

Full Length Research Article
Influence of diverse pH and temperatures on the gene expression of toxin-antitoxin systems in Klebsiella pneumoniae
Fatima J. Hassan*, Mohammed F. Al-Marjani, Intesar N. Khelkal
Adv. life sci., vol. 11, no. 4, pp. 815-821, November 2024
*– Corresponding Author: Fatima J. Hassan (fatimajh2022@uomustansiriyah.edu.iq)
Authors' Affiliations
[Date Received: 24/10/2023; Date Revised: 29/05/2024; Date Published: 15/10/2024]
Editorial Expression of Concern:
Editorial Note:
27 May 2025: You are viewing the latest version of this article having correction in article info (metadata) section.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: The type II toxin-antitoxin system consists of tightly linked genetic units that produce a toxin and its specific antitoxin. Under typical circumstances, the antitoxin neutralizes the toxic effects caused by the toxin. However, the toxin-antitoxin systems during stress regulate bacterial growth and persistence by affecting translation or DNA replication. This study aimed to assess how variations in temperature and pH stress influence the regulation of type II toxin-antitoxin gene expression within Klebsiella pneumoniae isolates.
Methods: Sixty-five Klebsiella pneumoniae isolates were identified by the VITEK®2 compact system and confirmed identity by Polymerase Chain Reaction assay by amplified rpoB gene (Housekeeping gene). The isolates were subjected to Reverse Transcriptase-polymerase chain Reaction assay to determine the transcriptional variations of the type II toxin-antitoxin system genes at diverse temperature and pH values.
Results: The results of the current study demonstrated higher expression levels of the antitoxin hipB gene at (37℃, 30℃, 44℃, pH 7, pH 9), mqsR toxin gene at (37℃, 30℃, pH 7, pH 5, pH 11) and relE toxin gene at (pH 5, pH 11).
Conclusion: These findings suggest the possible influence of pH and temperature on toxin-antitoxin systems, which might enhance bacterial antibiotic tolerance and persistence.
Keywords: Toxin-antitoxin systems; pH stress; Temperature stress; Klebsiella pneumoniae; Gene expression; Reverse Transcriptase PCR
Introduction![]()
Klebsiella (K.) pneumoniae is a gram-negative bacterium of the Enterobacteriaceae family that grows best at pH 7.2 and at temperatures ranging from 35℃ to 37℃ [1–3]. It is a prevalent opportunistic pathogen in humans, causing both hospital-acquired and community-acquired infections [4,5]. K. pneumoniae is one of the most antibiotic-resistant microorganisms and is rapidly developing multi-drug resistant (MDR) strains [6]. These strains acquire antibiotic resistance more swiftly than many other bacteria, posing significant concerns for patients due to the associated high risks of mortality and morbidity [6,7].
Toxin-antitoxin (TA) Systems are genetic loci within an operon found in bacterial plasmids, chromosomes, and phages that encode a stable toxin and a liable antitoxin [8]. TA systems were first discovered in plasmids in 1983 and recognized for their capability to stabilize plasmids through the post-segregation killing of plasmid-free daughter cells [9]. TA systems play roles not only in regular physiological processes but also in pathogenic mechanisms. Normally, antitoxin, whether protein or RNA-based, neutralizes the toxin’s harmful effects. However, under stress conditions, such as those induced by the guanosine tetraphosphate (ppGpp) signaling cascade, the antitoxin is degraded. This results in the release of the toxin, typically a protein that targets various intracellular molecules, inhibiting processes like translation and DNA replication [10,11].
The TA systems have been categorized into eight classes based on the mechanism and type of antitoxin action: (type I TA systems, type II TA systems, type III TA systems, type IV TA systems, type V TA systems, type VI TA systems, type VII TA systems, and type VIII TA systems) [12]. The Type II TA system represents the most prevalent configuration, wherein both the liable antitoxin protein and the more stable toxin protein are co-encoded within the same operon. The antitoxin mitigates the toxic effects of the toxin through direct protein-protein interactions, thereby neutralizing its harmful impact [13].
Pathogenic bacteria were shown to have more type II TA systems than non-pathogenic bacteria [14]. TA systems are known to offer assistance not only in general stress response but also in apoptosis, post-segregational killing, persister cell production, biofilm formation, phage abortive infection, and aiding in the transfer of antibiotic resistance genes through TA systems integrated within mobile genetic elements [15]. Persistence refers to the ability of certain genetically similar sub-populations of cells to withstand harsh environmental conditions like exposure to antibiotics or scarcity of nutrients by temporarily transitioning into a dormant state characterized by reduced metabolic activity and slowed growth rate, thereby ensuring bacterial survival and enhanced tolerance towards antibiotics [16].
Bacterial metabolism is impacted by a wide range of environmental stresses, including temperature, pH, nutrition availability, and salinity which impact not just growth and survival but also bacterial pathogenicity [17,18]. Antibiotic exposure and oxidative stress might cause the bacterial SOS response to be triggered [19]. The current study aimed to assess how pH levels and temperatures affect the gene expression of the type II TA system and the proliferation of K. pneumoniae isolates.
Methods![]()
Samples collection, and growth conditions of bacteria
Sixty-five K. pneumoniae isolates were acquired from many hospitals situated in Baghdad, Iraq, and collected from different clinical sources such as urine, blood, sputum, stool, ear swabs, burns, and wounds. These isolates were incubated in brain heart infusion (BHI) broth (TM media/ India) and on Luria-Bertani (LB) agar at a temperature of 37℃ for 24 h [20].
Identification of Klebsiella pneumoniae isolates
All isolates of K. pneumoniae were recognized using VITEK®2 compact system (BioMérieux/ France) and their identity was subsequently verified by Polymerase Chain Reaction (PCR) assay by amplified rpoB gene (Housekeeping gene) in DNA thermal cycler (DLab/ China). Reaction components in Final volume (25µl) which contains Green Master Mix 2x (Promega/ Canada) (12.5μl), forward primers (1µl), reverse primers (1µl), template DNA (5μl), nuclease-free water (5.5µl) (Promega/ Canada). PCR reaction conditions comprised of 95℃ (5min) to initial denaturation, followed by thirty cycles of 95℃ (30s) to denaturation, 57℃ (30s) to annealing, 72℃ (45s) to extension, and 72℃ (5 min) to final extension [5].
Determination of the impact of temperatures and pH levels on growth of Klebsiella pneumoniae isolates
The temperatures and pH values impact on K. pneumoniae isolates were assessed by cultivation and incubation at 37℃ for 24 h on Luria-Bertani (LB) agar. A few of pure colonies obtained from an overnight culture were introduced into 5 ml of BHI broth and incubated for 24 h at different temperatures (30℃, 37℃, 44℃). Moreover, a select number of colonies from an overnight culture were introduced into 5 mL of BHI broth with varying pH levels of 5, 7, 9, and 11, previously measured with a pH meter (LKB/Sweden) and incubated for 24 h at 37℃. The bacterial culture optical density was measured utilizing a spectrophotometer (Optima/Japan) at 600nm wavelength (OD600) for different time intervals (0, 8 h, and 24 h) [21]. The bacterial culture was subjected to a tenfold dilution process to achieve an optical density (OD) ranging from 0.08 to 0.6 at a wavelength of 600nm. The cells were serially diluted and plated onto nutritional agar (HiMedia/India). Following an incubation period of 24 h at a temperature of 37℃, the resulting colonies were meticulously enumerated to assess the growth and viability of the bacterial culture.
Type II TA system genes Identification
DNA extraction by using the boiling method [22]. Uniplex PCR was used to amplify (mqsR/mqsA, hipA/hipB, relE/relB, mazF/mazE) eight genes of type II TA system within K. pneumoniae isolates as illustrated in (Table 1). The PCR analysis was conducted using a DNA-thermocycler (DLab/China) to demonstrate the presence of genes related to type II TA systems. Thermal cycling included 94℃ (5 min) for an initial denaturation stage followed by thirty-five cycles (94℃ (1 min) for denaturation, 58.6℃ (45s) for annealing, and 72℃ (1 min) for extension with 72℃ (10 min) at a final extension step.
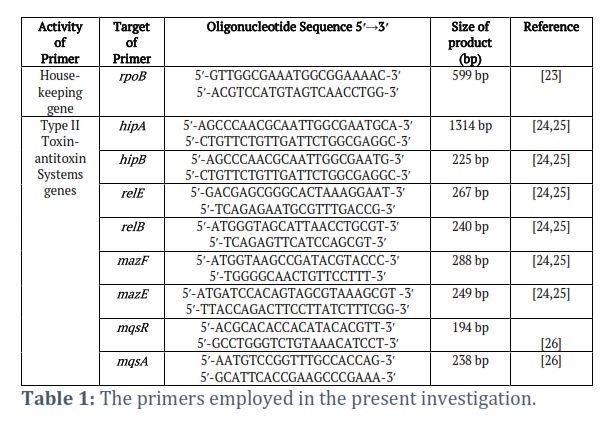
Each gene's initial and secondary primers consist of forward and reverse primers, separately, which play a crucial role in the process of PCR and qRT-PCR. rpoB House-keeping gene, hipA Toxin gene, hipB Antitoxin gene, relE Toxin gene, relB Antitoxin gene, mazF Toxin gene, mazE Antitoxin gene, mqsR Toxin gene, mqsA Antitoxin gene.
Type II TA system gene expression upon various temperatures and pH values
Total RNA was extracted from Overnight cultures of K. pneumoniae isolates at various temperatures (37℃ ± 7℃) and pH values (5, 7, 9, 11) by using MagPurix® Total RNA Extraction Kit with the MagPurix® instrument (Zinexts/ Taiwan, ROC). Then, the purity was assessed using a NanoDrop spectrophotometer (Thermo Fisher Scientific/USA). The synthesis of cDNA from total RNA was achieved through reverse transcription employing the WizScript RT-FDmix Kit (Wizbiosolution/ South Korea). Utilization of the Real-Time PCR system LM 2012 (Fosun Diagnostics/ China) facilitated the execution of this reaction.
Quantitative Reverse Transcriptase PCR assay
The assessment of type II TA systems (including relE toxin, mqsR toxin, and hipB anti-toxin) in terms of their gene expression levels through the utilization of the quantitative Reverse Transcriptase Polymerase Chain Reaction (qRT-PCR) assay was conducted employing the WizPure™ qPCR Master (SYBR) kit (Wizbiosolution/ South Korea). The experimental procedure consisted of a series of steps including an initial cycle at 95℃ lasting 5 min, followed by 40 amplification cycles at 95℃ for (10s) each, and a final cycle at 60℃ for (40s). To ensure accuracy and reliability, the housekeeping gene rpoB was employed as an internal control to standardize the mRNA levels of the genes associated with the type II TA systems (hipB, mqsR, relE). Subsequently, the mRNA expression levels were quantified utilizing the 2^∆∆CT method, enabling the calculation of fold change in expression levels for each gene of interest [27].
Statistical analysis
The results of this investigation were evaluated using Excel Professional Plus 2021 (Microsoft Software, Inc.) and Prism 8.3 (GraphPad Software, Inc.) for each group of three independent biological replicates. The statistical analysis included the comparison of numerical data via one-way ANOVA, with all statistical assessments considered significant at a level of p≤0.05.
Ethics Committee approval
All experiments involving human samples and bacteria collection were carried out under the Helsinki Declaration's standards. Approval was granted by the Ethics Committee within the Biology department at the College of Science, Mustansiryiah University, under the approval reference number BCSMU/1221/0002M.
Results![]()
Clinical Sample Characteristics
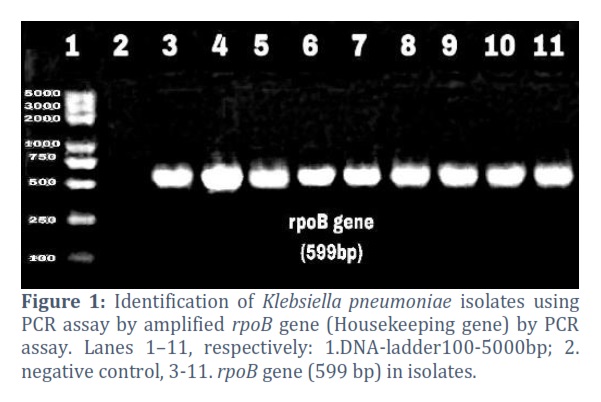
Klebsiella pneumoniae isolates that were sixty-five, were obtained from different hospitals in Baghdad in August -October of 2022. Identification of the isolates was done at the species level using VITEK®2 compact (BioMérieux/ France). The highest number of isolates was obtained from urine samples accounting for 30 isolates (46. 1%), followed by blood samples 11 (16. 9%), ward swabs 10 (15. 3%), sputum samples 8 (12. 3%), burn swabs 4 (6. 1%) and ear swabs 2 (3%). The identity of the K. pneumoniae isolates was confirmed using PCR amplification of the housekeeping gene rpoB (Fig. 1).
Type II TA system identification
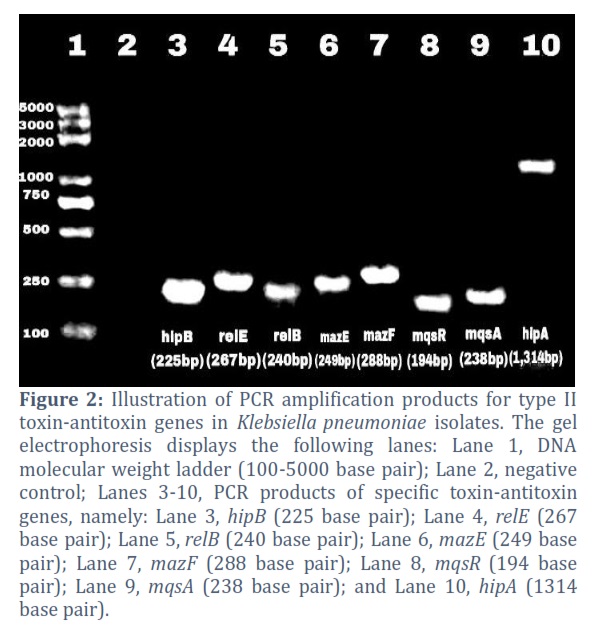
The existence of eight genes belonging to the type II TA systems in K. pneumoniae isolates was verified through PCR amplification. The type II TA system genes prevalence in twenty K. pneumoniae isolates were as follows: 100% for hipB, 10% for hipA, 70% for relE, 35% for relB, 30% for mqsR, 20% for mqsA, and 10% for both mazE and mazF (Fig. 2).
The impact of pH levels and temperatures on Klebsiella pneumoniae isolates growth
Study the influence of (30℃, 37℃, and 44℃) temperatures on the growth of K. pneumoniae at different interval times (0, 8 h, 24 h). The current results show the cell density increased significantly at 37℃, with OD of 1 at 24 h. Cell densities of K. pneumoniae revealed that the cells cultivated at 44℃ exhibited an average OD of 0.6 after 24 h, whereas those grown at 30℃ showed an average OD of 0.7 at the same time point. The findings from the investigation into the impact of various pH values (5, 7, 9, and 11) indicated a significant rise in K. pneumoniae culture cell density after 24 h compared to 8 h, with the most optimal growth observed at pH 7; the tolerance levels at pH 9 and 5 were similar to the results of another study carried [28]. The impact of pH 11 showed no substantial alteration in the K. pneumoniae cell density of from 8 h to 24 h, registering an average OD of 0.1 at 600nm. The growth of K. pneumoniae appeared to be notably slower at pH 11. The outcomes indicate a significant difference (p<0.0001) in the growth curves of bacteria.
The impact of pH levels and temperature on type II TA gene expression
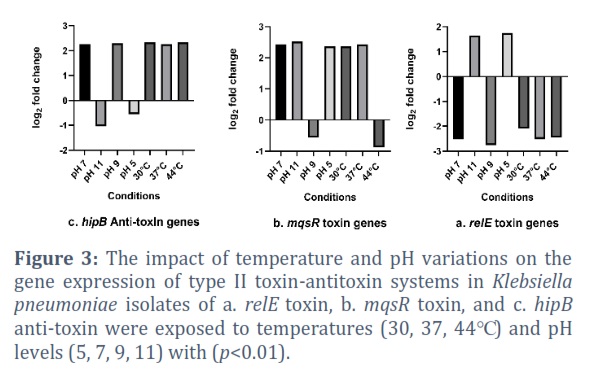
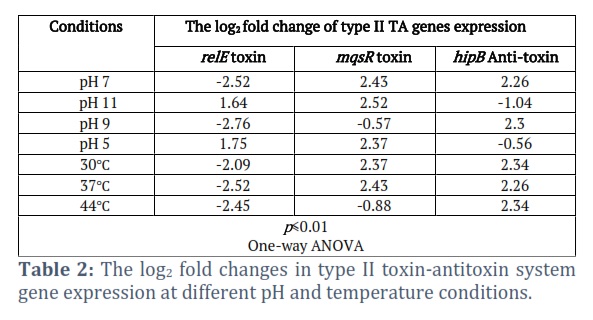
The effect of pH levels and temperatures on the expression levels of type II TA system genes associated with persistence and biofilm formation in K. pneumoniae (including relE toxin, mqsR toxin, and hipB anti-toxin) was assessed through quantitative RT-PCR analysis. The findings from the present investigation indicated that the expression levels of relE toxin, mqsR toxin, and hipB anti-toxin were variably influenced by changes in pH and temperatures (as illustrated in Fig. 3).
Specifically, under acidic conditions at pH 5, K. pneumoniae exhibited a 2.37-fold upregulation in mqsR expression, a 1.75-fold increase in relE expression, and a 0.56-fold decrease in hipB gene expression. Conversely, at alkaline pH 9, there was a 2.3-fold increase in hipB expression, a -0.57-fold reduction in mqsR expression, and a -2.76-fold decrease in relE expression. Moreover, at highly alkaline pH 11, the current results demonstrated a 2.52-fold increase in mqsR expression, a 1.64-fold increase in relE expression, and a -1.04-fold decrease in hipB expression. Similarly, when exposed to pH 7 and a temperature of 37°C, the outcomes revealed a 2.43-fold increase in mqsR expression, a 2.26-fold increase in hipB expression, and a -2.52-fold decrease in relE expression. Furthermore, investigating the impact of various temperatures (30℃ and 44℃) on the gene expression levels of type II TA systems, it was observed that at 30℃, there was a 2.37-fold increase in mqsR expression, a 2.34-fold increase in hipB expression, and a -2.09-fold decrease in relE expression. In contrast, at 44℃, there was a -0.88-fold decrease in mqsR expression, a -2.45-fold decrease in relE expression, and a 2.34-fold increase in hipB expression, all with a significance level of p<0.01 (as presented in Table 2).
Figures & Tables
The findings indicated that there was a significant variation in all three temperatures (30℃, 37℃, and 44℃) observed, with a notable cell density increase in K. pneumoniae isolates at 24 h compared to that at 8 h. Among the temperatures tested, the most favorable for growth was 37℃, as the isolates exhibited rapid replication under these conditions and showed a capacity to endure 44℃, which suggests their survival capability in both temperate and tropical environments. It was noted that K. pneumoniae tends to grow at a slower rate at 30℃ compared to 37℃. These results aligned with a previous study that also reported optimal growth at 37℃ [28]. Elevated temperatures can lead to inactivation processes primarily through protein degradation and destruction, while lower temperatures can extend the lifespan of microorganisms [29]. The stimulation of RpoH in response to elevated temperatures triggers a cascade of events in K. pneumoniae. This stimulation serves as a key mechanism in regulating the production of heat shock proteins (Hsps) which are vital for preventing aggregation, facilitating protein folding, and restoring damaged proteins under extreme heat conditions [30].
In the natural setting, a mere one-unit change in pH can result in up to a 50% reduction in the metabolic rate of microbial populations [18]. The pH level also plays a crucial role in influencing microbial growth, survival, structural integrity, functional aspects, as well as the activities of various components, and extracellular enzymes can be adversely impacted by alterations in the kinetics of redox reactions and thermodynamics [18,31].
Numerous environmental stressors including temperature fluctuations, pH variations, nutrient scarcities, exposure to antibiotics, and high cell densities can trigger the stringent response. This response inhibits critical cellular functions and elevates persister levels within a bacterial population by generating (p)ppGpp and activating TA systems [32,33]. Most type II TA systems implicated in environmental and nutritional stress responses can impede translation through diverse mechanisms. For instance, RelE acts as an endoribonuclease that disrupts translation by causing mRNA cleavage at the ribosomal A-site [34].
The current investigation revealed heightened expression levels of antitoxin hipB genes at various conditions (37℃, 30℃, 44℃, pH 7, pH 9), mqsR toxin gene at specific conditions (37℃, 30℃, pH 7, pH 5, pH 11), and relE toxin genes under certain pH conditions (pH 5, pH 11). These findings suggest a potential link between pH values and temperature with type II TA system gene expression that is associated with antibiotic tolerance, persistence, and biofilm formation, which could potentially enhance bacterial virulence and bolster bacterial survival under adverse conditions.
Recent research has highlighted that the mqsRA TA system displays reduced mqsA antitoxin expression, elevated mqsR gene expression, and a considerable increase in the RelE1 gene under stress conditions [26,35]. Another study showcased an upregulation of the TA system yafQ/dinJ gene expression at 55℃ and an increase in cell survival due to the ParDEI system, a member of the RelE/ParE superfamily, at 42℃. Furthermore, heightened gene expression levels of type II TA system genes (brnTA, relE/RHH-like, and fic/phd) were observed under acid conditions (pH 5.5) in Brucella spp. strains, underscoring the importance of these TA systems in acidic environments [34,36,37].
Bacteria can adapt to severe environments by changing the expression of various genes. The regulation of type II TA systems genes is influenced by various factors, such as pH and temperature. The current research has brought attention to the broad-reaching effects of temperature and pH stress on type II TA systems. The comprehension of how these factors impact the gene expression of type II TA systems, which play a role in antibiotic resistance and biofilm-formation, contributes to our understanding of bacterial behavior as it can help to identify novel mechanisms for controlling bacterial growth, and survival and devise new strategies for managing bacterial infections. The potential applications of manipulating the regulation of these systems are broad and could have significant implications for biotechnology and medicine.
Consent to Participate
Consent was acquired from the parents or legally authorized representatives of all study participants.
Acknowledgment
We extend our appreciation to the College of Science at Mustansiriyah University in Baghdad, Iraq (https://uomustansiriyah.edu.iq), for their provision of resources and assistance in facilitating this research.
Author Contributions
Fatima J. Hassan did all the lab work, collected the data, conducted data analysis, and wrote the final version of the article while Mohammed F. Al-Marjani and Intesar N. Khelkal supervised all research steps. Mohammed F. Al-Marjani, Intesar N. Khelkal, and Fatima J. Hassan participated equally in the research design.
The author(s) declare that there is no conflict of interest regarding the publication of this paper.
![]() References
References
- Riedel S, Hobden JA, Miller S, Morse SA, Mietzner TA, et al. Enteric Gram-Negative Rods (Enterobacteriaceae): Jawetz, Melnick, & Adelberg's Medical Microbiology 28 E. (2019); (n.p.): pp. 243. McGraw-Hill LLC.
- Ahmed HJ, Ganjo AR. Detection of Carbapenemase-Producing Klebsiella pneumoniae and Escherichia coli Recovered from Clinical Specimens in Erbil City Kurdistan Region of Iraq. Al-Mustansiriyah Journal of Science, (2019); 30(2): 10-18.
- AdulZahraa A, Al-Joofy IK, Khelkal IN. Evaluation of Immunomodulatory Activities of Purified Microcin Produced by Klebsiella Pneumoniae in Experimental Animals. Journal of Research on the Lepidoptera, (2019); 50(1): 32-39.
- Aslam B, Siddique MH, Siddique AB, Shafique M, Muzammil S, et al. Distribution of mcr-1 Harboring Hypervirulent Klebsiella pneumoniae in Clinical Specimens and Lytic Activity of Bacteriophage KpnM Against Isolates. Infection and Drug Resistance, (2022); 15: 5795-5811.
- Alansary IMM, Al-Saryi NA. Emergence of hypervirulent Klebsiella pneumoniae isolates from some Iraqi hospitals. Reviews and Research in Medical Microbiology, (2024); 35(2): 88-96.
- Nirwati H, Sinanjung K, Fahrunissa F, Wijaya F, Napitupulu S, et al. Biofilm formation and antibiotic resistance of Klebsiella pneumoniae isolated from clinical samples in a tertiary care hospital, Klaten, Indonesia. BMC Proceedings, (2019); 13(suppl.11): 20.
- Al-Hasso MZ, Gergees SG, Mohialdeen ZK. Molecular characterization of ESBLs and AmpC β-lactamases in Bacteria Isolated from Currency Notes Circulating in Mosul City, Iraq. Karbala International Journal of Modern Science, (2022); 8(3): 543-553.
- Jurėnas D, Fraikin N, Goormaghtigh F, Van Melderen L. Biology and evolution of bacterial toxin–antitoxin systems. Nature Reviews Microbiology, (2022); 20(6): 335-350.
- Al-Marjani MF, Authman SH, Ali FS. Toxin–antitoxin systems and biofilm formation in bacteria. Reviews and Research in Medical Microbiology, (2020); 31(2): 61-69.
- Singh G, Yadav M, Ghosh C, Rathore JS. Bacterial toxin-antitoxin modules: classification, functions, and association with persistence. Current Research in Microbial Sciences, (2021); 2: 100047.
- Fraikin N, Goormaghtigh F, Van Melderen L. Type II toxin-antitoxin systems: evolution and revolutions. Journal of bacteriology, (2020); 202(7); 00763-00719.
- Song S, Wood TK. A primary physiological role of toxin/antitoxin systems is phage inhibition. Frontiers in Microbiology, (2020); 11: 1895.
- Bleriot I, Blasco L, Pacios O, Fernández-García L, Ambroa A, et al. The role of PemIK (PemK/PemI) type II TA system from Klebsiella pneumoniae clinical strains in lytic phage infection. Scientific reports, (2022); 12(1): 4488.
- Yang QE, Walsh TR. Toxin–antitoxin systems and their role in disseminating and maintaining antimicrobial resistance. FEMS microbiology reviews, (2017); 41(3): 343-353.
- Dai Z, Wu T, Xu S, Zhou L, Tang W, et al. Characterization of toxin-antitoxin systems from public sequencing data: A case study in Pseudomonas aeruginosa. Frontiers in Microbiology, (2022); 13: 951774.
- Gerdes K, Bærentsen R, Brodersen DE. Phylogeny reveals novel HipA-homologous kinase families and toxin-antitoxin gene organizations. MBio, (2021); 12(3); 01058-01021.
- Roy PK, Ha AJ-W, Mizan MFR, Hossain MI, Ashrafudoulla M, et al. Effects of environmental conditions (temperature, pH, and glucose) on biofilm formation of Salmonella enterica serotype Kentucky and virulence gene expression. Poultry Science, (2021); 100(7): 101209.
- Jin Q, Kirk MF. pH as a primary control in environmental microbiology: 2. Kinetic perspective. Frontiers in Environmental Science, (2018); 6: 101.
- Zou J, Peng B, Qu J, Zheng J. Are bacterial persisters dormant cells only? Frontiers in Microbiology, (2022); 12: 708580.
- Atlas RM. Chapter L: Handbook of microbiological media. (2004); pp. 62. CRC press.
- Germain-Amiot N, Augagneur Y, Camberlein E, Nicolas I, Lecureur V, et al. A novel Staphylococcus aureus cis-trans type I toxin-antitoxin module with dual effects on bacteria and host cells. Nucleic Acids Research, (2019); 47(4): 1759-1773.
- Ruppé E, Hem S, Lath S, Gautier V, Ariey F, et al. CTX-M β-lactamases in Escherichia coli from community-acquired urinary tract infections, Cambodia. Emerging infectious diseases, (2009); 15(5): 741.
- Alansary IMM, Al-Saryi NA. Detection of biofilm formation in classical and hypervirulent Klebsiella pneumoniae. Al-Mustansiriyah Journal of Science, (2022); 33(5): 65-71.
- Hemati S, Azizi-Jalilian F, Pakzad I, Taherikalani M, Maleki A, et al. The correlation between the presence of quorum sensing, toxin-antitoxin system genes, and MIC values with ability of biofilm formation in clinical isolates of Pseudomonas aeruginosa. Iranian journal of microbiology, (2014); 6(3): 133.
- Karimi S, Ghafourian S, Kalani MT, Jalilian FA, Hemati S, et al. Association between toxin-antitoxin systems and biofilm formation. Jundishapur Journal of Microbiology, (2015); 8(1): e14540.
- Al-Husseini LB, Maleki A, Al-Marjani MF. Antisense mqsR-PNA as a putative target to the eradication of Pseudomonas aeruginosa persisters. New Microbes and New Infections, (2021); 41: 100868.
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods, (2001); 25(4): 402-408.
- Tsuji A, Kaneko Y, Takahashi K, Ogawa M, Goto S. The effects of temperature and pH on the growth of eight enteric and nine glucose non‐fermenting species of gram‐negative rods. Microbiology and immunology, (1982); 26(1): 15-24.
- Pepper IL, Gerba CP, Gentry TJ . Introduction to environmental microbiology: Environmental microbiology. (2015); pp. 3-8. Academic Press.
- Jones JA, Andreas MP, Giessen TW. Structural basis for peroxidase encapsulation inside the encapsulin from the Gram-negative pathogen Klebsiella pneumoniae. Nature Communications, (2024); 15(1): 2558.
- Lund PA, De Biase D, Liran O, Scheler O, Mira NP, et al. Understanding how microorganisms respond to acid pH is central to their control and successful exploitation. Frontiers in Microbiology, (2020); 11: 556140.
- Hauryliuk V, Atkinson GC, Murakami KS, Tenson T, Gerdes K. Recent functional insights into the role of (p) ppGpp in bacterial physiology. Nature Reviews Microbiology, (2015); 13(5): 298-309.
- Eisenreich W, Rudel T, Heesemann J, Goebel W. Persistence of intracellular bacterial pathogens—With a focus on the metabolic perspective. Frontiers in Cellular and Infection Microbiology, (2021); 10: 615450.
- Kamruzzaman M, Iredell J. A ParDE-family toxin-antitoxin system in major resistance plasmids of Enterobacteriaceae confers antibiotic and heat tolerance. Scientific Reports, (2019); 9(1): 9872.
- Narimisa N, Amraei F, Kalani BS, Azarnezhad A, Jazi FM. Biofilm establishment, biofilm persister cell formation, and relative gene expression analysis of type II toxin-antitoxin system in Klebsiella pneumoniae. Gene Reports, (2020); 21: 100846.
- Kamruzzaman M, Wu AY, Iredell JR. Biological functions of type II toxin-antitoxin systems in bacteria. Microorganisms, (2021); 9(6): 1276.
- Amraei F, Narimisa N, Mohammadzadeh R, Lohrasbi V. The expression of type II TA system genes following exposure to the sub-inhibitory concentration of gentamicin and acid stress in Brucella spp. Microbial Pathogenesis, (2020); 144: 104194-104194.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()