, C. rugosa, C. guilliermondii, C. inconspicua, and C. norvegensis, were found to be capable of inducing infections [1]. Meanwhile, the following NAC can cause VVC with degrading frequency of recorded incidence: C. glabrata, C. krusei, C. parapsilosis, and C. tropicalis. Moreover, according to the recent first fungal priority pathogens list ranking, C. auris and C. albicans belong to the critical priority group, C. glabrata, C. tropicalis, and C. parapsilosis – to high priority group. C. glabrata and C. krusei have recently been referred to as Nakaseomyces glabrata and Pichia kudriavzevii, consequently being less pathogenic for healthy, immunocompetent women. Despite the recently proposed grouping name “non-albicans clinical yeast (NACY)” [3] instead of NAC, in the current review traditional names are used to avoid confusion.
Considering the up-to-date digits reporting that 75% of women have at least one VVC episode, 5-44% of them have recurrent vulvovaginal candidiasis (RVVC), annually 135-138 M women affected by RVVC [2], claims on the inaccuracy of published values, and even worse prognosis on expected increase RVVC-affected women population up to 158 M by 2030 [3], further research is urgently needed to fulfil current gaps. No actual data was found for registered R/VVC (RVVC and VVC) incidences for Kazakhstan, except for values of 2985 cases per 100,000 females or a total of 273,258 incidences in 2015 [4]. Thus, not only investigations on developing new therapies or alternative strategies are required, but correction of imprecise estimates of fungal infection incidences [4] is also needed to adequately assess the situation. The current review focused on a thorough investigation of traditional, novel, and alternative therapies against VVC-inducer pathogens.
The goal of this review is to provide a thorough and comprehensive analysis of the role of antifungal therapy and probiotics in the management of vulvovaginal candidiasis (VVC), a common and often recurrent fungal infection affecting millions of women worldwide. By examining both traditional antifungal treatments and the emerging role of probiotics, this review seeks to explore the efficacy, mechanisms of action, safety, and long-term benefits of these therapeutic options.
Methods
![]()
Literature search strategy and selection criteria
The article presents a systematic review of distinct treatment strategies for VVC. Relevant literature was compiled via database search, which later allowed for the synthesis of the findings and the division of examined therapies into three groups of traditional, novel, and alternative approaches.
Databases used in literature collection included PubMed, MDPI, Scopus, ResearchGate. Criteria for the search were set as follows: articles should be up to date, published no earlier than in 2015; search queries included the combinations of terms ‘vulvovaginal candidiasis’, ‘Candida’, ‘vaginal microbiome’, ‘treatment’, ‘therapy’, ‘antifungal’, ‘inhibitor’. Eligible research works were vetted for the overall accuracy and relevance to the topic at hand.
Summarized findings are presented in a list of VVC treatment strategies with the description of their work principles and characteristics. Based on the literature review, classifications for the traditional and novel antifungal therapies were created in the forms of corresponding diagrams. Complimentary to the identified treatment groups, prospects of probiotics’ use as an adjunctive therapy were also discussed, as their effectiveness has been evaluated in selected scientific works.
Discussion![]()
Traditional Antifungal Therapy Against R/VVC
The history of antifungal agents dates back to 1939 when the antifungal properties of griseofulvin were discovered. Griseofulvin, a natural compound, is a metabolic product derived from Penicillium griseo-fulvum Dierckx, a species of mold [5]. The second antifungal agent, benzimidazole, was the first azole introduced in 1944. However, it was the introduction of fluconazole in 1990 that marked a breakthrough [1]. As the first broad-spectrum antifungal, fluconazole addressed key deficiencies such as poor solubility and the absence of an intravenous (IV) formulation, significantly impacting the antifungal market. The development of antifungal agents encounters a significant challenge, primarily due to the similarities between fungal and mammalian cells. Therefore, at the turn of the millennium, all discovered antifungal drugs targeted plasma membrane sterols, nucleic acid synthesis, or cell wall constituents. However, novel targets for antifungal agents are continuously developing to overcome limitations caused by the utilization of traditional antifungals, which will be discussed in the section “Novel Antifungal Treatments” in more detail.
Although historically C. albicans was believed to be a major inducer of R/VVC, being responsible for approximately 90% of detected incidences, evidence exists that NAC species can be prevailing [1]. The most common R/VVC-associated NAC are C. glabrata, C. tropicalis, C. parapsilosis and C. krusei. Moreover, NAC species are more azole-resistant compared to C. albicans [1]. A recent study on 543 strains isolated from VVC-infected patients reported that 15.7% of incidence belonged to RVVC with 84.7% of incidence caused by C. albicans, 8.66% – by C. glabrata, 1.47% – by C. krusei, 0.92% – by C. tropicalis, and 0.55% – by C. parapsilosis [5]. VVC cases caused by treatment-resistant species like Candida tropicalis, C. glabrata, and others are estimated at 25-45%. These species show resistance to common antifungal treatments, particularly azole-based drugs. This rising resistance, especially in non-albicans Candida (NAC) species, complicates treatment, leading to higher failure rates, recurrent infections, and fewer therapeutic options. The growing challenge highlights the need for new antifungal agents and strategies.
Ergosterol Synthesis Inhibitors
Considering the antifungal classes targeted to plasma membrane sterols of a pathogen, in our case R/VVC-inducers, their mechanism of action is directly linked with ergosterol synthesis. In fact, there are two ways of action connected with ergosterol: direct (polyenes) and indirect (azoles, allylamines, and thiocarbamates).
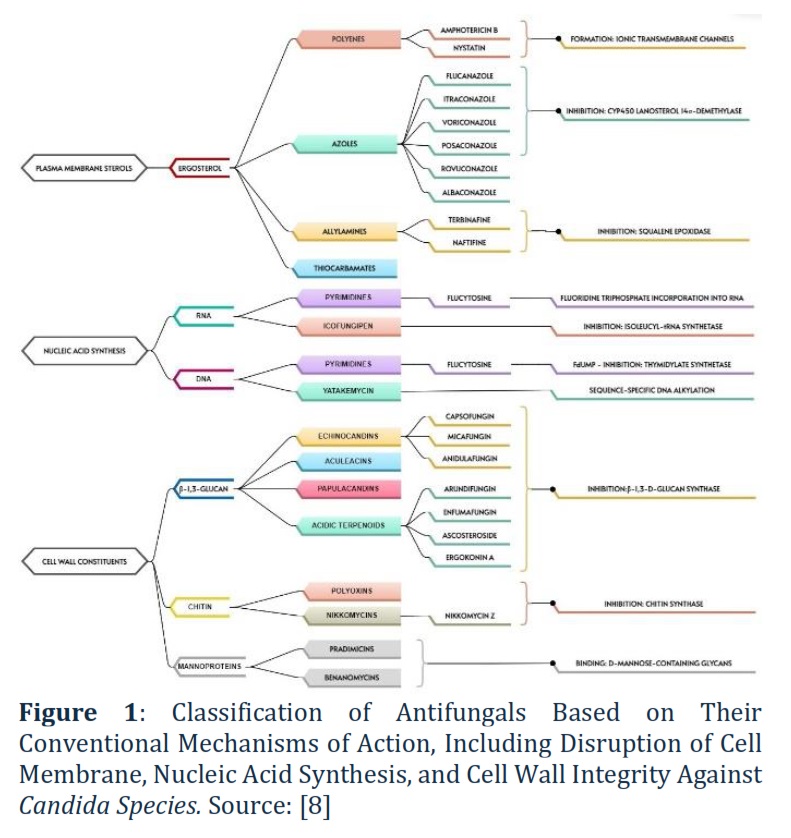
Polyenes lead to the formation of ionic transmembrane channels which in turn increases fungal membrane permeability (Figure 1). The main constraint of polyenes results from the poor ability of the drug to differentiate ergosterol from fungal cells and cholesterol in mammalian cells, which was fixed by the introduction of a lipid-based drug delivery system. Indeed, nystatin is considered efficient in the R/VVC therapy caused by fluconazole-resistant C. albicans or C. glabrata [6]. In C. albicans and C. glabrata, reduced susceptibility to amphotericin B can result from mutations in various ergosterol biosynthesis enzymes. For C. albicans, these mutations include those in ERG2, ERG3, ERG5, and ERG11, while for C. glabrata, they occur in ERG2, ERG6, and ERG11. These mutations disrupt ergosterol production, reducing the efficacy of amphotericin B, which targets fungal cell membranes. Furthermore, a reduction in ergosterol can cause polyene resistance [6]. C. albicans, C. glabrata, C. guilliermondii, C. krusei, C. lusitaniae, and C. auris were found to be resistant to amphotericin B [7].
Azoles inhibiting cytochrome P450 lanosterol 14α-demethylase results in fungal membrane integrity disruption (Figure 1). Logan et al. [6] carefully reviewed the potential mechanisms of azole resistance and defined them as follows: change in target binding site, overexpression of binding site, poor membrane penetration, and efflux pumps. A direct link between C. albicans resistance and exposure to fluconazole was defined.
Figure 1 illustrates classification of antifungals with conventional mechanisms of action against Candida sp. according to Perlin [8]. Figure 1 illustrates the variety of mechanisms by which antifungal agents act to disrupt different parts of fungal cells. They either target the cell membrane by interfering with ergosterol, inhibit nucleic acid synthesis (RNA or DNA), or damage the fungal cell wall by preventing glucan or chitin formation. For example, about Polyenes (e.g., Amphotericin B, Nystatin). These antifungal agents form ionic transmembrane channels by binding to ergosterol, a key component of the fungal cell membrane. This increases membrane permeability, leading to cell death.
Therefore, evaluation of post-antifungal effects (PAFE) should be considered obligatory to adjust the dosage and therapeutic regimens of existing and novel drugs. Albaconazole is under development for the treatment of acute VVC [8].
Allylamines (naftifine, butenafine, and terbinafine) and thiocarbamates inhibit squalene epoxidation resulting in fungal membrane damage. Terbinafine has a weak inhibitory impact against Candida sp., except for C. parapsilosis. Moreover, C. albicans resistance to terbinafine is believed to be linked to MAT locus homozygosity [8].
Nucleic Acid Synthesis Inhibitors
Pyrimidines, such as 5-fluorocytosine, act by either converting to fluorouridine triphosphate (FUTP), which gets incorporated into RNA and causes early chain termination, or to fluorodeoxyuridine monophosphate (FdUMP), which inhibits thymidylate synthetase, disrupting DNA synthesis. However, C. albicans and C. glabrata have been found to be resistant to 5-fluorocytosine [7].
Icofungipen, isolated from Bacillus cereus and Streptomyces setonii, inhibits isoleucyl-tRNA synthetase, blocking protein synthesis and fungal growth. It shows potent antifungal activity, with a minimum inhibitory concentration (MIC) of 0.5 µg/mL against Candida albicans. This unique action makes icofungipen a promising treatment, especially for resistant fungal infections. Its potential broad-spectrum efficacy could also be valuable against emerging fungal pathogens [8].
Yatakemycin derived from Streptomyces sp. TP-A0356 was reported to be more efficient compared to AmpB and itraconazole in relation to C. glabrata (azole-resistant) with minimum inhibitory concentration (MIC) of 0.0625 µg mL−1 compared to 0.125 µg mL−1 for amphotericin B and itraconazole [8].
β-1,3-Glucan Synthesis Inhibitors
Echinocandins, such as anidulafungin, caspofungin, and micafungin, are cyclic peptide antifungals that disrupt fungal cell wall synthesis by inhibiting β-1,3-glucan, effectively lysing Candida albicans. These drugs are interchangeable in terms of spectrum, safety, and clinical utility. Resistance among C. albicans and other Candida species is relatively low (1-3%), but C. glabrata shows higher resistance (10-15%), often involving multidrug resistance. This resistance is linked to mutations in the FKS1 and FKS2 genes, which reduce the effectiveness of echinocandins, particularly in C. glabrata. A recent study on site-selective reductive dehydration of caspofungin, anidulafungin, and (novel) rezafungin, specifically by removing benzylic alcohol, resulted in up to 128-fold increase in antifungal efficiency against echinocandin-resistant Candida sp., thereby developing a strategy to overcome Fks mutation-linked resistance [10].
Change in target binding sire and production of alternative cell membrane components can also cause resistance to echincocandins [6]. C. albicans, C. glabrata, and C. auris were found to be resistant to caspofungin, anidulafungin, and micafungin [7].
Chitin Synthase Inhibitors
Polyoxins and nikkomycins are peptide-nucleoside compounds produced by Streptomyces cacaoi, Streptomyces aureochromogenes, Streptomyces piomogenus and Streptomyces tendae & Streptomyces ansochromogenes, respectively. The main constraint of these classes of antifungals is low permeability leading to the different susceptibility of fungal species. Indeed, polyoxins and nikkomycins have been extensively investigated, but no product has been commercially developed.
Nowadays only five antifungal classes, specifically polyenes, azoles, pyrimidines, allylamines, and echinocandins, represent the existing antifungal armamentarium leading to the resistance development. A common challenge in treating Candida-induced infections is the development of clinical antifungal resistance. Candida species are known to employ various mechanisms to initially tolerate and eventually develop resistance to antifungal treatments, complicating effective management of these infections [11]. A recent study on the evaluation of the antifungal susceptibility of C. albicans and 9 NAC, including C. krusei, C. dubliniensis, C. glabrata, C. tropicalis, C. kefyr, C. guilliermondii, C. parapsilosis, C. lusitaniae, and C. inconspicua, concerning 5 conventional antifungals (fluconazole, voriconazole, caspofungin, micafungin, and fluorocytosine) reported the inefficiency of fluconazole and fluorocytosine for C. albicans, C. krusei, and C. dubliniesis with share of susceptible isolates of 98.0, 0, and 87.5% and 98.0, 66.7, and 87.5%, respectively [5]. Another study on evaluating the susceptibility of C. albicans (49), C. glabrata (48), C. parapsilosis (47), C. tropicalis (24), C. krusei (8), and C. guilliermondi (8) isolates to fluconazole, amphotericin B, and caspofungin reported that: a) caspofungin-resistant isolates were detected within all Candida species under research (3 isolates – C. albicans; 8 – C. glabrata; 4 – C. parapsilosis; 4 – C. tropicalis; 1 – C. krusei; 6 – C. guilliermondi); b) fluconazole-resistant isolates was prevailing among studied Candida sp. except for C. guilliermondi (6 isolates – C. albicans; 1 – C. glabrata; 2 – C. parapsilosis; 6 – C. tropicalis; all (8) – C. krusei); c) amphotericin B-resistant isolates were found only in C. glabrata (1) and C. parapsilosis (1) [11]. Thus, increasing resistance to current antifungals, their limited efficacy against biofilms, and their association with higher cytotoxicity underscore the urgent need to discover new compounds.
Novel Antifungal Treatments Against R/VVC
Cell Membrane
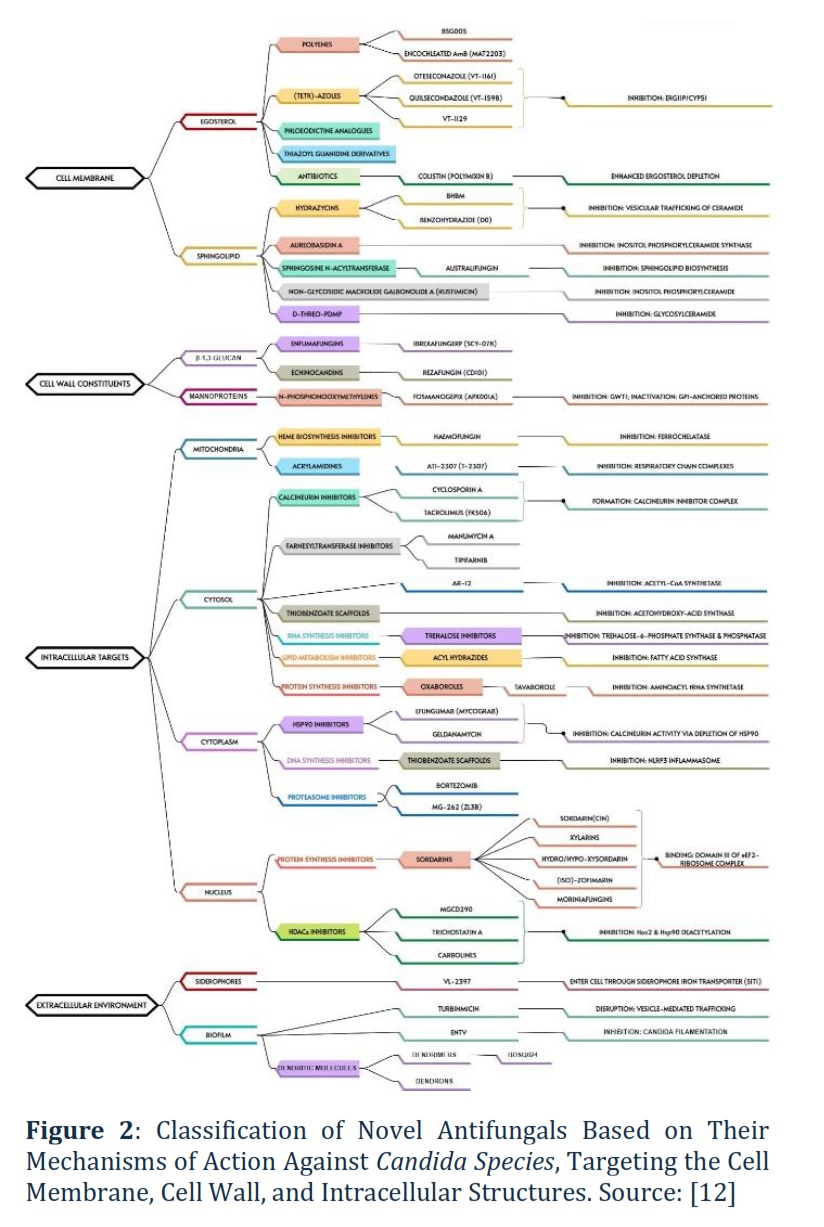
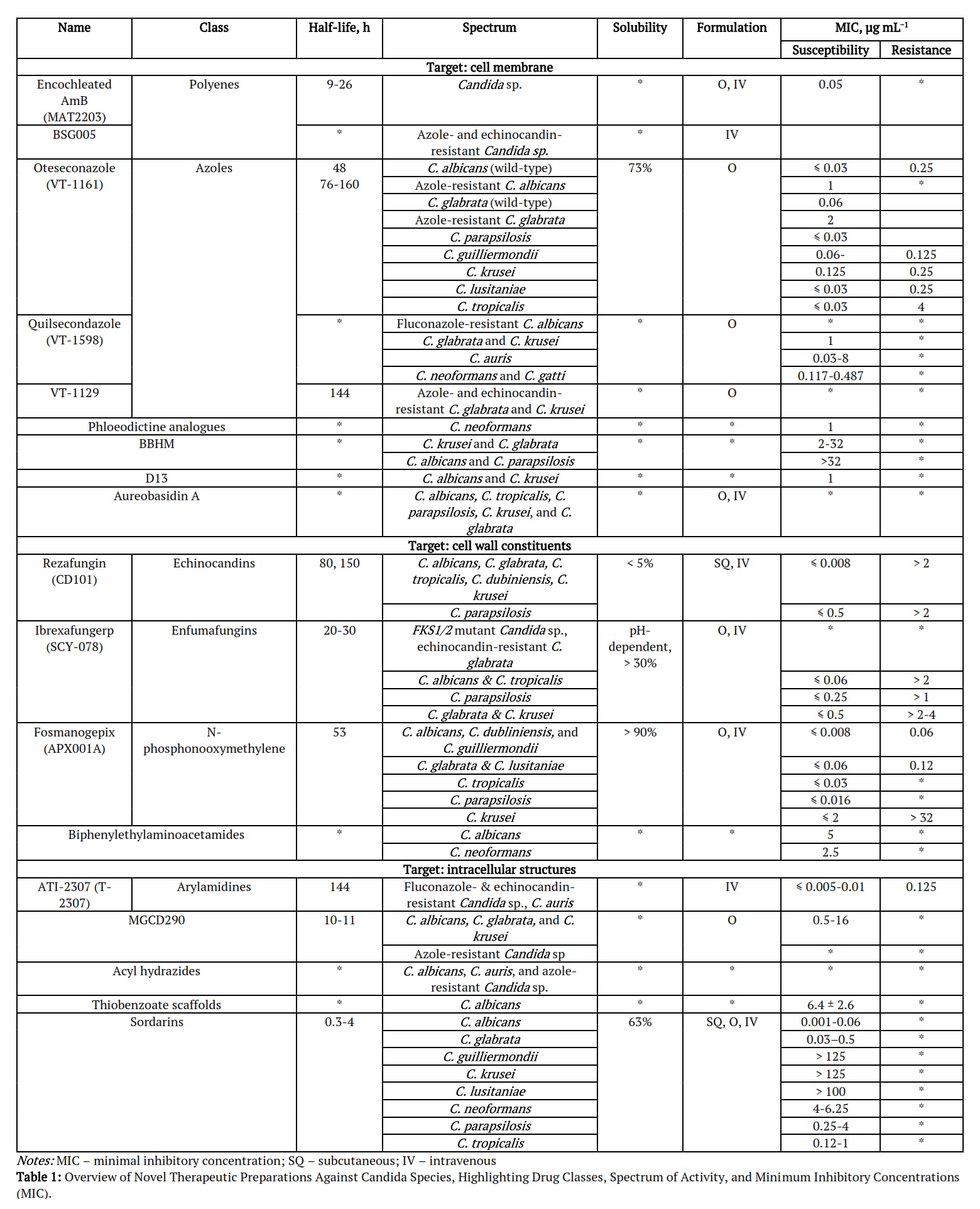
Ergosterol Synthesis Inhibitors. Encochleated AmB (MAT2203; polyenes; phase II). Activity spectrum: C. albicans, C. tropicalis, C. parapsilosis, C. krusei, C. glabrata, and C. auris [7, 12]. BSG005, a polyene antifungal compound isolated from Streptomyces noursei, has demonstrated potent fungicidal activity against a broad spectrum of fungal strains, including those resistant to commonly used antifungal classes such as azoles and echinocandins. Polyenes like BSG005 work by binding to ergosterol, a key component of the fungal cell membrane, creating pores that lead to cell leakage and eventual death. This mode of action makes BSG005 effective against resistant Candida species, offering a valuable option for infections where traditional treatments fail due to resistance [8]. Oteseconazole (VT-1161; tetrazoles; phase III) is a new oral azole drug that does not bind to and inhibit human CYP51, having greater selectivity for fungal enzymes [13]. Oteseconazole was approved by the FDA in 2022 for RVVC treatment and with particular interest to women with resistant strain-induced RVVC, allergy, or intolerance to fluconazole or even that experience drug-drug interactions. Activity spectrum: C. albicans, C. tropicalis, C. parapsilosis, C. krusei, and C. glabrata; azole- and echinocandin-resistant C. albicans [12]. Quilsecondazole (VT-1598; tetrazoles; pre-clinical) was found to be active against fluconazole-resistant C. albicans and having anticandidal activity, especially against C. auris [7]. VT-1129, a tetrazole currently in preclinical development, has shown promising in vitro activity against a variety of Candida species, including azole- and echinocandin-resistant strains of Candida glabrata and Candida krusei. Tetrazoles are a newer class of antifungal agents designed to inhibit the enzyme lanosterol 14α-demethylase, critical for ergosterol synthesis, similar to azoles but with enhanced efficacy against resistant strains. The broad-spectrum activity of VT-1129, especially against drug-resistant C. glabrata and C. krusei, highlights its potential as a novel treatment for difficult-to-treat fungal infections, addressing the growing problem of antifungal resistance in clinical settings. If successful in clinical trials, VT-1129 could provide a crucial new tool in the fight against resistant fungal pathogens.
Colistin (polymyxin B; antibiotics; pre-clinical) enhances ergosterol depletion, having minimal antifungal activity against Candida sp. (Figure 2). However, colistin has fungicidal activity when combined with fluconazole, especially in azole-tolerant strains [14]. Colistin also synergises with echinocandins in C. albicans and with caspofungin in C. auris.
Sphingolipid Synthesis Inhibitors. (E)-N′-(3-bromo-6-hydroxybenzylidene)-2methylbenzo-hydrazide (BHBM; hydrazycins; pre-clinical), benzohydrazide (D0; hydrazycins), and BBHM derivative (D13; hydrazycins) inhibit the vesicular trafficking of ceramide, a precursor lipid of glucosylceramide (GlcCer), thereby halting GlcCer and sphingolipid biosynthesis and disrupting cell division. Activity spectrum (Table 1): C. krusei and C. glabrata (BBHM MIC of 2-32 µg mL−1), C. albicans and C. parapsilosis (BBHM MIC >32 µg mL−1), C. albicans and C. krusei (D13 MIC of 1 µg mL−1). Aureobasidin A (pre-clinical), isolated from Aureobasidium pullulans, inhibits inositol phosphoryl ceramide (IPC) synthase, i.e. sphingolipid synthesis [8]. Activity spectrum: C. albicans, C. tropicalis, C. parapsilosis, C. krusei, and C. glabrata [12].
Figure 2 demonstrates the diverse antifungal targets, from the cell membrane and wall to intracellular components like mitochondria and the cytosol. Each class of antifungal works by disrupting a critical cellular process, either by inhibiting the synthesis of key structural components or by interfering with vital intracellular pathways, ultimately leading to fungal cell death. For example, N-Phosphonomethylmethylenes inactivate GPI-anchored proteins, which are vital for mannoprotein function, destabilizing the cell wall.
Cell Wall Constituents
β-1,3-Glucan Inhibitors. Ibrexafungerp (SCY-078 or MK-3118; enfumafungins; phase III), produced by Hormonema carpetanum [15], is the first oral formulation in the glucan synthase inhibitors class. The mechanism of action is similar to echinocandins, however, ibrexafungerp is structurally different [13]. Oral ibrexafungerp showed 88% efficiency in the case of VVC with a 4% recurrence rate compared to oral fluconazole of 65 and 15%, respectively.
Moreover, ibrexafungerp resistance in C. glabrata was found to be linked to the FKS2 gene, primarily involved in β-(1,3) D-glucan production in C. glabrata. Ibrexafungerp demonstrated antifungal activity against species that often exhibit reduced fluconazole susceptibility or azole resistance. Activity spectrum: C. albicans, C. tropicalis, C. parapsilosis, C. krusei, C.
glabrata, C. dubliniensis, C. lusitaniae, and C. auris [12, 16]. Moreover, ibrexafungerp has uniform and potent activity against C. auris, an emerging multidrug-resistant fungus.
Rezafungin (CD101; echinocandins; phase III; approved by FDA in 2023) showed a 95% inhibition of C. glabrata and 100% inhibition of C. albicans, C. dubliniensis, C. tropicalis, and C. krusei at MIC of 0.12 µg mL−1 [9]. Comparing the effectiveness against C. krusei, caspofungin showed fourfold higher efficiency compared to rezafungin [9]. Rezafungin, a next-generation echinocandin, is a structural analogue of anidulafungin with a modified choline group, enhancing its chemical stability, metabolic durability, and solubility. This allows for less frequent dosing and administration without an indwelling catheter, reducing associated risks. It’s particularly useful when fluconazole is contraindicated or ineffective, especially in resistant Candida infections. Its broad-spectrum activity and improved pharmacokinetics make it a promising option for treating invasive fungal infections, especially in complex or resistant cases [17]. Activity spectrum: C. albicans, C. tropicalis, C. parapsilosis (less susceptible: MIC = 4 µg mL−1), C. krusei, C. glabrata, C. dubliniensis, C. lusitaniae, C. fabianii, C. inconspicua, C. kefyr, C. lipolytica, C. pulcherrima, C. sojae, C. metapsilosis, C. orthopsilosis, C. guilliermondii, and C. auris [12, 16].
GPI-Anchor Synthesis Pathway Inhibitors. Fosmanogepix (APX001A & E1210), a compound in the N-phosphonooxymethylene class currently undergoing phase II clinical trials, exhibits potent antifungal properties through a highly targeted mechanism of action. This drug works by inhibiting the fungal enzyme Gwt1, which is crucial for the proper functioning of glycosylphosphatidylinositol (GPI) anchor proteins, also referred to as mannoproteins. These proteins are essential for fungal cell wall integrity and function. By disrupting this posttranslational modification process, Fosmanogepix effectively weakens the fungal cell wall, making it more susceptible to antifungal treatment. This targeted approach provides a promising avenue for treating infections caused by resistant fungal strains, particularly those that have developed resistance to conventional antifungal therapies [12]. Activity spectrum: C. albicans, C. tropicalis, C. parapsilosis, C. krusei, C. glabrata, C. dubliniensis, C. lusitaniae, and C. auris [12, 16]. Fosmanogepix has equally potent activities against fluconazole- and echinocandin-resistant Candida strains, and it showed no cross-resistance with azoles in Candida spp. [16].
Intracellular Structures
Mitochondrial Membrane Inhibitors. ATI-2307 (T-2307; arylamidines; phase I) inhibits mitochondrial function and causes the collapse of membrane potential in fungal cells [17]. Activity spectrum: C. albicans, C. tropicalis, C. parapsilosis, C. krusei, and C. glabrata [7, 12], including azole- and echinocandin-resistant Candida sp. [15].
Calcineurin Pathway Inhibitors. Cyclosporin A and Tacrolimus (FK506; pre-clinical) antifungal activity relies on binding to immunophilins forming a potent calcineurin inhibitor complex, which in turn interrupts the binding of calcineurin to calcium-calmodulin complex in fungal cytoplasm, thereby preventing the phosphatase activation. In fact, the calcineurin pathway mediates C. albicans tolerance to antifungal agents and virulence. The combination of cyclosporin A and tacrolimus with both azole and non-azole inhibitors of ergosterol biosynthesis has shown synergistic potential, enhancing the effectiveness of antifungal treatments. This is particularly evident when combined with fluconazole, increasing its fungicidal activity against Candida species like C. albicans, C. glabrata, and C. krusei. Additionally, inhibiting calcineurin activity by depleting heat shock protein 90 (Hsp90) through agents like efungumab (mycograb) and geldanamycin further enhances antifungal action. These combinations offer a promising strategy to combat resistant Candida infections by targeting multiple fungal survival pathways [7]. This mechanism was developed due to emerging fluconazole- and echinocandins-resistant C. albicans and C. glabrata strains and was confirmed to modulate the azole resistance.
De-acetyltransferases Inhibitors. MGCD290 (phase I) and Trichostatin A inhibit the fungal histone deacetylase 2 (Hos2) with an additional target through inhibition of the deacetylation of fungal Hsp90, involved in fungal stress adaptation. Activity spectrum: C. albicans, C. glabrata, and C. krusei (MGCD290 MIC of 0.5–16 µg mL−1); C. albicans when synergistically applied with fluconazole, itraconazole and terbinafine31. No ongoing trials on MGCD290 [12].
Acetyl-CoA Synthetase Inhibitors. AR-12 (phase I) has fungicidal activity at low doses against a range of Candida isolates, including C. albicans biofilms: C. albicans, C. glabrata, C. parapsilosis, C. tropicalis, and C. krusei (MIC of 2-4 µg mL−1) [7]. The combination of AR-12 with fluconazole has been shown to re-sensitize certain fluconazole-resistant strains of Candida albicans and Candida glabrata, restoring the effectiveness of the antifungal treatment. Furthermore, AR-12 has demonstrated continued activity against echinocandin-resistant strains that possess FKS mutations, a significant factor in drug resistance. Additionally, AR-12 has exhibited synergistic effects when combined with caspofungin, particularly in strains of C. glabrata that have developed resistance to caspofungin. Despite these promising results, there are currently no ongoing clinical trials focused on AR-12, leaving its broader potential in antifungal therapy unexplored [12].
Protein Synthesis Inhibitors. Sordarins represent a wide group of compounds having unique tetracyclic diterpene cores with a norbornene system and possessing antifungal activity. Sordarins inhibit protein synthesis at the elongation step of the translational cycle, binding to the fungal eukaryotic translation elongation factor 2 (eEF2)-ribosome complex, specifically at domain III [18]. Furthermore, sordarins target only the fungal eFF2 not affecting host translational systems. Sordarin (SL-2266) was first isolated from Sordaria araneosa and patented in 1969. Naturally occurring representatives of sordarins based on source of isolation can be grouped as follows: sordarin (Sordaria araneosa and Podospora pleiospora); sordarins C-F (Xylotumulus gibbisporus); sordaricin (Sordaria araneosa and Podospora pleiospora); hypoxysordarin (Sordaria araneosa, Podospora pleiospora, and Hypoxylon croceum); hydroxysordarin (Sordaria araneosa); xylarins a-c (Xylaria sp.); zofimarin (Zopfielle marina and Xylaria sp. Acra); isozofimarin (Xylaria sp. Acra); trichosordarin A (Trichoderma harzianum); moriniafungin (Morinia pestalozzioides, Curvularia hawaiiensis, and Setosphaeria rostrata); moriniafungins B-G (Curvularia hawaiiensis); GR 135402 (Graphium putredinis); BE31405 (Penicillium minioluteum) [18]. Activity spectrum of sordarins: C. albicans, C. glabrata, C. neoformans, C. parapsilosis, and C. tropicalis.
Extracellular Environment
Siderophores. VL-2397 (ASP2397), isolated from Acremonium persicinum, resembles the fungal ferrichrome siderophores, but preferentially chelates Al ions instead of Fe ions, and enters the fungal cells through specific transporters known as siderophore iron transporter 1 (Sit1) [12].
Antibiofilms. Turbinmicin, a highly oxidized type II polyketide produced by Micromonospora sp., inhibits Candida biofilm growth by disrupting fungal extracellular vesicle-mediated trafficking. It has demonstrated broad-spectrum fungicidal activity in vitro, including effectiveness against pan-resistant C. auris and multi-drug-resistant C. glabrata, with minimum inhibitory concentrations (MICs) of 0.5 µg/mL-1 or less. Furthermore, a recent study summarised the potential of turbinmicin to almost completely abrogate the production of biofilm vesicles in Candida sp., including C. tropicalis, C. glabrata, and C. auris [19]. EntV, a pre-pro-peptide produced by Enterococcus faecalis, inhibits C. albicans filamentation and biofilm formation, however, not possessing antifungal activity [19]. Another study discussed the potential of cationic dendritic compounds to be used as antibiofilm, specifically BDSQ024 dendrimer showed the highest activity (MIC of 16–32 µg mL−1) against biofilm formation of C. albicans [20].
Table 1 summarizes novel therapeutic preparations against Candida species, focusing on their drug class, half-life, spectrum of activity, and minimum inhibitory concentration (MIC). The table highlights new antifungal agents, many targeting resistant strains of Candida, such as azole- and echinocandin-resistant species, with varying methods of administration (oral or IV).
Alternative Anti-Candida Strategies
In addition to conventional and novel antifungal treatments, several natural approaches have shown promising results in the treatment of Candida sp. Infections [20]. Alternative antifungal strategies such as the use of an immunotherapy vaccine (NDV-3A), designed antimicrobial peptides (dAMPs), and complementary alternative medicine (CAM) [12].
Immunotherapeutic Vaccines
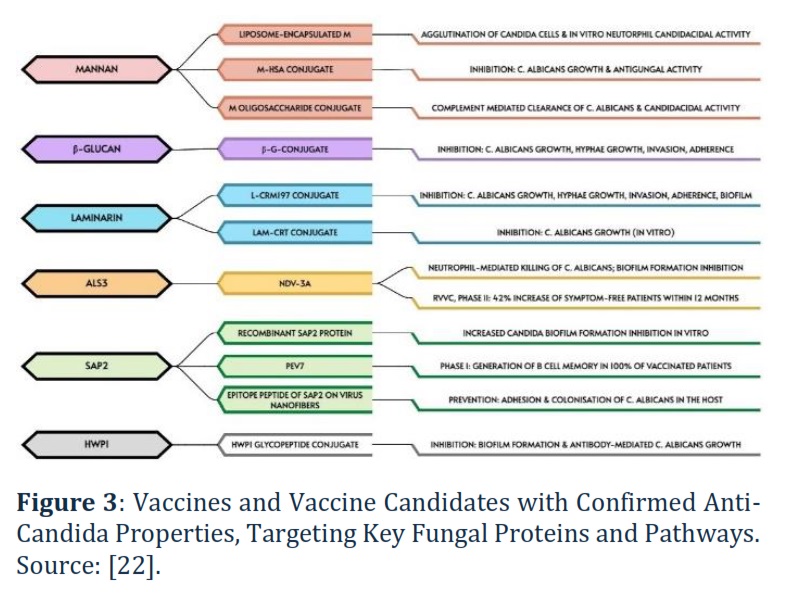
In 2017, Sui et al. [21] proposed a recombinant N-terminal Als3 vaccine (NDV-3A; rAls3p) as antifungal therapy for C. albicans-induced infections. The vaccine targets the N-terminus of Als3p, a hyphal-specific GPI cell wall protein, participating in adhesion, invasion, and biofilm formation [21]. NDV-3A was successfully implemented against RVVC in a phase II trial, resulting in a 42% increase in symptom-free patients within 12 months. A phase I clinical trial in humans of the ‘maternal’ vaccine, NDV-3 (rAls3p-N), confirmed its safety and ability to induce a robust immune response in patients. A Phase I clinical trial involving 48 healthy volunteers tested an innovative anti-Candida vaccine, PEV7 (rSap2t), which is formulated using a truncated and enzymatically inactive version of recombinant aspartyl proteinase-2 from Candida albicans. The results of this trial were highly promising, as 100% of the participants who received the vaccine developed specific and functional B cell memory. This immune response is critical for long-term protection against C. albicans infections, suggesting that PEV7 has the potential to be a highly effective vaccine for preventing candidiasis. The vaccine's ability to trigger such a robust immune memory response marks a significant step forward in the development of targeted fungal vaccines. Recently, Shukla et al. [22] have summarised potential anti-Candida vaccines candidates (Figure 3) focusing on the following targets: mannan, β-glucan, and laminarin, agglutinin-like sequence gene (Als3), secreted aspartyl proteinase (Sap2), heat shock protein (Hsp90), hyphally-regulated protein (Hyr1), hyphal wall protein (Hwp1), enolase (Eno), phospholipase (PLB), pyruvate kinase (Pk), fructose bisphosphate aldolase (Fba1), superoxide dismutase gene (Sod5), and malate dehydrogenase (Mdh1).
However, rationales are contributing against the vaccines' widespread use in R/VVC therapy, specifically: complicated selection of epitopes due to well-known phenotypic plasticity of C. albicans, a major R/VVC inducer; targeting host immune system to representative of local microflora (C. albicans) sounds to be non-fully justified; initiating immune response in immunocompromised patients seems to be challenging.
Figure 3, being modified from [22], demonstrates various antifungal strategies focusing on conjugates and formulations designed to combat Candida albicans. These include inhibition of fungal growth, prevention of biofilm formation, and enhanced immune-mediated clearance. The formulations shown have a wide range of applications, from direct antifungal activity to vaccine development, highlighting their therapeutic potential in both treatment and prevention of Candida infections. For instrance, NDV-3A leads to neutrophil-mediated killing of Candida albicans and prevents biofilm formation. In a clinical trial for recurrent vulvovaginal candidiasis (RVVC), this resulted in a 42% increase in symptom-free patients within 12 months, indicating significant therapeutic potential.
Designed Antimicrobial Peptides (dAMPs)
Antimicrobial peptides (AMPs) can be divided into natural and designed/synthesised antimicrobial peptides (dAMPs). Pérez-Laguna et al. [23] have summarised and classified existing AMPs with the antifungal potential against Candida sp. into six groups based on their origin (plants, human, insects and arachnids, bovine, amphibian skin, and filamentous fungi). Indeed, natural AMPs share the targets with conventional or novel antifungal drugs. Natural AMPs and their amino acid sequences serve as templates for designing functional peptides with improved activity, bioavailability, and stability in physiological conditions. dAMPs are artificially created molecules based on these natural AMPs and are designed to exhibit direct fungicidal activity, offering a targeted approach for combating fungal infections. These engineered peptides often feature enhanced properties, making them more effective and stable in clinical applications. dAMPs should be highly selective and have low hemolytic-cytotoxicity. Guevara et al. [24] investigated the efficiency of cecropin D-derived dAMPs (ΔM3 and ΔM4) against C. albicans, C. glabrata, C. tropicalis, and C. parapsilosis. Both dAMPs showed identical efficacy against C. albicans, C. tropicalis, and C. parapsilosis with minimal fungicidal concentrations (MFC99.9) of 1.6 and 3.1 µg mL−1, respectively [24]. In the case of C. glabrata, the fungicidal activity was determined only in ΔM4 with MFC99.9 of 50 µg mL−1. Furthermore, ΔM4 showed greater activity against biofilm formation in C. albicans with 100% inhibition at 12.5 µg mL−1, while ΔM3 was less effective, achieving a similar effect only at 100 µg mL−1. Woodburn et al. [25] evaluated the efficacy in vitro of four dAMPs (RP504, RP554, RP556, and RP557) for their potential use in RVVC therapy. MIC values of studied dAMPs against fluconazole-resistant C. albicans, C. glabrata, C. parapsilosis, and C. krusei ranged from 16 to > 32 µg mL−1, similar values were obtained for fluconazole-susceptible C. albicans, C. glabrata, and C. parapsilosis. Whereas in the case of fluconazole-resistant and susceptible C. tropicalis, MIC values ranged from 2-4 and 4-8 µg mL−1, respectively. RP554 and RP556 were more effective against C. lusitaniae (MIC of 2 µg mL−1), C. dubliniensis (MIC of 8 µg mL−1), C. famata (MIC of 4 µg mL−1) and C. kefyr (MIC of 2-4 µg mL−1), C. dubliniensis (MIC of 8 µg mL−1), C. sphaerica (MIC of 4 µg mL−1), respectively [25]. The similarity of dAMPs MIC and MFC values allowed them to state their fungicidal activity. Torres et al. [26] examined antifungal activity of three dAMPs (PNR20, PNR20-1, and 35409) against C. albicans, C. glabrata, C. parapsilosis, C. krusei, and C. tropicalis, and fluconazole-resistant C. auris. All three dAMPs were capable of 100% inhibition of C. krusei (MIC of 25 µg mL−1 for PNR20 and PNR20-1 and MIC of 100 µg mL−1 for 35409) and C. tropicalis (MIC of 50 µg mL−1 for PNR20 and 35409 and MIC of 100 µg mL−1 for PNR20-1). PNR20 and PNR20-1 inhibited growth (MIC of 100 µg mL−1) and biofilm formation (MBIC24 of 12.5 and 25 µg mL−1, respectively) of C. albicans, whereas growth of C. glabrata and C. tropicalis were inhibited by PNR20 only (MIC of 100 µg mL−1). Furthermore, dAMPs were capable of 50% biofilm formation inhibition in fluconazole-resistant C. auris (MBIC24 of 25, 25, and 12.5 µg mL−1 for PNR20, PNR20-1, and 35409, respectively) [26].
Complementary Alternative Medicine (CAM)
A recent study highlighted that up to 40% of patients suffering from recurrent vulvovaginal candidiasis (RVVC) resort to complementary and alternative medicine (CAM) to treat or prevent vulvovaginal candidiasis (VVC), despite the widespread availability of conventional antifungal agents (27). This significant reliance on CAM underscores the growing demand for alternative treatments, particularly among individuals seeking natural or holistic approaches to managing chronic or recurrent infections.
Complementary and alternative medicine encompasses a broad spectrum of natural products and practices, including essential oils, plant extracts, honey, and probiotics. Essential oils, such as tea tree oil, lavender oil, and oregano oil and other natural products are widely used for their antifungal, anti-inflammatory, and antimicrobial properties in both humans and animals [25, 27, 28].
Bee-Related Products
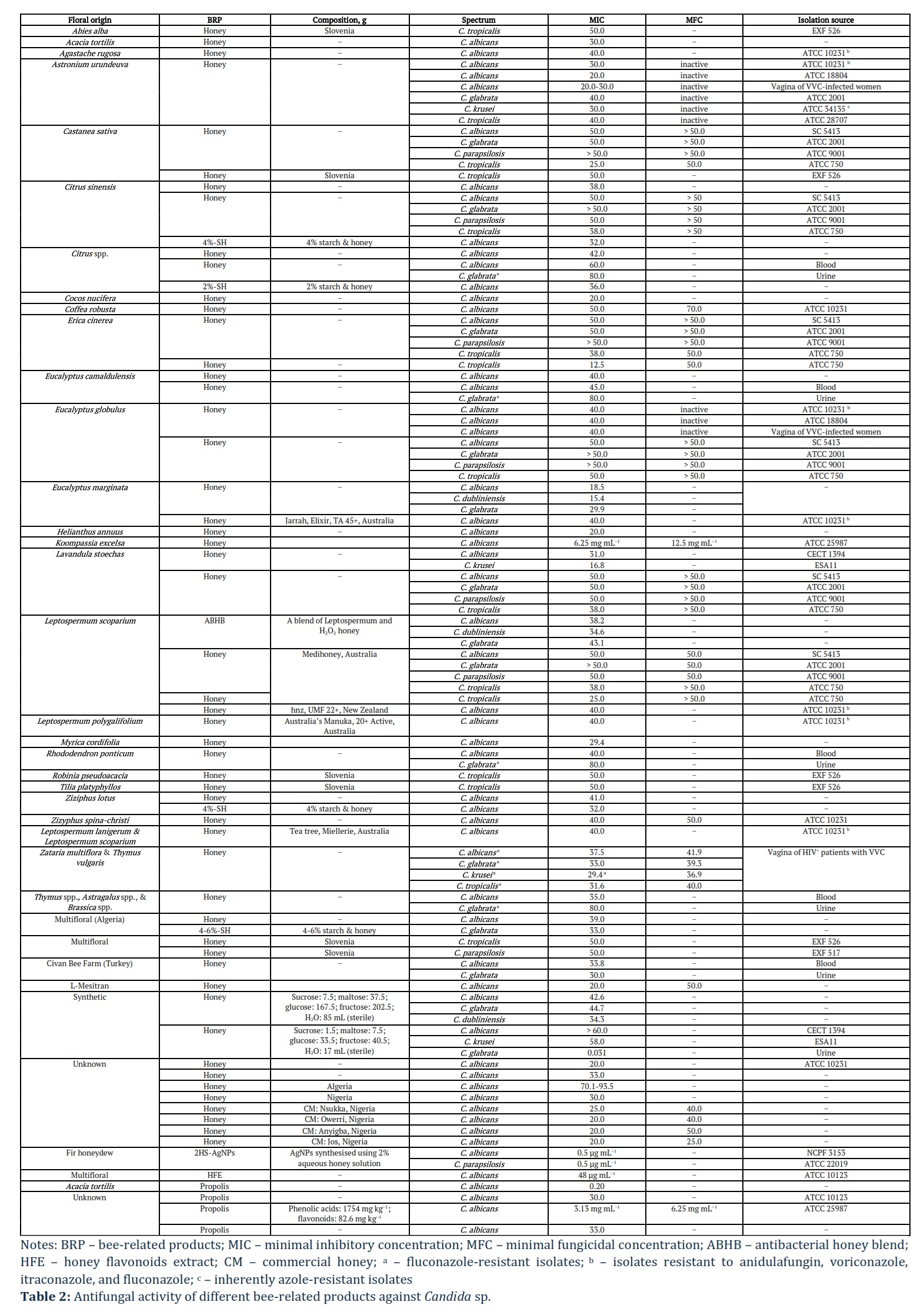
Honey is a supersaturated solution of sugars, comprising approximately 80% of its composition. Overall honey compounds responsible for antipathogenic activity can be classified as a) H2O2-dependent compounds produced by β-D-glucose transformation induced by bee’s glucose oxidase enzyme to gluconic acid and H2O2, where gluconic acid responsible for acidity and H2O2 for sporicidal activity; b) in turn non-H2O2-dependent compounds comprises of factors responsible for osmolarity and floral origin-dependent factors including methylglyoxal (MGO), melanoidin, phenolic compounds (flavonoids (flavanols) and phenolic acids), terpenes, and alkaloids; c) honey microbiota-derived compounds are divided to compounds of microbial (bacteriocins, kunkecin A, surfactants, non-ribosomal peptides (NRPs), and siderophores) and fungal (mycotoxins, β-lactams, surfactants, and siderophores) origin [29]. Furthermore, the floral origin of honey was confirmed to play a key role in its antimicrobial activity, whereas flavanols are known for their potent activity against pathogens, including resistant strains [29]. In addition, honey microbiota-derived compounds have also been proven to have antifungal activity: zygocin (mycotoxin) produced by Zygosaccharomyces bailii inhibited growth of C. albicans, C. krusei, and C. glabrata.
Fernandes et al. [30] examined six monocultural (chestnut: Castanea sativa; eucalyptus: Eucalyptus globulus; orange blossom: Citrus sinensis; rosemary: Lavandula stoechas; heather: Erica cinerea; Portugal) including one commercial (manuka: Leptospermum scoparium; Medihoney ®, Derma Sciences; Australia) honeys for the antifungal activity against C. albicans SC 5413, C. glabrata ATCC 2001, C. tropicalis ATCC 750, and C. parapsilosis ATCC 9001. Study findings revealed that C. tropicalis was the most susceptible to all honeys assessed: 25.0% w.v for chestnut honey and 38.0% w.v for eucalyptus, orange blossom, rosemary, heather, and manuka honeys [30]. The total phenolic concentrations in honeys were as follows (in mg of GAE/100 g): 104 for chestnut; 112 for eucalyptus; 34.8 for orange blossom; 55.4 for rosemary; 180 for heather; 163 for manuka. De Groot et al. [31] compared the effectiveness of L-Mesitran® Soft (40% medical-grade honey) and unprocessed local honey (Mook, the Netherlands) against clinical isolates of C. auris (KCTC 17809, VPCI 482/P/13, and 10-08-01-02), C. albicans (1082257609, 154, and AM 07/0267), C. glabrata (144, IFRC 649, and 5042020417), C. krusei (VPCI-1623/P/16, 197, and ATCC 6258), and C. parapsilosis (8655, 104. and VPCI 2289/P/16). Unprocessed honey was more effective in reducing all Candida sp., except for C. krusei, at MIC of 40.0% v.v [31].
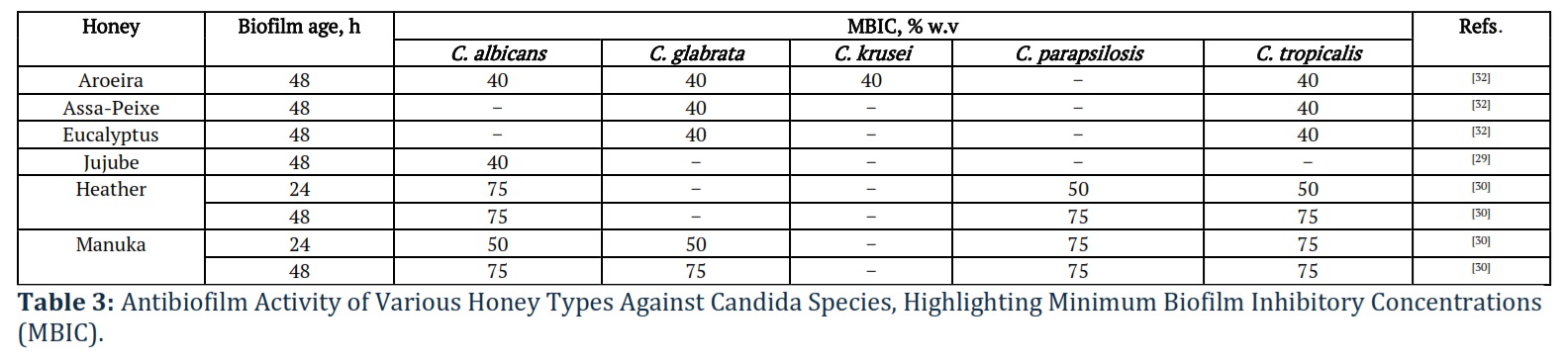
Lima et al. [32] tested antifungal and antibiofilm activities of three monofloral honeys (Aroeira: Astronium urundeuva; Eucalyptus: Eucalyptus globulus; Assa-Peixe: Vernonia polyanthes; Brazil) and one synthetic honey (in g: sucrose – 7.5; maltose – 37.5; glucose – 167.5; fructose – 202.5; 85 mL sterile water) against C. albicans ATCC 10231, C. albicans ATCC 18804, three clinical isolates of C. albicans from vaginal secretions of VVC-infected women, C. glabrata ATCC 2001, C. krusei ATCC 34135, and C. tropicalis ATCC 28707. In fact, Aroeira honey exhibited antifungal activity in relation to all isolates, Eucalyptus honey – only against C. albicans isolates, whereas Assa-Peixe and synthetic honey were inactive against Candida sp. [32]. Aroeira honey reduced 48-h-old biofilm biomass of C. albicans ATCC 10231, C. glabrata, C. krusei, and C. tropicalis by 35.5, 53.1, 45.9, and 41.4%, respectively. Antibiofilm activity of Eucalyptus and Assa-Peixe honey was observed only in relation to C. glabrata and C. tropicalis with a reduction of up to 34.9 and 43.0%, respectively. Synthetic honey reduced the biofilm biomass of C. tropicalis (by ~ 20%). The superior antifungal activity of Aroeira honey, followed by Eucalyptus honey, can be explained by the total phenolic concentrations of 99.7 ± 4.66 and 73.2 ± 2.16 GA/100 g, respectively, in contrast to 43.1 ± 3.63 and 8.85 ± 2.77 GA/100 g for Assa-Peixe and synthetic honeys. Dang et al.[33] explained the antifungal activity of monofloral coffee honey (Coffea robusta; Vietnam) against C. albicans ATCC 10231 by providing gene expression profile, which showed downregulation of EFG1 and overexpression of DPP3 playing an important role in the biofilm formation and yeast-to-hyphae transition as well as increased expression of Hsf1 responsible for C. albicans virulence and viability and increased sensitivity to antifungal treatment [33].
Bulgasem et al. [34] reported that lactic acid bacteria (LAB) isolated from seven honey samples, among which are Al-Hanon (Arbutus unedo; Libya), Al-Seder (Ziziphus spina‐christi; Yemen), multifloral Al-Maray and others, had similar inhibition tendency against C. albcans ATCC14053, C. parapsilosis ATCC22019, and C. tropicalis ATCC750. Furthermore, Lactobacillus curvatus of Al-Hanon and Pediococcus pentosaceus of Al-Maray had a stronger inhibitory effect on C. glabrata ATCC2001 compared to Lactobacillus plantarum of Al-Seder and Pediococcus acidilactici of Tualang [34].
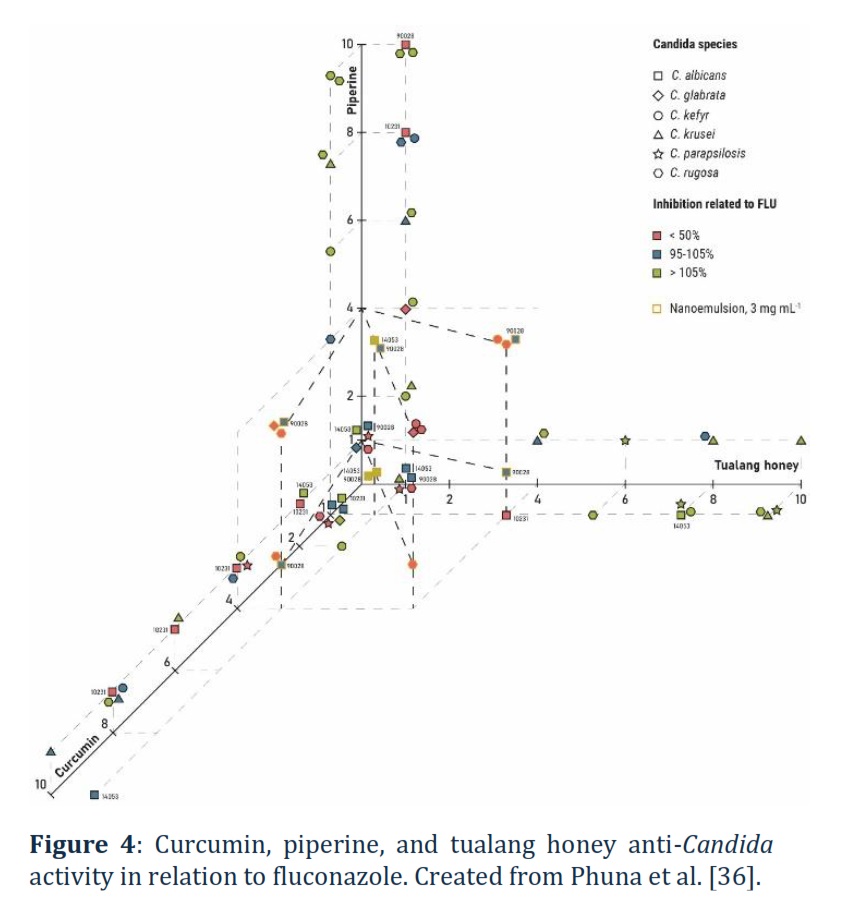
Czernel et al. [35] examined the potential of silver nanoparticles (AgNPs) synthesised using an aqueous solution of organic fir honeydew honey (Bieszczady Mountains, Poland) at different concentrations of 2, 10, and 20% against C. albicans NCPF 3153 and C. parapsilosis ATCC 22019. A 2% honey-synthesised AgNPs (2HS-AgNPs) showed the greatest inhibitory effect, whereas 10HS-AgNPs and 20HS-AgNPs showed no activity or even stimulated fungal growth [35]. In another study, Phuna et al. [36] examined curcumin and piperine as an additive to tualang honey (Koompassia excelsa; Malaysia) and in the form of nano-emulsions against C. albicans (ATCC 10231, ATCC 14053, and ATCC 90028), C. glabrata (ATCC 2001), C. kefyr (ATCC 2512), C. krusei (ATCC 14243), C. parapsilosis (ATCC 22019), and C. rugosa (ATCC 10571). Individual application of curcumin, piperine, and tualang honey had no superior antifungal activity against C. kefyr, C. parapsilosis, and C. rugosa compared to fluconazole, whereas their effect on C. albicans ATCC 90028 was similar to fluconazole. Indeed, individual application of curcumin was more efficient against C. albicans ATCC 10231 and C. glabrata, while, in the case of C. albicans ATCC 14053, piperine was more efficient than fluconazole (Figure 4) [36]. A single-blind clinical trial on reducing VVC symptoms (vaginal discharge, itching, burning, dyspareunia, and urinary problems) using 50% honey gel (Sabalan Mountains, Iran) reported a significant reduction of itching, burning, dyspareunia, and urinary problems by 94.3-98.1% [37]. Continued trials utilising the same honey gel showed promise in eliminating VVC symptoms such as vaginal discharge, vulva burning, and vaginal burning 52.8, 100, and 100%, respectively, within 8 days. Furthermore, a triple-blind study involving 70 non-pregnant women with VVC reported that a vaginal cream (produced by Armaghan Teb Laboratory, Tehran, Iran) containing yoghurt and honey was more effective in treating symptoms such as vaginal discharge, itching, dyspareunia, irritation, and dysuria compared to a 1% clotrimazole cream, specifically, the reductions were 92.3% vs. 72.4%, 93.3% vs. 68.8%, 76.0% vs. 59.1%, 92.6% vs. 62.5%, and 94.4% vs. 80.0% within 7 days, respectively. A double-blinded clinical trial involving 80 non-pregnant women with VVC compared the efficiency of honey (70% honey & 30% neutral cream) and 1% clotrimazole (Tehran Chemie, Iran) ointments in reducing VVC symptoms (inflammation, discharge, and itching) within 8 days and reported a 79.2-86.8% and 57.7-65.6% reduction for honey and clotrimazole groups [37].
Furthermore, authors evaluated the VVC reoccurrence incidences after 3 months, however, only 17 (38.6% of 44) and 15 (41.7% of 36) patients from honey and clotrimazole groups participated in the evaluation. Honey ointment reduced RVVC incidences compared to clotrimazole: 29.4% of 17 (or 11.4% of 44) vs. 53.3% of 17 (or 22.2% of 36) of responded patients, respectively [37]. Currently, researchers are conducting a clinical trial involving RVVC-infected (> 3 incidences per year) 252 women (non-pregnant, non-breastfeeding) with confirmed susceptible Candida sp. presence utilising intravaginal application of L-Mesitran (40% medical grade honey) [38].
Reviewed studies’ findings regarding the use of bee-related products for the treatment of fungal infections in women are compiled in Tables 2, 3.
Table 3 focuses on the ability of different types of honey to inhibit biofilm formation by various Candida species, including C. albicans, C. glabrata, and C. tropicalis. The table lists honey types, the age of the biofilm, and the minimum biofilm inhibitory concentration (MBIC) required to prevent biofilm development. For example, Manuka honey demonstrates strong antibiofilm activity, inhibiting biofilm formation in multiple Candida species.
Biotics
Boahen et al. [39] discussed the potential of probiotics, prebiotics, and postbiotics synergy against R/VVC, specifically biofilm formation. All three groups confer a health benefit to the host: probiotics are live microorganisms; postbiotics cover the broad range of compounds delivered to consumers in the form of non-viable compounds or cell fractions; prebiotics are defined as substrates that are selectively utilised by already existing host microorganisms. The most common probiotics used for R/VVC treatment are Lactobacillus and Bifidobacteria. Thus, utilising symbiotics (pre-, pro-, and postbiotics) aid in effective prevention (preventing pathogen attachment) or disruption (suppressing pathogen growth and inhibiting hyphae formation) of biofilm formation, specifically prebiotics stabilises vagina pH and enhances growth and viability of probiotics; latter in turn produces organic acids and antimicrobial peptides, postbiotics inhibit quorum sensing and decrease pathogen adhesion [39]. The role of biotics in R/VVC therapy will be thoroughly discussed in the following sections.
Although herbal topical preparations have gained popularity for treating vulvovaginal candidiasis (VVC), they are not currently recommended as a standard treatment. High-quality clinical evidence supporting the effectiveness of complementary and alternative therapies for recurrent vulvovaginal candidiasis (RVVC) remains limited. As a result, these alternative approaches are not advised for managing RVVC until more robust research is available to validate their efficacy [27].
Role of Probiotics in Women's Health as Adjunctive Therapy
Due to a reduction in the protective functions of the vaginal biotope, conditions are created for ascending infections of the female genital tract and foetal infection. When immunobiological barriers are compromised, there is a translocation of conditionally pathogenic bacteria into the bloodstream, leading to bacteraemia with internal organ infection.
Di Paola et al. [40] investigated the effects of probiotics on cervical cytological changes and HPV infection clearance. In this study, 24 women were given the Yakult® probiotic daily, containing the Lactobacillus paracasei Shirota strain, while 27 women served as the control group. After 3 months, the results showed that HPV disappeared in 25% of women taking the probiotic, compared to only 7.7% in the control group. After 6 months, the HPV clearance rates increased to 29.2% in the probiotic group and 19.2% in the control group. These findings suggest that probiotics may help in managing HPV infections, though further research is needed to fully understand how Lactobacillus-based products work and to determine the best formulations for promoting vaginal health.
The role of probiotics in enhancing vaginal health appears promising, particularly when used as an adjunct to traditional treatments for conditions like HPV infection or other vaginal disorders. Probiotics may help restore and maintain a healthy vaginal microbiome by introducing beneficial bacteria, which can balance the microbial environment and potentially prevent pathogenic overgrowths. However, beyond probiotics, other factors such as diet, hygiene, and sexual behavior also influence the vaginal microbiome. Maintaining this balance is critical for preventing infections and promoting overall vaginal health.Research shows that healthy vaginal ecosystems, known as community state types (CSTs), are typically dominated by Lactobacillus species, which maintain low anaerobic bacteria levels and a balanced immune response. Lactobacillus plays a key role in immunomodulation and restoring healthy microbiota in both the gut and vagina. Though rare cases of Lactobacillus bacteremia in immunocompromised patients exist, its benefits in reducing vaginal infections are well-documented, suggesting probiotic treatments could benefit these patients [41].
Lactobacillus regulates the vaginal microbiota by competing with harmful microbes and producing antimicrobial compounds, including acids that lower vaginal pH, hydrogen peroxide, and bacteriocin-like substances. Vaginal dysbiosis, seen in conditions like BV, CT, and VVC, is often linked to increased pH from reduced lactate levels.
Das et al. [42] studied vaginal lactic acid bacteria for preventing urogenital infections, isolating beneficial probiotics like Lactobacillus crispatus, L. gasseri, and L. vaginalis. Their probiotic supernatant inhibited pathogens, with L. crispatus showing the strongest antimicrobial effect due to lactic acid and hydrogen peroxide production. The study also demonstrated that lyophilized L. crispatus and PCFS retained their antibacterial properties over a long period of time [42].
It is also known that not all bacteria respond to the bactericidal activity of lactobacilli.
Studies have shown that certain bacterial strains can persist in the vaginal microbiota for extended periods, with some strains demonstrating the ability to remain for at least one year [43].
This persistence is significant because, while long-term stability of bacterial strains has been extensively observed in the gut, less is known about how bacterial populations behave in other parts of the body, such as the vagina. The stability and longevity of specific bacterial strains within the vaginal ecosystem remain relatively unexplored. However, early findings suggest that some strains may persist for multiple years, contributing to the long-term balance of the vaginal microbiota.
The vaginal microbiota is typically dominated by Lactobacillus species, which play a crucial role in maintaining a healthy environment through the production of lactic acid and other antimicrobial compounds. Despite the dominance of Lactobacillus, the vaginal microbiota also includes a diverse array of facultative and obligate anaerobes, which can coexist in this ecosystem. The composition of these microbial communities may remain stable over several menstrual cycles or exhibit fluctuations, depending on factors such as hormonal changes, sexual activity, and hygiene practices.
France et al. [43] sought to determine whether the bacterial strains within the vaginal microbiota could persist over a longer period, given the observed stability of some strains. Their research is part of ongoing efforts to better understand the dynamics of the vaginal microbiome, including how it maintains its balance, how persistent strains contribute to vaginal health, and how changes in microbial populations might influence conditions such as bacterial vaginosis (BV), yeast infections, or other reproductive health issues.
Tables and Figures
Understanding the long-term stability of vaginal bacterial strains could have important implications for probiotic therapies, personalized health interventions, and the prevention of recurring infections. Using metagenomic analysis, they identified stable strains with similar species of Lactobacillus iners, Lactobacillus crispatus, and Lactobacillus jensenii, with some participants experiencing significant shifts in relative abundance over time. De novo assembly revealed that stable strains had mutations that fixed within populations during the observation period, providing insights into the evolution of these bacteria. The results suggest that bacterial strains can persist in the vagina for extended periods, allowing them to evolve in the host's microenvironment. Thus, numerous studies indicate the prevalence of lactic acid bacteria, primarily lactobacilli, in vaginal bacterial communities of healthy women, with minimal diversity of anaerobic and conditionally pathogenic bacteria.
Advances in our understanding of the human microbiota will facilitate the faster development of new biopharmaceuticals using live bacteria to minimize or even eliminate the need for antibiotic therapy. Lactobacilli, owing to their Generally Recognized as Safe (GRAS) status, have become a popular and effective alternative to traditional antimicrobial treatments in the management of vaginal health. These beneficial bacteria are increasingly being utilized to prevent and treat chronic vaginitis, which encompasses conditions such as bacterial vaginosis (BV), yeast infections, and other forms of vaginitis caused by harmful pathogens. Moreover, the effectiveness of lactobacilli for prevention is also well established with long-term use, as there is a positive effect of lactobacillus-derived products with antibiofilm, antioxidant, pathogen-inhibitory, and immunomodulatory activities in the development of agents for vaginal infections [41].
Das et al. [42], as well as other researchers emphasize the importance of using probiotic lactobacilli for the prevention and treatment of vaginal infections. The study highlights the protective role of lactic acid bacteria against pathogens colonizing the urogenital area, demonstrating the potential of these probiotics in enhancing vaginal health. Additionally, different findings suggest that these probiotic formulations might be useful in preventing recurrent urinary tract infections, underscoring the significance of probiotic interventions in women's health.
Acknowledgements
This study was supported by the Ministry of Science and Higher Education of the Republic of Kazakhstan (grant number: BR21882248).
Author Contributions
All authors developed the concept, designed the methodology, collected and analyzed the data, and prepared the original draft of the manuscript.
The author(s) declare no conflict of interest.
![]() References
References
- Gonçalves B, Ferreira C, Alves CT, Henriques M, Azeredo J, Silva S. Vulvovaginal candidiasis: Epidemiology, microbiology and risk factors. Critical Reviews in Microbiology, (2016); 42(6): 905-927.
- Willems HME, Ahmed SS, Liu J, Xu Z, Peters BM. Vulvovaginal Candidiasis: A Current Understanding and Burning Questions. Journal of Fungi, (2020); 6(1): 27.
- San Juan Galán J, Poliquin V, Gerstein AC. Insights and advances in recurrent vulvovaginal candidiasis. PLOS Pathogens, (2023); 19(11): e1011684.
- Denning DW. Global incidence and mortality of severe fungal disease. The Lancet Infectious Diseases, (2024); 24(7): e428-e438.
- Bitew A, Abebaw Y. Vulvovaginal candidiasis: species distribution of Candida and their antifungal susceptibility pattern. BMC Womens Health, (2018); 18(1): 94.
- Logan A, Wolfe A, Williamson JC. Antifungal Resistance and the Role of New Therapeutic Agents. Current Infectious Disease Reports, (2022); 24(9): 105-116.
- Murphy SE, Bicanic T. Drug Resistance and Novel Therapeutic Approaches in Invasive Candidiasis. Frontiers in Cellular and Infection Microbiology, (2021); 11: 759408.
- Bouz G, Doležal M. Advances in Antifungal Drug Development: An Up-To-Date Mini Review. Pharmaceuticals, (2021); 14(12): 1312.
- Perlin DS. Cell Wall-Modifying Antifungal Drugs. In: Latgé JP, ed. The Fungal Cell Wall: An Armour and a Weapon for Human Fungal Pathogens. Current Topics in Microbiology and Immunology. 2020; 255-275. Springer International Publishing.
- Jospe-Kaufman M, Ben-Zeev E, Mottola A, Dukhovny A, Berman J, Carmeli S et al. Reshaping Echinocandin Antifungal Drugs To Circumvent Glucan Synthase Point-Mutation-Mediated Resistance. Angewandte Chemie, (2024); 136(9): e202314728.
- Silva-Dias A, Miranda IM, Branco J, Monteiro-Soares M, Pina-Vaz C, Rodrigues AG. Adhesion, biofilm formation, cell surface hydrophobicity, and antifungal planktonic susceptibility: relationship among Candida spp. Frontiers in Microbiology, (2015); 6: 205.
- Rauseo AM, Coler-Reilly A, Larson L, Spec A. Hope on the horizon: novel fungal treatments in development. Open Forum Infectious Diseases, (2020); 7(2): ofaa016.
- Wiederhold NP. Pharmacodynamics, Mechanisms of Action and Resistance, and Spectrum of Activity of New Antifungal Agents. Journal of Fungi, (2022); 8(8): 857.
- Bibi M, Murphy S, Benhamou RI, Rosenberg A, Ulman A, Bicanic T et al. Combining Colistin and Fluconazole Synergistically Increases Fungal Membrane Permeability and Antifungal Cidality. ACS Infectious Diseases, (2021); 7(2): 377-389.
- Scorzoni L, Fuchs BB, Junqueira JC, Mylonakis E. Current and promising pharmacotherapeutic options for candidiasis. Expert Opinion on Pharmacotherapy, (2021); 22(7): 887-888.
- Hoenigl M, Sprute R, Egger M, Arastehfar A, Cornely OA, Krause R et al. The Antifungal Pipeline: Fosmanogepix, Ibrexafungerp, Olorofim, Opelconazole, and Rezafungin. Drugs, (2021); 81(15): 1703-1729.
- McCarty TP, Pappas PG. Antifungal Pipeline. Frontiers in Cellular and Infectious Microbiology, (2021); 11: 732223.
- Shao Y, Molestak E, Su W, Stankevič M, Tchórzewski M. Sordarin— An anti-fungal antibiotic with a unique modus operandi. British Journal of Pharmacology, (2022); 179(6): 1125-1145.
- Ajetunmobi OH, Badali H, Romo JA, Ramage G, Lopez-Ribot JL. Antifungal therapy of Candida biofilms: Past, present and future. Biofilm, (2023); 5: 100126.
- Heredero-Bermejo I, Gómez-Casanova N, Quintana S, Soliveri J, Javier de la Mata F, Perez-Serrano J et al. In Vitro Activity of Carbosilane Cationic Dendritic Molecules on Prevention and Treatment of Candida Albicans Biofilms. Pharmaceutics, (2020); 12(10): 918.
- Sui X, Yan L, Jiang YY. The vaccines and antibodies associated with Als3p for treatment of Candida albicans infections. Vaccine, (2017); 35(43): 5786-5793.
- Shukla M, Chandley P, Rohatgi S. The Role of B-Cells and Antibodies against Candida Vaccine Antigens in Invasive Candidiasis. Vaccines, (2021); 9(10): 1159.
- Pérez-Laguna V, Barrena-López Y, Gilaberte Y, Rezusta A. In Vitro Effect of Photodynamic Therapy with Different Lights and Combined or Uncombined with Chlorhexidine on Candida spp. Pharmaceutics, (2021); 13(8): 1176.
- Guevara-Lora I, Bras G, Juszczak M, Karkowska-Kuleta J, Gorecki A, Manrique-Moreno M et al. Cecropin D-derived synthetic peptides in the fight against Candida albicans cell filamentation and biofilm formation. Frontiers in Microbiology, (2023); 13: 1045984.
- Woodburn KW, Clemens LE, Jaynes J, Joubert L-M, Botha A, Nazik H et al. Designed Antimicrobial Peptides for Recurrent Vulvovaginal Candidiasis Treatment. Antimicrobial Agents and Chemotherapy, (2019); 63(11): e02690-18.
- Torres R, Barreto-Santamaría A, Arévalo-Pinzón G, Firacative C, Gomez BL, Escandon P et al. In Vitro Antifungal Activity of Three Synthetic Peptides against Candida auris and Other Candida Species of Medical Importance. Antibiotics, (2023); 12(8): 1234.
- Gholamine B, Alwaily ER, Fakri Mustafa Y, Habash RT, Naghdi N, et al. An overview of the uses of propolis for oral health. Advancements in Life Sciences, (2023); 10(4): 515-524.
- Adeli OA, Prasad KD, Khalaf HA, Hjazi A, Hussien BM, Hussein HA, Naghdi N, Pirhadi M. Phytotherapy in sexual disorder: overview of the most important medicinal plants effective on sexual disorders. Advancements in Life Sciences, (2024); 10(4): 505-14.
- García-Tenesaca M, Navarrete ES, Iturralde GA, Villacres Granda IM, Tejera E, Beltran-Ayala P et al. Influence of Botanical Origin and Chemical Composition on the Protective Effect against Oxidative Damage and the Capacity to Reduce In Vitro Bacterial Biofilms of Monofloral Honeys from the Andean Region of Ecuador. International Journal of Molecular Science, (2018); 19(1): 45.
- Fernandes L, Ribeiro H, Oliveira A, Silva AS, Freitas A, Henriques M et al. Portuguese honeys as antimicrobial agents against Candida species. Journal of Traditional and Complementary Medicine, (2021); 11(2): 130-136.
- de Groot T, Janssen T, Faro D, Cremers NAJ, Chowdhary A, Meis JF. Antifungal Activity of a Medical-Grade Honey Formulation against Candida auris. Journal of Fungi, (2021); 7(1): 50.
- Lima WG, Moreira Brito JC, Stephanie da Cruz Nizer W, Sampaio de Assis DC. Antifungal, antibiofilm and anti-resistance activities of Brazilian monofloral honeys against Candida spp. Biocatalysis and Agricultural Biotechnology, (2022); 42: 102335.
- Dang QD, Nguyen TTT, Bui CD, Tran-Van H. Antifungal activity against Candida albicans biofilm of Coffea robusta monofloral honey from Vietnam. Notulae Scientia Biologicae, (2024); 16(1): 11600-11600.
- Bulgasem BY, Lani MN, Hassan Z, Yusoff WMW, Fnaish SG. Antifungal Activity of Lactic Acid Bacteria Strains Isolated from Natural Honey against Pathogenic Candida Species. Mycobiology, (2016); 44(4): 302-309.
- Czernel G, Bloch D, Matwijczuk A, Ciesla J, Kedzierska-Matysek M, Florek M et al. Biodirected Synthesis of Silver Nanoparticles Using Aqueous Honey Solutions and Evaluation of Their Antifungal Activity against Pathogenic Candida Spp. International Journal of Molecular Science, (2021); 22(14): 7715.
- Phuna ZX, Yu JKE, Tee JY, Shao QC, Tan NWH, Vijayabalan S et al. In Vitro Evaluation of Nanoemulsions of Curcumin, Piperine, and Tualang Honey as Antifungal Agents for Candida Species. Journal of Applied Biotechnology Reports, (2020); 7(3): 189-197.
- Banaeian S, Sereshti M, Rafieian M, Farahbod F, Kheiri S. Comparison of vaginal ointment of honey and clotrimazole for treatment of vulvovaginal candidiasis: A random clinical trial. Journal of Medical Mycology, (2017); 27(4): 494-500.
- Maastricht University Medical Center. A Randomized Controlled Trial: The Effect of a Medical Grade Honey Formulation (L-Mesitran) on Clinical Symptoms of Recurrent Vulvovaginal Candidiasis. clinicaltrials.gov, (2023). Accessed January 1, 2024. https://clinicaltrials.gov/study/NCT05367089
- Boahen A, Than LTL, Loke YL, Chew SY. The Antibiofilm Role of Biotics Family in Vaginal Fungal Infections. Frontiers in Microbiology, (2022); 13: 787119.
- Di Paola M, Sani C, Clemente AM, Iossa A, Perissi E, Castronovo G et al. Characterization of cervico-vaginal microbiota in women developing persistent high-risk Human Papillomavirus infection. Scientific Reports, (2017); 7(1): 10200.
- Chee WJY, Chew SY, Than LTL. Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health. Microbial Cell Factories, (2020); 19(1): 203.
- Das S, Konwar BK. Prophylactic application of vaginal lactic acid bacteria against urogenital pathogens and its prospective use in sanitary suppositories. International Microbiology, (2024); 27(1): 179-202.
- France M, Alizadeh M, Brown S, Ma B, Ravel J. Towards a deeper understanding of the vaginal microbiota. Nature Microbiology, (2022); 7(3): 367-378.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0