

Full Length Research Article
Bioinformatic Analysis of Oxygen Sensitivity in Arcobacter bivalviorum's Pyruvate:Ferredoxin Oxidoreductase (PFOR)
Abdulmajeed Alqurashi*
Adv. life sci., vol. 11, no. 4, pp. 843-848, November 2024
*– Corresponding Author: Abdulmajeed Alqurashi (aqurashi@taibahu.edu.sa)
Authors' Affiliations
[Date Received: 25/01/2024; Date Revised: 15/05/2024; Date Published: 15/10/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Arcobacter bivalviorum is an emerging foodborne pathogen particularly found in shellfish and other bivalves. This bacterium's metabolic versatility is highlighted by its possession of both the Pyruvate Dehydrogenase Complex (PDC) and Pyruvate:Ferredoxin Oxidoreductase (PFOR). The PFOR enzyme is crucial for linking glycolysis to the citric acid cycle. Previous studies of PFOR enzyme reveal its susceptibility to oxygen damage due to its reliance on Fe-S clusters for electron transfer.
Methods: The current study employs bioinformatic approaches to explore the oxygen sensitivity of A. bivalviorum's PFOR, comparing it with Desulfovibrio africanus and Moorella thermoacetica. The UniProt database was used to obtain the sequences of PFOR from D. africanus (CAA70873), A. bivalviorum (AXH11209), and M. thermoacetica (Q2RMD6). This comparative analysis sheds light on the structural similarities and differences between the enzymes, providing a deeper understanding of their functional mechanisms.
Result: PFOR enzyme is a heterodimer, with its functional subunits containing three [4Fe-4S]²⁺ clusters. Its exposed Fe-S clusters are vulnerable to oxygen and ROS, leading to enzyme inactivation. A comparison of PFOR sequences from D. africanus, M. thermoacetica, and A. bivalviorum reveals a crucial difference: the final 43 residues at the C-terminal of D. africanus are missing in the other two enzymes, depriving the M. thermoacetica and A. bivalviorum enzymes of a self-protective mechanism.
Conclusion: Pyruvate:ferredoxin oxidoreductase (PFOR) in A. bivalviorum lacks protection from oxidative damage due to the absence of the final 43 amino acids at the C-terminal.
Keywords: PFOR; Homology modeling; oxygen sensitivity; Foodborne pathogen
Introduction![]()
Arcobacter is a genus of bacteria that has gained attention in the field of food safety and public health [1]. This genus includes several species, such as Arcobacter butzleri, Arcobacter bivalviorum, and Arcobacter skirrowii, known for their potential to cause gastrointestinal infections in humans and animals. These species are commonly found in water environments, and their presence has been reported in a variety of foods, including meat, shellfish, poultry, and dairy products [2]. Arcobacters are capable of surviving under aerobic conditions, which distinguishes them from their close relatives in the Campylobacter genus [1]. The increasing detection of Arcobacter species in food and water sources highlights the need for more research to understand their pathogenicity, develop reliable detection methods, and establish effective control measures in food processing and handling to ensure public health safety. A. bivalviorum, in particular, is frequently isolated from food products and is considered an emerging foodborne pathogen [3].
A. bivalviorum is a relatively recently identified species within the Arcobacter genus, known for its presence in shellfish, particularly bivalves [4]. This species was first isolated and identified from mussels and oysters, highlighting its relevance in the context of marine and aquatic environments. A. bivalviorum, like some other Arcobacter species, is Gram-negative and exhibits both aerobic and microaerophilic characteristics [5]. This means it can grow in the presence of oxygen, but also thrives in environments with low oxygen levels. Its discovery has raised concerns in the field of food safety, particularly regarding shellfish consumption. The potential health risks associated with A. bivalviorum are not fully understood, but its close relation to other pathogenic Arcobacter species suggests a possible risk of gastrointestinal infections in humans [6]. Research is ongoing to better understand the pathogenicity, ecological niche, and transmission routes of this bacterium, as well as to develop effective detection methods for ensuring the safety of seafood products.
In bacteria, the conversion of pyruvate to acetyl-CoA can be facilitated by two main enzyme complexes: Pyruvate Dehydrogenase Complex (PDC) and Pyruvate:Ferredoxin Oxidoreductase (PFOR). PDC, like its eukaryotic counterpart, catalyzes the oxidative decarboxylation of pyruvate, producing acetyl-CoA, NADH, and CO₂ [7]. This process is crucial in aerobic and facultative anaerobic bacteria for connecting glycolysis to the citric acid cycle. In contrast, PFOR operates under anaerobic conditions, typically found in strict anaerobes. It catalyzes a similar reaction but transfers electrons to ferredoxin instead of NAD⁺, adapting to the electron transport chain’s requirements in anaerobic environments [7]. A. bivalviorum possesses a circular genome packed with 2,684,688 base pairs, with an average GC content of 28.1%. Remarkably, it possesses both the Pyruvate Dehydrogenase Complex (PDC) and the Pyruvate:Ferredoxin Oxidoreductase (PFOR) pathways, equipping it with exceptional metabolic versatility [4]. The presence of both PDC and PFOR provides this bacterium with the ability to adapt to diverse environments and utilize various sources of energy. This dual pathway ensures efficient conversion of pyruvate to acetyl-CoA, fueling the citric acid cycle and providing the essential energy for growth and survival.
Pyruvate:ferredoxin oxidoreductase (PFOR) is a microbial enzyme that catalyzes the reversible conversion of pyruvate into acetyl-CoA and carbon dioxide. It employs thiamine pyrophosphate (TPP), three [4Fe-4S] clusters, and coenzyme A (CoA) for this process. Notably, PFOR predates the multi-enzyme complex dehydrogenases [8] and utilize ferredoxin or flavodoxin (with a mid-point redox potential around -415 mV) as electron acceptors rather than NAD+ (mid-point redox potential of -320 mV). However, its reliance on Fe-S clusters for electron transfer makes it vulnerable to damage from molecular oxygen and oxygen radicals [9].
The purification and characterization of PFOR (Pyruvate:ferredoxin oxidoreductase) is challenging under aerobic conditions due to its vulnerability to oxygen damage. PFOR enzymes could be isolated from Helicobacter pylori using anaerobic buffers and dithiothreitol, the enzyme's activity is still lower than expected [10]. In Bacteroides thetaiotaomicron, the PFOR loses almost all its activity after just two hours of air exposure. Attempts to revive this activity in cell-free extracts using superoxide dismutase (SOD) or catalase were unsuccessful, suggesting that molecular oxygen, rather than superoxide or peroxide radicals, is responsible for the enzyme's inactivation [11]. However, isolating PFOR from the anaerobic organism Desulfovibrio africanus is comparatively easier due to its higher oxygen stability [12], enabling the successful determination of the enzyme’s structure [13]. There are two cysteine residues in the C-terminal region that is conserved amongst all species of Desulfovibrio. Site-directed mutagenesis of PFOR at either one of these two cysteines resulted in a protein that is more sensitive to oxygen and nonreactive to the reducing agent dithioerythritol(DTT). This confirms the critical role of a disulfide bond formation in protecting the enzyme under aerobic conditions [14]. The C-terminal region of D. africanus PFOR acts as a 'switch' through a disulfide bond. It covers the terminal solvent-exposed Fe-S cluster in the presence of oxygen, deactivating but safeguarding the enzyme. In more reducing conditions, this mechanism reverses, re-exposing the Fe-S cluster and reinstating enzyme activity [14]. The resilience of D. africanus PFOR to oxygen is entirely dependent on this C-terminal segment, as demonstrated by the increased susceptibility to oxygen damage in mutant proteins lacking this region [15].
In this study, bioinformatic approaches are employed to investigate the oxygen sensitivity of PFOR from A. bivalviorum. The sequence of A. bivalviorum PFOR compared to D. africanus, whose PFOR exhibits a moderate tolerance to oxygen, and Moorella thermoacetica, whose PFOR is highly sensitive to oxygen [12, 16]. Also, 3D models of A. bivalviorum PFOR is generated. Understanding the molecular basis of this oxygen sensitivity is crucial for developing strategies to control bivalviorum growth and its associated food safety risks.
Methods![]()
Sequence analysis:
The UniProt database was utilized to acquire the sequences of pyruvate ferredoxin oxidoreductase (PFOR) from three distinct organisms: D. africanus (UniProt Accession Number CAA70873), A. bivalviorum (UniProt Accession Number AXH11209), and M. thermoacetica (UniProt Accession Number Q2RMD6). To analyze the amino acid sequences of these PFOR enzymes, two different alignment tools from the European Molecular Biology Open Software Suite (EMBOSS) were employed. Clustal Omega, renowned for its efficacy in multiple sequence alignment, was used to identify regions of similarity across the sequences, providing insights into their functional, structural, and evolutionary relationships. Additionally, EMBOSS Needle was utilized, which employs the Needleman-Wunsch algorithm for global sequence alignment. This approach ensures a comprehensive alignment of the entire sequence length, facilitating a detailed comparison of the sequences in their entirety.
Active site determination of A. bivalviorum PFOR:
For the active site determination of Pyruvate:Ferredoxin Oxidoreductase (PFOR) from A. bivalviorum, InterPro was employed, an integrated resource that combines various protein signature databases [17]. This tool was instrumental in identifying potential active sites and critical residues that play a role in substrate binding and catalysis. The strength of InterPro lies in its ability to amalgamate data from multiple sources, facilitating the identification of conserved sequences and functional domains across different proteins. Such analysis is pivotal in predicting protein functions, especially in elucidating the interactions between proteins and their substrates during catalytic actions. Through InterPro's comprehensive analysis, we successfully identified specific amino acid residues and domains in the A. bivalviorum PFOR that are likely essential for its enzymatic function.
Modelling the structure of A. bivalviorum PFOR:
To elucidate the structural characteristics of Pyruvate:Ferredoxin Oxidoreductase (PFOR) from A. bivalviorum, advanced computational modeling techniques were employed. The protein sequence of A. bivalviorum PFOR was used to model the structure of the enzyme using the PHYRE-2 and SWISS-MODEL, both highly regarded in the field of protein homology and analogy recognition [18]. The model was generated with reference to the structure of the pyruvate-ferredoxin oxidoreductase from M. thermoacetica, as referenced in the RCSB Protein Data Bank (PDB ref. 6CIQ) [16], with 52% i.d. and 100% confidence.
Protein visualization and analysis:
PyMOL, a powerful molecular visualization system, was utilized to analyze the three-dimensional structure of Pyruvate:Ferredoxin Oxidoreductase (PFOR) from A. bivalviorum. This advanced visualization tool enabled us to perform detailed 3D alignments, particularly focusing on two key domains of the enzyme: the 4Fe-4S ferredoxin, an iron-sulfur binding domain, and the Thiamine pyrophosphate (TPP family), a pyrimidine (PYR) binding domain. The comparative analysis involved aligning these domains against the corresponding domains of pyruvate-ferredoxin oxidoreductase from M. thermoacetica. Such a comparison is crucial for understanding the structural similarities and differences between these enzymes, which, in turn, can shed light on their functional mechanisms. PyMOL's sophisticated rendering and alignment capabilities allowed us to observe the spatial arrangement of amino acids and the overall architecture of these domains with high precision.
Results![]()
Sequence analysis:
Pyruvate:ferredoxin oxidoreductase (PFOR) is a heterodimer consisting of two functional subunits, each containing one molecule of TPP and three [4Fe-4S]2+ clusters [15]. It is likely that molecular oxygen and reactive oxygen species (ROS) are damaging the solvent-exposed Iron-sulfur (Fe–S) clusters of these enzymes, resulting in an inactive enzyme [9]. protein sequence alignment of PFOR from D. africanus (D.Apor), M. thermoacetica (M.Tpor) and A. bivalviorum (A.Bpor), shows one key difference between these enzymes; the final 43 residues at the C-terminal end of D.Apor are missing in M.Tpor and A.Bpor (Figure1). C-terminal region has two cysteine residues that is conserved amongst all species of Desulfovibrio. It has been shown that a di-sulphide bond mediated conformational switch that acts to occlude the terminal solvent-exposed Fe-S cluster under oxidizing conditions (inactivating the enzyme but limiting oxidative damage) and re-expose it under reducing conditions (reactivating the enzyme) [12, 14]. Without the 43 residue C-terminal region, the M.Tpor and A.Bpor enzymes lack any kind of self-contained protective mechanism against damage by molecular oxygen or oxidative radicals. For the proper function of the enzyme under hypertoxic conditions it seems M. thermoacetica and A. bivalviorum must rely on external factors for protection.
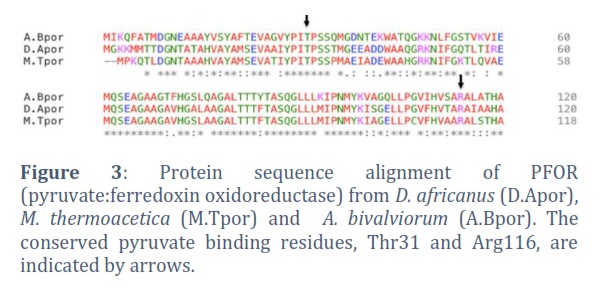
To investigate the relationship between the PFOR enzymes from M. thermoacetica and A. bivalviorum, EMBOSS Needle is used for pairwise sequence alignment. The alignment of both proteins M. thermoacetica PFOR and A. bivalviorum PFOR indicated a higher degree of similarity (68.8%, 834/1212) between these two proteins (Figure2). The presence of gaps was relatively minimal (4.3%, 52/1212), suggesting a substantial degree of conservation in the overall sequence, despite the noted differences. Figure3 shows the conserved pyruvate binding residues, Thr31 and Arg116. The conserved pyruvate binding residues, Thr31 and Arg116, play a crucial role in the functionality of certain enzymes. These residues are often found in the active sites of enzymes where they participate directly in the binding and catalysis of pyruvate. Threonine, being a polar amino acid, can form hydrogen bonds with the pyruvate molecule, aiding in its proper orientation and stabilization within the active site. Arginine, with its positively charged side chain, can interact with the negative charges of pyruvate, further stabilizing the enzyme-substrate complex.
Modelling the structure of A. bivalviorum PFOR:
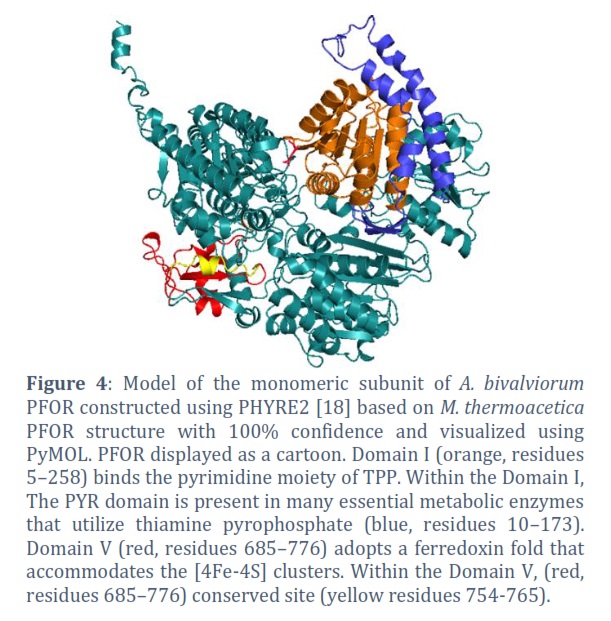
A search of the RCSB Protein Data Bank [19] for protein structures for similar pyruvate:ferredoxin reductase in other anaerobic or microaerobic organisms yielded a structure; the functionally identical pyruvate:ferredoxin reductase (PFOR) from the anaerobe M. thermoacetica (referred to as M.Tpor( [16]. Model of A. bivalviorum PFOR (referred to as A.Bpor ( constructed using PHYRE2 based on M.Tpor structure with 100% confidence and visualized using PyMOL.
The A. Bpor enzyme is an alpha 2 homodimer. Each monomeric subunit is composed of six domains. Domain, I bind the pyrimidine moiety of TPP. Domain II is known as (TKC) domain. Domain III is a CoA-binding domain. Domain IV is a structured linker that connects domain III and domain V. Domain V adopts a ferredoxin fold that accommodates the iron sulphur clusters that are distal and medial to the TPP molecule. Domain VI binds the pyrophosphate moiety of TPP and the proximal iron sulphur cluster. Figure 4 shows the ribbon figure of the monomeric subunit. Domain I is red while Domain V is orange.
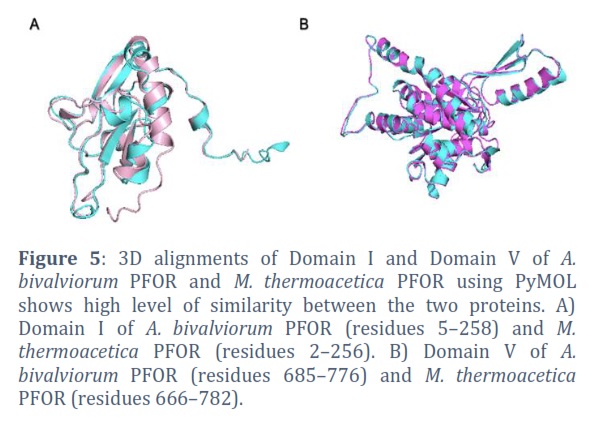
Figure 5 shows three-dimensional alignments of Domain I and Domain V in A.Bpor enzyme and M.Tpor enzyme conducted using PyMOL, revealing a significant degree of resemblance between these two enzymes. The comparison involves Domain I of A. bivalviorum PFOR, encompassing amino acids 5 to 258, and that of M. thermoacetica PFOR, covering amino acids 2 to 256. For Domain V, the alignment includes amino acids 685 to 776 from A. bivalviorum PFOR and amino acids 666 to 782 from M. thermoacetica PFOR.
Figures & Tables


Pyruvate:ferredoxin oxidoreductase (PFOR) is an important enzyme found in many microaerobic/anaerobic bacteria. It plays a crucial role in converting pyruvate to acetyl-CoA, a key intermediate in energy production. The acetyl-CoA produced can then enter various metabolic pathways, including the synthesis of fatty acids and the generation of ATP through the citric acid cycle. PFOR typically requires several cofactors to function, including thiamine pyrophosphate (TPP), coenzyme A (CoA), and iron-sulfur (Fe-S) clusters. The Fe-S clusters are particularly important for electron transfer during the catalytic process. The enzyme's structure allows these cofactors to interact efficiently with the pyruvate substrate. Iron-sulfur (Fe-S) clusters are versatile cofactors present in all life forms, utilized for various biochemical roles including electron transfer, redox reactions, and sensing of iron (Fe) or/and oxygen (O2) [20]. For instance, E. coli is estimated to contain over 150 different enzymes that incorporate Iron-sulfur clusters [21]. The widespread occurrence of these clusters is attributed to their stability in anaerobic conditions and the environmental availability of their basic components, making them ideal redox catalysts for early life before the advent of an oxygen-rich atmosphere. Iron-sulfur clusters come in several configurations, ranging from simple structures with a single iron atom bound by four cysteine residues to more complex and prevalent forms like [2Fe-2S] (rhombic) and [4Fe-4S] (cubic) clusters [9]. Their functionality stems from the ability of each iron atom to alternate between the ferrous (Fe2+) and ferric (Fe3+) states, facilitating efficient electron transfer or binding to oxygen (O2) or nitrogen (N2) atoms in organic molecules [9]. These clusters are vulnerable to damage from oxidative radicals or/and oxygen (O2) molecules. PFOR is generally sensitive to oxygen, which can damage the Fe-S clusters essential for its activity. This sensitivity is a key reason why PFOR is predominantly found in anaerobic organisms. The enzyme's structure and function can be significantly altered in the presence of oxygen, leading to inactivation. Different PFOR enzymes exhibit variations in active-site solvent accessibility. For instance, M.Tpor possesses an active site that is open, while the active site of D.Apor is shielded from the solvent. This study underscores the vulnerability of PFOR enzymes to molecular oxygen and reactive oxygen species (ROS), which can inactivate the enzyme by damaging its solvent-exposed Iron-sulfur (Fe–S) clusters. This vulnerability is particularly pronounced in M.Tpor and A.Bpor, which lack the C-terminal region present in D.Apor. This region in D.Apor, containing two conserved cysteine residues, is crucial for forming a disulfide bond that protects the enzyme under oxidizing conditions. The absence of this protective C-terminal region in M.Tpor and A.Bpor suggests their increased susceptibility to oxidative damage, highlighting the necessity of external protective mechanisms in these species.
Sequence and structural analysis show a high degree of similarity between M.Tpor and A.Bpor, with minimal gaps indicating substantial conservation. This conservation is particularly notable in the pyruvate binding residues, Thr31 and Arg116, which are crucial for the enzyme's functionality. These residues' ability to form hydrogen bonds and ionic interactions with pyruvate underscores their importance in stabilizing the enzyme-substrate complex. The structural modelling of A.Bpor, based on the M.Tpor structure, reveals an alpha 2 homodimer configuration with six distinct domains. Each domain plays a specific role, from TPP binding to accommodating iron-sulfur clusters crucial for the enzyme's function. The 3D alignments of Domain I and Domain V between A.Bpor and M.Tpor, visualized using PyMOL, demonstrate a significant resemblance, indicating evolutionary conservation and possibly similar functional mechanisms.
PFOR is a key enzyme in the metabolism of anaerobic organisms, facilitating the conversion of pyruvate to acetyl-CoA under oxygen-limited conditions. Its study is important for understanding fundamental biological processes and has potential applications in various scientific and industrial fields. It plays a significant role in global carbon and energy cycles. It is involved in the breakdown of organic compounds in anaerobic environments, such as sediments and the guts of animals, including humans. Understanding the mechanism and structure of PFOR is important for various fields of research, including biochemistry, microbiology, and biotechnology. Insights into PFOR can aid in the development of biocatalysts for industrial processes, the study of metabolic diseases, and the understanding of microbial ecology in anaerobic environments.
The author declare that there is no conflict of interest regarding the publication of this paper.
![]() References
References
- Hsu TT, Lee J. Global Distribution and Prevalence of A rcobacter in Food and Water. Zoonoses and public health, (2015); 62(8): 579-89.
- Kanaan MH. Prevalence, resistance to antimicrobials, and antibiotypes of Arcobacter species recovered from retail meat in Wasit marketplaces in Iraq. International Journal of One Health, (2021); 7(1): 142-150.
- DEMİRCİ Ü: Cronobacter Sakazakii: An Emerging Pathogen In Food Safety. International Journal of Health Administration and Education Congress (Sanitas Magisterium), (2020); 6(1): 54-63.
- Miller WG, Yee E, Bono JL. Complete genome sequence of the Arcobacter bivalviorum type strain LMG 26154. Microbiology Resource Announcements, (2018); 7(12): 10-1128
- Levican A, Collado L, Aguilar C, Yustes C, Diéguez AL, Romalde JL, Figueras MJ. Arcobacter bivalviorum sp. nov. and Arcobacter venerupis sp. nov., new species isolated from shellfish. Systematic and applied microbiology, (2012); 35(3): 133-8.
- Mateus C, Martins R, Luis A, Oleastro M, Domingues F, Pereira L, Ferreira S. Prevalence of Arcobacter: from farm to retail–A systematic review and meta-analysis. Food Control, (2021); 128(10): 108177.
- Katsyv A, Schoelmerich MC, Basen M, Müller V. The pyruvate: ferredoxin oxidoreductase of the thermophilic acetogen, Thermoanaerobacter kivui. FEBS Open Bio, (2021); 11(5): 1332-42.
- Weerakoon DR, Olson JW. The Campylobacter jejuni NADH: ubiquinone oxidoreductase (complex I) utilizes flavodoxin rather than NADH. Journal of bacteriology, (2008); 190(3): 915-25.
- Imlay JA. Iron‐sulphur clusters and the problem with oxygen. Molecular microbiology, (2006); 59(4): 1073-82.
- Hughes NJ, Chalk PA, Clayton CL, Kelly DJ. Identification of carboxylation enzymes and characterization of a novel four-subunit pyruvate: flavodoxin oxidoreductase from Helicobacter pylori. Journal of Bacteriology, (1995); 177(14): 3953-9.
- Pan N, Imlay JA. How does oxygen inhibit central metabolism in the obligate anaerobe Bacteroides thetaiotaomicron. Molecular microbiology, (2001); 39(6): 1562-71.
- Pieulle L, Guigliarelli B, Asso M, Dole F, Bernadac A, Hatchikian EC. Isolation and characterization of the pyruvate-ferredoxin oxidoreductase from the sulfate-reducing bacterium Desulfovibrio africanus. Biochimica et Biophysica Acta (BBA)-Protein Structure and Molecular Enzymology, (1995); 1250(1): 49-59.
- Chabrière E, Charon MH, Volbeda A, Pieulle L, Hatchikian EC, Fontecilla–Camps JC. Crystal structures of the key anaerobic enzyme pyruvate: ferredoxin oxidoreductase, free and in complex with pyruvate. Nature structural biology, (1999); 6(2): 182-90.
- Vita N, Hatchikian EC, Nouailler M, Dolla A, Pieulle L. Disulfide bond-dependent mechanism of protection against oxidative stress in pyruvate-ferredoxin oxidoreductase of anaerobic Desulfovibrio bacteria. Biochemistry, (2008); 47(3): 957-64.
- Pieulle L, Magro V, Hatchikian EC. Isolation and analysis of the gene encoding the pyruvate-ferredoxin oxidoreductase of Desulfovibrio africanus, production of the recombinant enzyme in Escherichia coli, and effect of carboxy-terminal deletions on its stability. Journal of bacteriology, (1997); 179(18): 5684-92.
- Chen PY-T, Aman H, Can M, Ragsdale SW, Drennan CL. Binding site for coenzyme A revealed in the structure of pyruvate: ferredoxin oxidoreductase from Moorella thermoacetica. Proceedings of the National Academy of Sciences, (2018); 115(15): 3846-51.
- Paysan-Lafosse T, Blum M, Chuguransky S, Grego T, Pinto BL, Salazar GA, et al. InterPro in 2022. Nucleic acids research, (2023); 51(D1): D418-D427.
- Kelley LA, Sternberg MJ. Protein structure prediction on the Web: a case study using the Phyre server. Nature protocols, (2009); 4(3): 363-71.
- Berman H, Henrick K, Nakamura H. Announcing the worldwide protein data bank. Nature structural & molecular biology, (2003); 10(12): 980-980.
- Py B, Barras F. Building Fe–S proteins: bacterial strategies. Nature Reviews Microbiology, (2010); 8(6): 436-46.
- Roche B, Aussel L, Ezraty B, Mandin P, Py B, Barras F. Reprint of: Iron/sulfur proteins biogenesis in prokaryotes: formation, regulation and diversity. Biochimica et Biophysica Acta (BBA)-Bioenergetics, (2013); 1827(8-9): 923-37.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()