

Full Length Research Article
Characterization of garlic virus A, garlic virus D, and onion yellow dwarf virus infecting onion
Mahmoud A. Amer*, Muhammad Amir, Khadim Hussain, Ibrahim M Al-Shahwan, Mohammed A. Al-Saleh
Adv. life sci., vol. 11, no. 4, pp. 791-796, November 2024
*– Corresponding Author: Mahmoud A. Amer (ruamerm@ksu.edu.sa)
Authors' Affiliations
[Date Received: 30/08/2023; Date Revised: 19/07/2024; Date Published: 15/10/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Allium cepa is one of the major vegetable crops cultivated in Saudi Arabia. This economically important crop faces several biotic stresses which adversely affect the quality and quantity of its production. Several virus groups (potyviruses, carlaviruses, tospoviruses, and allexiviruses) have been reported infecting Allium crops.
Methods: During the growing season of 2021-2022, a total of 81 onion samples exhibiting virus-like symptoms were collected from two different geographical regions in Saudi Arabia. The serological technique (ELISA) was used to detect the important allexiviruses and potyviruses. RT-PCR amplification of partial genome sequence was done using degenerate primers for allexiviruses and potyviruses and the phylogenetic trees were constructed using different bioinformatic tools.
Results: The results obtained from ELISA tests showed that 26% and 32% of onion samples were positive with both Garlic Virus A (GarV-A) and Onion yellow dwarf virus (OYDV) respectively. RT-PCR amplification and sequencing results showed that two allexiviruses, GarV-A, garlic virus D (GarV-D), and one Potyvirus (OYDV) were detected in both regions. Sequence data were deposited in the GenBank database with accession numbers, OQ397545, OQ397546 for GarV-A, OQ397547 for GarV-D, and OQ397548, OQ397549 for OYDV, sequentially. Phylogenetic tree analysis of these virus isolates showed making clades with closely related isolates of their respective viruses. Pairwise nucleotide sequence identity showed their similarity with GarV-A, GarV-D, and OYDV isolates reported earlier in the GenBank.
Conclusion: To the best of our knowledge, these two distinct allexiviruses (GarV-A, GarV-D) and one Potyvirus (OYDV) were isolated for the first time from an onion crop in Saudi Arabia.
Keywords: Allexiviruses; Potyviruses; Serological Detection; Allium cepa; RT-PCR; Sequence
Introduction![]()
Among monocotyledonous plants, Allium is one of the largest genera which comprises almost 950 species [1]. Onion (Allium cepa L.) is one of the economically important crops which is grown worldwide [2]. Onion cultivation makes up around 24% of all vegetable production in the world (FAOSTAT, 2018). The top ten nations that cultivate onions are China, India, the United States, Egypt, Iran, Turkey, Pakistan, Brazil, the Russian Federation, and the Republic of Korea [3]. In Saudi Arabia, onion crops are cultivated over a 7650 ha area and its production reached up to 297974 tons according to APSB, 2019 [4].
Viruses, bacteria, fungi, and the cost of cultivation all have an impact on onion production. Unfortunately, several viral infections have an adverse impact on the quantity and quality of crops annually. In cases of severe infections, the losses increase up to 25–50% of the production. When multiple virus infections occur frequently in the onion crop, it is referred to as a “garlic viral complex" and plants show a variety of symptoms such as yellow stripes, stunted growth, leaf deformation, mosaics on leaves, and leaf striping [2, 5].
The Allium plants are affected by more than 20 viruses belonging to the four genera: Potyvirus, Carlavirus, Allexivirus, and Tospovirus that have been reported to lessen the yields and quality of garlic and onion crops significantly [5-12].
The viruses belonging to the genus Potyvirus and Carlavirus are transmitted by aphids in a non-persistent manner [13] whereas a Tospovirus is efficiently transmitted by thrips at the larval stage [14]. While eriophyid mites are responsible for the transmission of Allexivirus. All these four virus groups are also transmitted through mechanical inoculation [15, 16]. The host range for these four groups is limited to some indicator plants such as onions, garlic, shallots, a few ornamental Alliums, Chenopodium amaranticolor, C. quinoa, Datura stramonium and Vigna unguiculata [16, 17]. In Saudi Arabia, viruses exhibiting symptoms such as light and dark green patches, yellowing, leaf striping, stunted growth, and leaf deformation were observed in onion fields in two different geographical regions of Riyadh and Al-Baha. The aim of the present study is to identify the Allexivirus and Potyvirus infecting onion plants in Saudi Arabia using serological and molecular techniques.
Methods![]()
Sample collection and ELISA
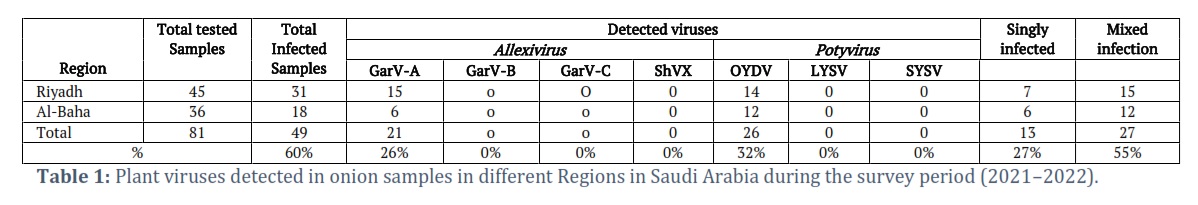
Onion plants that exhibited the symptoms of yellowing, dwarfism, yellow mosaic, and twisted leaves were observed in Riyadh and Al-Baha regions during field surveys in the years 2021-2022 (Figure 1). A total of 81 samples of onion plants were collected from the Al-Baha and Riyadh Regions (Table 1). Double antibody sandwich enzyme-linked immunosorbent assay (DAS-ELISA) with specific antibodies obtained from LOEWE®, Biochemica, and DSMZ, Germany, were used to test the collected samples against the important allexiviruses including Garlic virus A (GarV-A), Garlic virus B (GarV-B), Garlic virus C (GarV-C), shallot virus X (ShVX), and potyviruses including Onion yellow dwarf virus (OYDV), Leek yellow stripe virus (LYSV) and Shallot yellow stripe virus (SYSV).
The procedure was performed and demonstrated according to the manufacturer’s protocol using BioTek ELx808 ELx808IU Absorbance Microplate Reader96-Well. Samples were considered positive according to Clark and Adams,1977 [18] and Daryono et al., 2003 [19].
Reverse transcription polymerase chain reaction (RT-PCR)
Total RNA was extracted from all collected leave samples (100mg) using Gene JET Plant RNA Purification Mini Kit and One-step RT-PCR was carried out by following the instruction of the manufacturer’s protocol of Thermo Scientific, USA using degenerate primer for allexiviruses (Allex-CP (+) and Allex-NABP (−) [20]. RT-PCR cycle was performed as follows: 60℃ for 10 min and 98℃ for 2 min followed by 35 cycles, 94℃ for 45 sec, 60℃ for 45 sec, 72℃ for 1 min, 72℃ for 10 min as final extension. For Potyvirus, the degenerate primers (Nib2-F and Nib3-R) were used [21] and the RT-PCR cycles were set to 42°C for 45 minutes, 95°C for 12 seconds, 40 cycles, 95°C for 15 seconds, 48°C for 20 seconds, 72°C for 30 seconds, and 72°C for 1 minute as the final extension.
The RT-PCR products were electrophoresed using 1% agarose gel in TBE buffer (Tris Borate EDTA) [22]. A Gel Documentation system was used to analyze the PCR-amplified products. The expected molecular sizes have been determined using the 50 bp HyperLadder DNA (Bioline Ltd., USA) and the 100 bp DNA Ladder (GeneRuler, Thermo Scientific, USA).
Analysis of phylogenetic trees with nucleotide sequencing
Three and two RT-PCR products were obtained from onion samples by general primers of Allexivirus and Potyvirus groups, respectively. These amplified products were purified and sent to Macrogen Inc. Seoul, South Korea for two-directional Sanger sequencing of the DNA. Sequencing data was aligned using DNA star computer software and the National Centre for Biotechnology Information (NCBI). Using 1000 bootstrap replications and the Maximum-Likelihood method algorithm, the phylogenetic trees were constructed from ClustalW-aligned sequences by MEGA-X to analyze the phylogenetic relationship of the Saudi isolates with the other isolates reported from different countries. Pairwise nucleotide sequence identity percentages were determined using the MegAlign tool in the DNAStar software package by Lasergene and a graphical explanation of pairwise nucleotide percentage identity was presented by Sequence Demarcation Tool (SDT) [23].
Results![]()
Field Observation and ELISA
Eighty-one symptomatic samples exhibited mosaic, dwarfism, yellowing, leaf striping, stunted growth, and deformation of leaves were collected from two main different onion growing regions (Al-Baha and Riyadh) in Saudi Arabia (Figure 1). The obtained results of ELISA from 81 onion plant samples indicated the occurrence of GarV-A and OYDV with a detection percentage of 26% and 32%, respectively. The obtained ELISA results showed that 47/81 (58%) onion samples were positive for at least one of the tested viruses. As far as a single and mixed infection of GarV-A and OYDV was concerned, 27% of the infected samples were singly infected with either GarV-A or OYDV and 55% of those had mixed infection of both viruses. The GarV-B, GarV-C, and ShVX were not detected in any of the tested samples (Table 1).
RT-PCR Reaction
For the detection of allexiviruses from onion samples using general primers, the amplified RT-PCR products have been of expected size 750 bp (Figure 2 A). In the case of Potyvirus amplification, the expected size of the amplicon was 350 bp (Figure 2 B). RT-PCR amplification was not observed with total RNA isolated from healthy plants as a negative control with any of the primer pairs used in this study. The outcome of these results revealed that ELISA-positive onion samples were infected with at least one of the viruses belonging to Allexivirus and Potyvirus. The presence of the exact virus infection was confirmed after Sanger DNA sequencing of these RT-PCR amplified samples.
Phylogenetic tree analysis with partial nucleotide sequencing
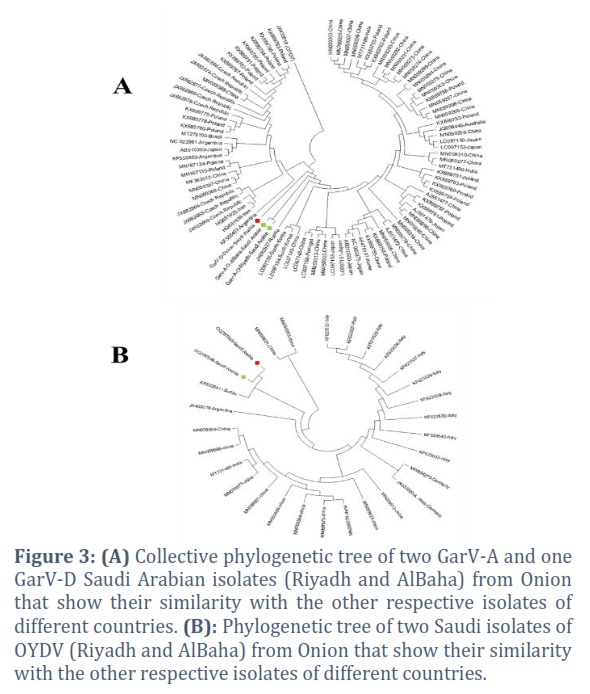
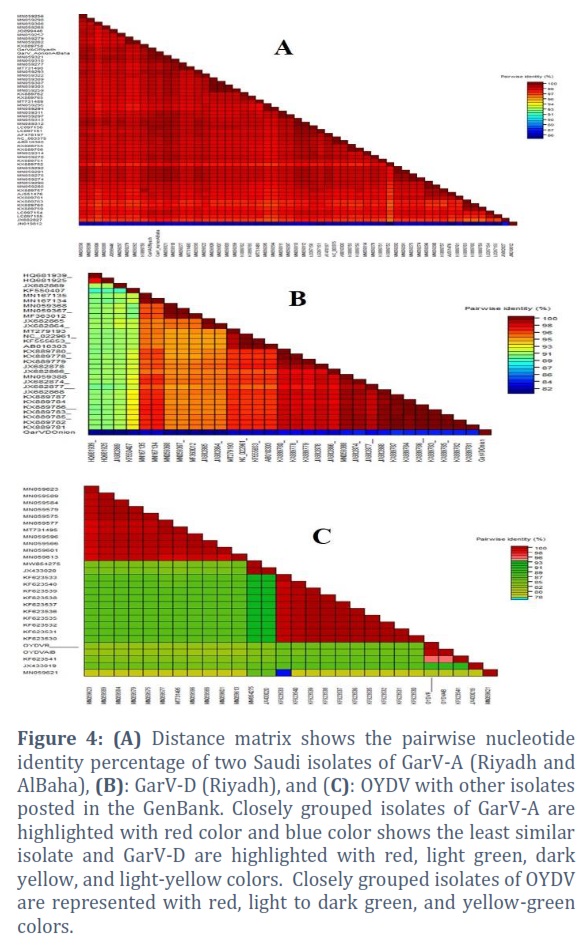
For general allexiviruses, two isolates of GarV-A and one isolate of GarV-D isolated from the onion samples showed a higher level of sequence identity with GarV-A and GarV-D based on their BLAST (basic local alignment search tool) analysis. After completing the analysis, sequences have been submitted to GenBank with the following accession numbers: OQ397545 (GarVA-O-Riyadh), OQ397546 (GarVA-O-AlBaha), and OQ397547 (GarVD-O-Riyadh) sequentially. The phylogenetic tree of GarV-A showed that our Saudi isolates grouped with the GarV-A isolates from Russia which were isolated from Allium sativum (JX682827), and China (LC097148) and our GarV-D isolate made a clade with GarV-D isolates from Argentine isolated from A. sativum (KF550407; Figure 3 A). As far as their nucleotide sequence identity percentage is concerned, GarV-A isolates from Riyadh and Al-Baha regions had 100% identity among themselves and 96.7% to 98.5% with the already published isolates in NCBI GenBank databases. The highest percentage identity was observed with GarV-A Chinese isolate (MN059312, MN059313), and the lowest percentage was observed with Poland isolate (KX889752; Figure 4 A). Moreover, GarV-D shared 80.6% to 83.6% of identity with the other available isolates in the NCBI GenBank. The highest percentage was observed with Poland isolate (KX889787), and the lowest percentage was also observed with Poland isolate (HQ681925; Figure 4 B).
For Potyvirus, the nucleotide sequence analysis of the two isolates of Potyvirus isolated from the onion samples in Riyadh and Al-Baha revealed that these sequences were related to OYDV. These sequences were submitted to GenBank with the accession numbers OQ397548 (OYDV-Riyadh) and OQ397549 (OYDV-AlBaha), respectively. The phylogenetic tree and SDT analysis of these two isolates of OYDV were constructed (Figures 3 B and Figure 4 C). Both isolates showed 99% to 100% identity between themselves. However, OYDV from Riyadh shared 80% to 95% identity with other isolates from NCBI GenBank, and OYDV from Al-Baha showed 80% to 95.5% similarity with the NCBI GenBank isolates.
Figures & Tables


Onion is a major bulbous crop among vegetables and is of global importance [24, 25]. As a result of vegetative propagation, which causes viruses to accumulate and replicate in planting material, viruses are the primary issue in bulb crops like onion and garlic. Although viruses that infect Allium species have been intensively studied. Due to the multiple virus infections, serological and molecular studies were not easy to conduct for the detection of these viruses [26-28]. Viruses are ubiquitous worldwide and cause impacts in the form of crop production loss and reduction in quality [29, 30, 7]. It is tough to identify and estimate yield losses that are precisely attributable to each virus due to the incidence of mixed virus infections.
In eighty-one samples, 47 samples of onion (58%) were positive with GarV-A and OYDV. Hence, a large number of infected samples showed that the exchange of infected planting material from one region to another region could be the main reason for their occurrence in surveyed regions, but the presence of their vectors (aphids and mites) cannot be neglected because they play a significant role in their short distance disseminations within the fields.
Furthermore, in this study, two allexiviruses (GarV-A and GarV-D) and one Potyvirus (OYDV) isolated from onion were amplified by degenerate primers in the Riyadh and Al-Baha regions of Saudi Arabia. Interestingly, GarV-D could not be detected during the ELISA assay (unavailability of antisera) but got amplified in RT-PCR using general allexiviruses primers. This showed along with ELISA tests, a variety of other molecular techniques, RT-PCR, and hybridization must be incorporated into detection methods. Thus, a virus's entire genome can be identified by molecular analysis, and only a small number of viruses can be identified through serological investigation [31-36]. Therefore, molecular analysis is always chosen over serological testing due to their higher sensitivity and robustness.
In RT-PCR, two Saudi Arabian isolates of GarV-A from Riyadh and Al-Baha regions and one Saudi Arabian isolate of GarV-D from Riyadh region showed their nucleotide identities of 96.7% to 98.5% and 80.6% to 83.6%, respectively with already published isolates in GenBank from different countries. Moreover, two Saudi Arabian isolates of OYDV from Riyadh and Al-Baha regions showed 100% nucleotide sequence identity with each other and 80.4% to 95.5% nucleotide identity with the other isolates of different countries. So, natural genetic variation could be the reason for the variation in the nucleotide identity of these isolates. Generally, phylogenetic analyses unveiled that Saudi Arabian onion isolates are normally grouped together, and the phylogenetic similarity relationship between them and their geographical roots is frequently unrelated. The great diversity and swift evolution of these viruses have been associated with the international exchange of vegetative planting materials between states, based on numerous studies conducted in other countries [37-40].
The results of this investigation would be useful in providing information about the evolutionary changes and diversity of these significant viruses. Also, this research may play a significant role in enhancing disease management procedures. Therefore, biological research is necessary to identify the transmission routes and assess the effects of isolated and combined infections of different isolates and different viruses causing mixed infections on onion crops. According to the best of our information, this is the first report of GarV-A, GarV-D, and OYDV infecting onion crops in Saudi Arabia, and this is also the first report of GarV-A from onion in the world.
Acknowledgements
The authors extend their appreciation to the Deputyship for Research and Innovation, Ministry of Education in Saudi Arabia for funding this research work through project no. (IFKSUOR3-556-2).
Author Contributions
Mohammed Amir, Mohammed Al-Saleh, and Mahmoud Amer designed the study, prepared the materials, collected the data, conducted the analysis, wrote and edited the manuscript. Ibrahim M. Al-Shahwan and Khadim Hussain conducted the data analysis.
The author declare that there is no conflict of interest regarding the publication of this paper.
![]() References
References
- Friesen N, Fritsch RM, Blattner FR. Phylogeny and new intrageneric classification of Allium (Alliaceae) based on nuclear ribosomal DNA ITS sequences. Aliso: A Journal of Systematic and Floristic Botany, (2006); 22 (1): 372-395.
- Abraham AD, Kidanemariam DB, Holton, TA. Molecular identification, incidence, and phylogenetic analysis of seven viruses infecting garlic in Ethiopia. European Journal of Plant Pathology, (2019); 155: 181-191.
- Dossa FK, Miassi YE, Banzou K. Onion (Allium cepa) production in urban and peri-urban areas: financial performance and importance of this activity for market gardeners in Southern Benin. Current Investigations in Agriculture and Current Research, (2018); 3(2): 334-346.
- Agricultural Production Survey Bulletin (2019), https://www.stats.gov.sa/sites/default/files/Agriculture%20Production%20Survey%202019%20EN.pdf, (Accessed: 9.9.2024).
- Van Dijk P, Van der Vlugt RA. New mite-borne virus isolates from rakkyo, shallot and wild leek species. European Journal of Plant Pathology, (1994); 100: 269-277.
- Maliogka VI, Dovas CI, Lesemann DE, Winter S, Katis NI. Molecular identification, reverse transcription-polymerase chain reaction detection, host reactions, and specific cytopathology of Artichoke yellow ringspot virus infecting onion crops. Phytopathology, (2006); 96(6): 622-629
- Cafrune EE, Perotto MC, Conci VC. Effect of two Allexivirus isolates on garlic yield. Plant Disease, (2006); 90(7): 898-904.
- Miller ME, Schwartz, HF. Stemphylium leaf blight and stalk rot: Compendium of Onion Garlic Diseases and Pests. 2008; 45-47. American Phytopathological Society
- Shahraeen N, Lesemann DE, Ghotbi T. Survey for viruses infecting onion, garlic and leek crops in Iran. EPPO Bulletin, (2008); 38(1): 131-135.
- Perotto MC, Cafrune EE, Conci VC. The effect of additional viral infections on garlic plants initially infected with Allexiviruses. European Journal of Plant Pathology, (2010); 126(4): 489-495.
- King AM, Adams MJ, Carstens EB, Lefkowitz EJJ. Virus taxonomy:Ninth report of the International Committee on Taxonomy of Viruses. 2012; 1327. Elsevier
- Dąbrowska E, Lewandowski M, Koczkodaj S, Paduch-Cichal E. Transmission of Garlic virus B, Garlic virus C, Garlic virus D and Garlic virus X by Aceria tulipae (Keifer) in leek. European Journal of Plant Pathology, (2020); 157: 215-222.
- Karavina C, Ibaba JD, Gubba A, Pappu HR. First report of iris yellow spot virus infecting garlic and leek in Zimbabwe. Plant Disease, (2016); 100(3): 657-657.
- Gera A, Kritzman A, Ganaim N, Raccah B. Transmission of tomato spotted wilt Tospovirus by F. occidentalis and analysis of the distribution of the virus in the field in Israel. In Proceedings of the Sixth International Symposium on Thysanoptera. (1998); 25-29.
- Mahmoud SY, Abo-el-Maaty SA, El-Borollosy AM, Abdel-Ghaffar, MH. Identification of onion yellow dwarf Potyvirus as one of the major viruses infecting garlic in Egypt. American-Eurasian Journal of Agricultural & Environmental Sciences, (2007); 2(6): 746-755.
- Bag S, Pappu, HR. Symptomatology of Iris yellow spot virus in selected indicator hosts. Plant Health Progress, (2009); 10(1): 41.
- Mansouri F, Krahulec F, Duchoslav M, Ryšánek P. Newly identified host range of viruses infecting species of the genus Allium and their distribution in six habitats in the Czech Republic. Plant Pathology, (2021); 70(6): 1496-1507.
- Clark MF, Adams A. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. Journal of General Virology, (1977); 34(3): 475-483.
- Daryono BS, Somowiyarjo S, Natsuaki KT. New source of resistance to Cucumber mosaic virus in melon. SABRAO Journal of Breeding and Genetics, (2003); 35: 19-26.
- Chen J, Zheng HY, Antoniw JF, Adams MJ, Chen JP et.al. Detection and classification of allexiviruses from garlic in China. Archives of Virology, (2004); 149: 435-445.
- Sokhandan-Bashir N, Ghasemzadeh A, Masoudi N, Khakvar R, Farajzadeh D. Degenerate primers facilitate the detection and identification of potyviruses from the northwest region of Iran. Iranian Journal of Biotechnology, (2013); 11(2): 115-122.
- Sambrook J, Russell D. Molecular Cloning: A Laboratory Manual. 2001; Cold Spring Harbor Laboratory Press
- Muhire BM, Varsani A, Martin DP. SDT: a virus classification tool based on pairwise sequence alignment and identity calculation. PloS one, (2014); 9(9): e108277
- Onion a vegetable with global market, https://www.dawn.com/news/275386/onion-a-vegetable-with-global-market, (Accessed: 10.9.2024). .
- Anisuzzaman M, Ashrafuzzaman M, Ismail MR, Uddin MK, Rahim MA. Planting time and mulching effect on onion development and seed production. African Journal of Biotechnology, (2009); 8(3): 412-416.
- Van Dijk P. Virus diseases of Allium species and prospects for their control. International Symposium on Alliums for the Tropics, (1993); 358: 299-306
- Chen J, Chen J, Adams MJ. Molecular characterization of a complex mixture of viruses in garlic with mosaic symptoms in China. Archives of Virology, (2001); 146: 1841-1853.
- Fajardo TV, Nishijima M, Buso JA, Torres AC, Ávila AC et al. Garlic viral complex: identification of Potyviruses and Carlavirus in Central Brazil. Fitopatologia Brasileira, (2001); 26: 619-626.
- Paludan N. Virus attack on leek: Survey, diagnosis, tolerance of varieties and winter hardiness [variety trials, overwintering, Denmark]. Tidsskrift for planteavl, (1980); 84: 371-385.
- Lot H, Chovelon V, Souche S, Delecolle B. Effects of onion yellow dwarf and leek yellow stripe viruses on symptomatology and yield loss of three French garlic cultivars. Plant Disease, (1998); 82(12): 1381-1385.
- Boonham N, Kreuze J, Winter S, van der Vlugt R, Bergervoet J et al. Methods in virus diagnostics: from ELISA to next generation sequencing. Virus Research, (2014); 186: 20-31.
- Hadidi A, Flores R, Candresse T, Barba M. Next-generation sequencing and genome editing in plant virology. Frontiers in Microbiology, (2016); 7: 1325.
- Jones S, Baizan-Edge A, MacFarlane S, Torrance L. Viral diagnostics in plants using next generation sequencing: computational analysis in practice. Frontiers in Plant Sciences, (2017); 8: 1770.
- Singh J, Singh MK, Ranjan K, Kumar A, Kumar P et al. First complete genome sequence of garlic virus X infecting Allium sativum-G282 from India. Genomics, (2020); 112(2): 1861-1865.
- Shahid MS, Sattar MN, Iqbal Z, Raza A, Al-Sadi AM. Next-generation sequencing and the CRISPR-Cas nexus: A molecular plant virology perspective. Frontiers in Microbiology, (2021); 11: 609376.
- Prajapati MR, Manav A, Singh J, Kumar P, Kumar A et al. Identification and Characterization of a Garlic Virus E Genome in Garlic (Allium sativum L.) Using High-Throughput Sequencing from India. Plants, (2022); 11(2): 224.
- Koo BJ, Kang SC, Chang MU. Survey of garlic virus disease and phylogenetic characterization of garlic viruses of the genus Allexivirus isolated in Korea. Plant Pathology Journal, (2002); 18(5): 237-243.
- Wylie SJ, Li H, Jones MG. Phylogenetic analysis of allexiviruses identified on garlic from Australia. Australasian Plant Disease Notes, (2012); 7: 23-27.
- Bereda M, Paduch-Cichal E, Dąbrowska E. Occurrence and phylogenetic analysis of allexiviruses identified on garlic from China, Spain, and Poland commercially available on the polish retail market. European Journal of Plant Pathology, (2017); 149: 227-237.
- Taglienti A, Tiberini A, Manglli A, Rea R, Paoletti S et al . Molecular identification of allexiviruses in a complex mixture of garlic viruses in Latium (central Italy). European Journal of Plant Pathology, (2018); 150: 797-801.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()