

Full Length Research Article
Construction of subunit-based DNA vaccines against tuberculosis using five different Mycobacterium specific genes
Muhammad Mohsin Zaman1, Mirza Imran Shahzad1, Aeman Jilani1, Areeba Yousaf1, Ansah Bashir1, Saher Riaz1, Gildardo Rivera2
Adv. life sci., vol. 11, no. 3, pp. 585-592, August 2024
*– Corresponding Author: Muhammad Mohsin Zaman (mohsinzaman3@gmail.com)
Authors' Affiliations
2. Instituto Politécni co Nacional Centro de Biotecnología Genó mica Mexico City – Mexico
[Date Received: 26/05/2023; Date Revised: 29/03/2024; Date Published: 10/07/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Tuberculosis remains the major infectious and contagious disease with respect to morbidity and mortality around the globe. The main etiological agent of TB is Mycobacterium tuberculosis (M. tb). The cure and/or control of this disease is getting difficult day by day especially due to emergence of drug resistant strains, HIV co-infection and unavailability of good vaccines. BCG is the only available and permitted vaccine against TB, which has variable efficacy only limited to childhood. Therefore, new, well defined, antigen specific and state of the art Molecular Biology based DNA vaccines are required.
Methods: The current study is designed to develop subunit-based DNA vaccines against tuberculosis by using five different Mycobacterium specific genes, namely Rv1908c/KatG, Rv0350/DnaK, Rv0440/GroEL2, Rv0934/PstS and Rv3418c/GroES. All the selected genes were amplified through PCR and cloned into mammalian expression pVAX1 vector.
Results: All the transformed constructs were confirmed through restriction digestion, colony PCR and sequence analysis. These constructs will be used for various in Vivo immunization studies and also for the challenge studies against TB in future experiments.
Conclusion: DNA vaccines, alone or in combination with BCG, have enough potential to be a good therapeutic tool for TB and reduce the treatment time in future.
Keywords: DNA vaccine; pVAX1; Tuberculosis; Vaccines
Introduction![]()
Tuberculosis (TB) is the main infectious and communicable disease with respect to ill health and mortality worldwide, produced by group of bacteria that are closely related to each other and belong to Mycobacterium Tuberculosis Complex (MTBC) [1]. It was on the top regarding cause of deaths by a single infectious agent until the coronavirus (COVID-19) pandemic. According to (WHO Global TB Report 2021), almost 1.7 million people (16%) around the world were killed by TB from 10.4 million infected individuals [2]. Additionally, 161000 MDR (Multi-Drug resistance) cases of Tuberculosis were recorded. In Pakistan, 8.4% people died from 525000 annually diagnosed TB cases, positioning the country as the 5th highest in terms of high-burden TB carrying nations [3].
Production and development of capacity to stimulate protective immunity by injecting inactivated or attenuated pathogen into host, has improved our ability to control or eradicate the different human pathogenic diseases [4]. Tang et al. showed in 1992 that immune responses could be induced by injecting DNA [5]. Latterly, efficacy and advantages of DNA vaccines was reported by producing immunity in various animal models of bacterial, viral and parasitic infections [6].
The only permitted and available vaccine by the WHO for use in human TB cases is Bacille Calmette Guerin (BCG) which is prepared from the attenuation of M. Bovis [7]. BCG is injected in infants at the time of birth as routine immunization and has been effective to provide immunity with reduction in severe form of childhood TB. But its efficacy decreases with time which results in insufficiency to control the disease in adults [8].
The efficacious limitation of BCG vaccine against TB is the motivating factor for the production of modern and better vaccines [9]. These include Recombinant BCG vaccine, plasmid-based DNA vaccines which encode genes of M. tuberculosis, attenuated M. tuberculosis and subunit based recombinant protein antigens [10].
Amongst the latest vaccine platforms, MTBVAC has the potential to offer enhanced protection against tuberculosis (TB) in comparison to traditional BCG vaccines. But MTBVAC is currently undergoing phase II clinical trials to evaluate its safety, immunogenicity, and protective efficacy in humans. These trials aim to assess the vaccine's ability to prevent TB infection and disease progression [11,12]. Furthermore, various plasmid DNA vectors like recombinant plasmid-DNA vectors have been widely used for microbial antigen-coding genes transformation [13]. It is reported that these recombinant vectors (DNA vaccines) could efficiently induce both CD8+ and CD4+ T lymphocyte responses and stimulation of B lymphocytes, which is required for vaccination of TB [14]. Early studies exploring the use of DNA vaccines against TB were released in 1996, utilizing antigens derived from M. leprae, specifically Hsp65 and Ag85A. Subsequently, research expanded, with more than sixty antigens being investigated in various preclinical TB models as DNA vaccines [15]. Generally, it is widely acknowledged that administering Mycobacterium antigens intradermally or intramuscularly in the form of DNA vaccines to mice leads to the induction of robust TH1 immune responses. This immune response is characterized by increased levels of IFNγ and IL-2. Among the DNA vaccines studied, Hsp65, ESAT6, Ag85A, and Ag85B have been particularly well-investigated [16,17]. A vaccine candidate known as M72/AS01E, designed to protect against tuberculosis, is currently undergoing phase II-b clinical trials. While the results of these trials are anticipated to be significant in terms of its defensive efficacy, phase III trials are also expected to provide further insight into its effectiveness. Additionally, another vaccine candidate named TB/FLU-04L, which incorporates Ag85a and ESAT-6 antigens, is currently in phase II-a clinical trials. This vaccine is formulated using an attenuated viral vector of influenza [18]. Further, various clinical trials are being performed to evaluate the efficacy of DNA vaccines. Production of DNA vaccines is a promising area because very few DNA vaccines have been constructed in the past 30 years while many are in the pipeline [19].
This study was designed and conducted with the aim to produce subunit-based DNA vaccines against tuberculosis by using five different Mycobacterium specific genes, namely Rv1908c/KatG, Rv0440/GroEL2, Rv3418c/GroES, Rv0350/DnaK and Rv0934/PstS. All these selected genes encode for the proteins which are involved in the survival, growth and propagation of the organism and can be good candidates for the DNA vaccine development.
Methods![]()
This study was performed at Department of Biochemistry, Institute of Biochemistry, Biotechnology and Bioinformatics, The Islamia University of Bahawalpur, Pakistan.
Selected genes, vector and bacterial strains
All the selected genes were obtained in cloned pET vector form through BeiResources, National Institute of Allergy and Infectious Diseases (NIAID), USA. pVAX11™ (Catalog # V260-20) used as mammalian expression vector and the bacterial strain DH5α used for transformation was purchased from Invitrogen, Thermo Fisher Scientific™, USA.
Primer Designing
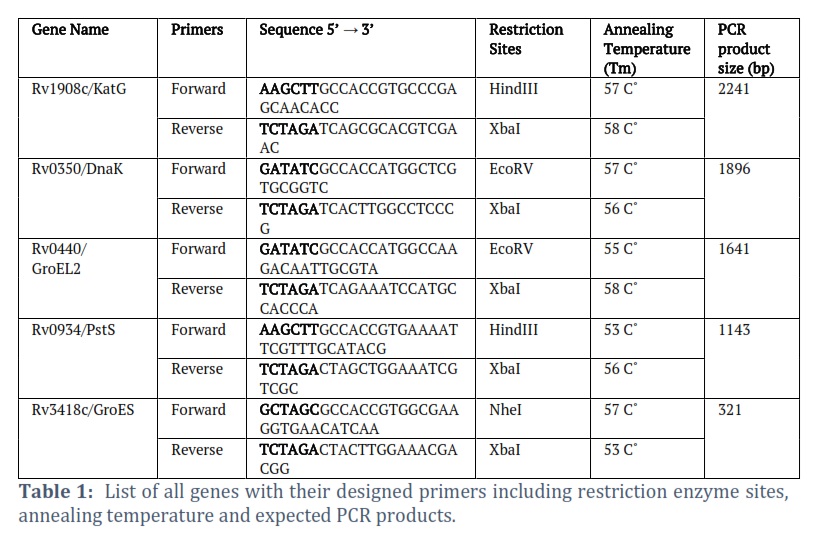
Primers were designed by adding appropriate restriction sites in both forward and reverse primers by flanking the start and stop codons. Kozak consensus sequence was also inserted in forward primers of each gene prior to Restriction site for appropriate initiation of translation. The complete sequence along with restriction sites of all primers are given in Table # 1.
Expression cloning of the selected genes
All the selected genes were amplified through Polymerase Chain Reaction by using DreamTaqᵀᴹGreen PCR–Master Mix (Catalog # K1081) from Thermo Scientific. The PCR profile comprised an initial denaturation step at 95°C for 5 minutes, succeeded by 35 cycles of denaturation at 95°C for 45 seconds, annealing at a specific Tm (as indicated in the Table # 1) for 1 minute, and extension at 72°C for 1 minute. A final extension was conducted at 72°C for 10 minutes. PCR reaction of Control DNA (700 bp) with its Control Primers was also run parallel to all gene reactions as positive control. All amplified genes and pVAX1 vector were double digested with their specific restricted endonucleases enzymes and purified. Double digested pVAX1 were also treated with rSAP enzyme to prevent it from self-ligation. After confirmation and quantification of double digested genes and vector, both were ligated to each other using T4 DNA ligase enzyme.
Once the accomplishment of ligation was confirmed through electrophoresis, the ligated product was immediately transformed into competent cells which were prepared by TSS method, following the protocols given by Chung et al [20]. The transformed ligated products were spread over kanamycin positive respective labelled plates one by one. Positive and negative control plates were also structured and incubated at 37˚C for overnight. Next morning the plates were observed, and transformation was confirmed through appearance of colonies on plates.
A single colony from the center of each plate was picked and added in kanamycin positive autoclaved LB media and placed in shaking incubator for overnight. Next day the growth was observed, and plasmid extraction was performed using Thermo Scientific™ GeneJET Plasmid Miniprep Kit catalog#K0502. The extracted plasmid was confirmed by agarose gel electrophoresis.
Clone Confirmation though PCR And Restriction Digestion
Colony PCR as well PCR of extracted plasmid was done using both gene and pVAX1 specific primers for confirmation of constructs. Further, these constructs were also confirmed through single and double digestion with different restriction endonuclease enzymes.
Sequencing of all positive Constructs
Final confirmation of the positive clones was done, by sending purified plasmid constructs to BGI Tech Solutions Hongkong for DNA sequencing. After sequence analysis, clones which have the right sequence were selected and their glycerol stocks were made.
Results![]()
All genes PCR amplification
All the selected Mycobacterium specific genes were amplified by PCR using sequence specific complementary reverse and forward primers. The PCR conditions and annealing temperature of primers were optimized. The result of PCR amplification of all the selected genes are shown in Figure 1.
Restriction Digestion
The amplified genes and extracted pVAX1vector were double digested with specific restriction digestion enzymes which are mentioned in table no. 1. pVAX1was also restricted with each single enzyme parallelly to check the enzyme activity as positive control, shown in Figure # 2, 3 & 4.
Ligation and transformation
After purification and quantification of double restriction digestion of gene and vector, both were ligated to each other using T4 DNA ligase enzyme. Quantification was done by using EPOCH microplate spectrophotometer through nanodrop. The ligated products were immediately transformed into competent cells and spread over kanamycin positive agar plates overnight. Self-ligated pVAX1 plasmid was transformed as positive control while linear single cut pVAX1 plasmid was transformed as negative control, shown in Figure # 5.
Confirmation of Constructs though Restriction Digestion and PCR
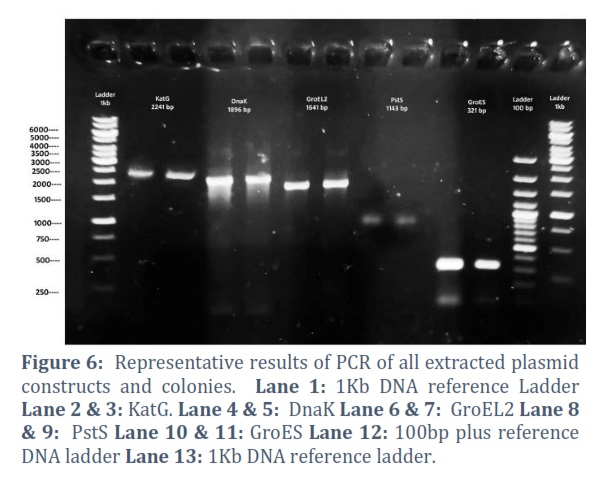
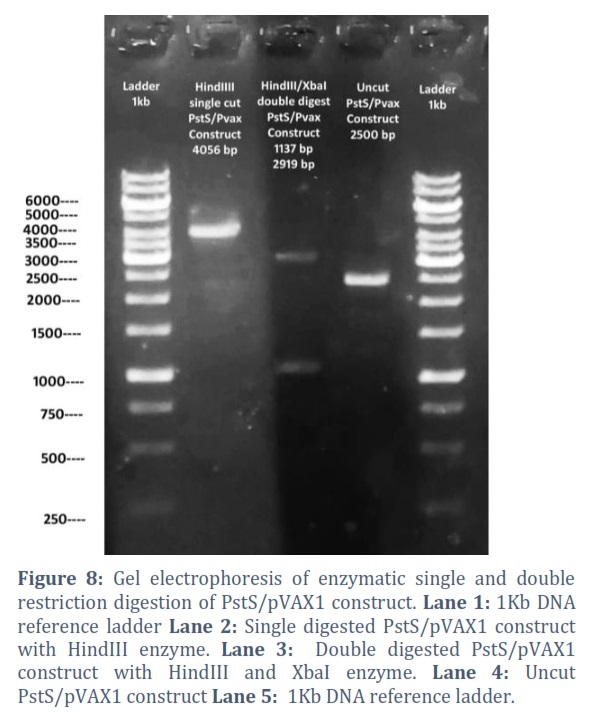
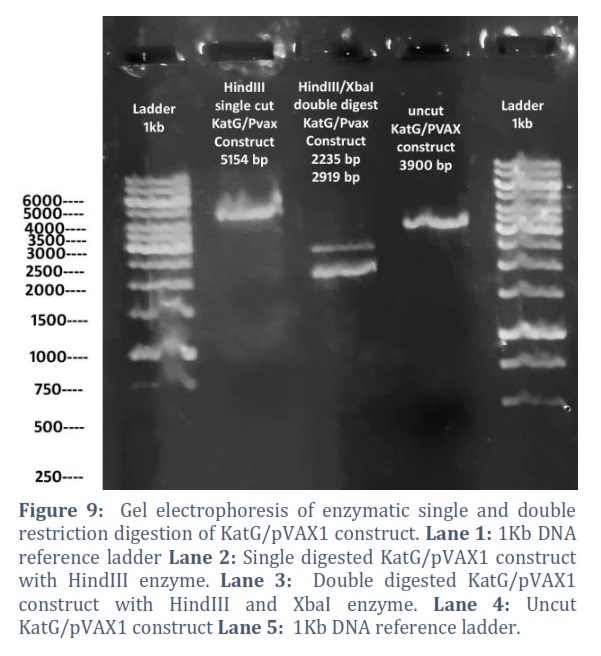
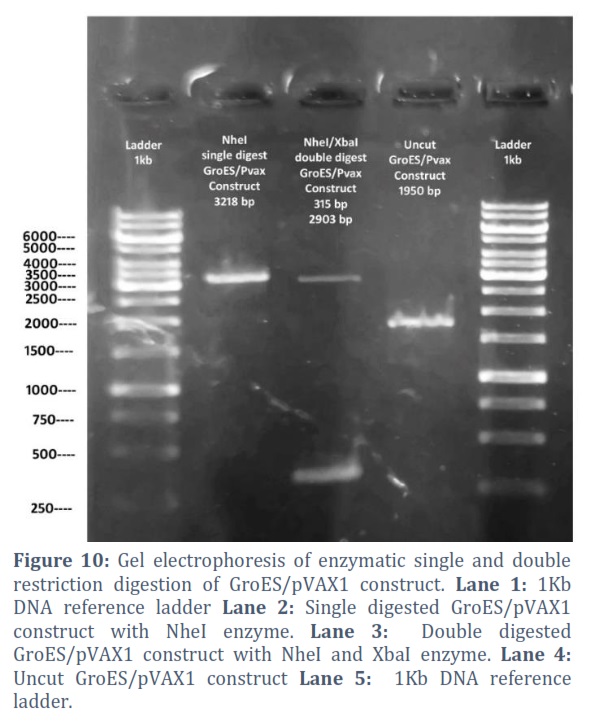
The extracted constructs were confirmed through PCR using both gene and pVAX1 specific primers, shown in Figure # 6. Single and double digestion with different restricted enzymes was performed to further confirm the constructs and successful transformation of desired gene into the vector, shown in Figure # 7-10.
Sequence Analysis of Constructs
Proper orientation of the specific gene into the vector was finally verified through sequence analysis performed by BGI Tech Solutions Hongkong.
Figures & Tables
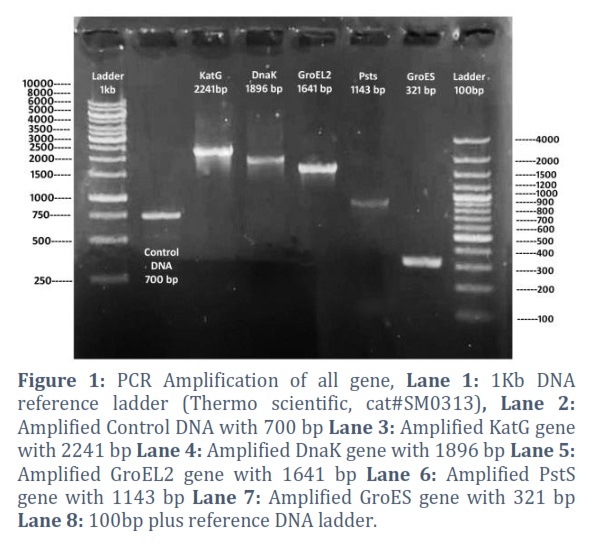
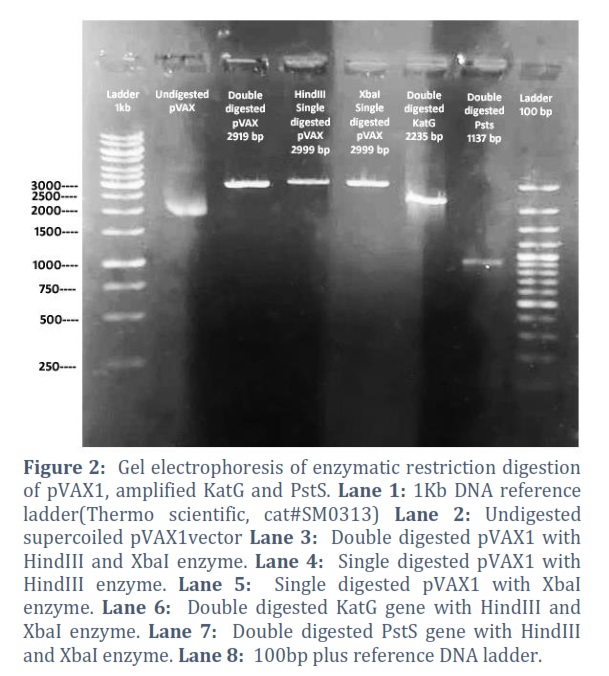
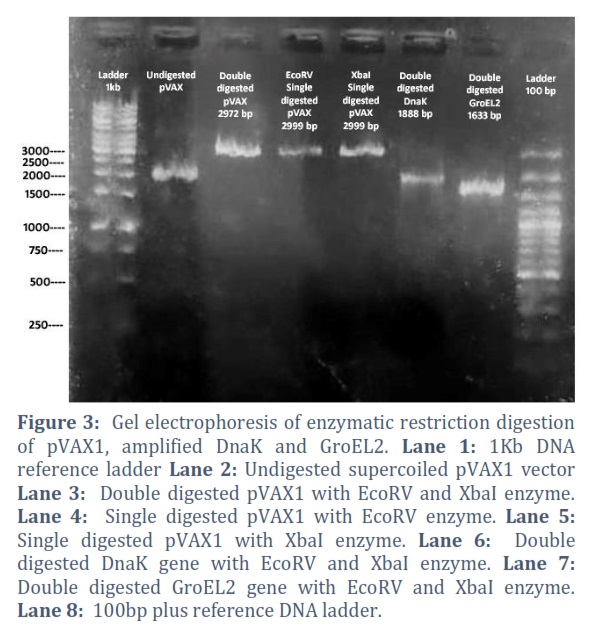

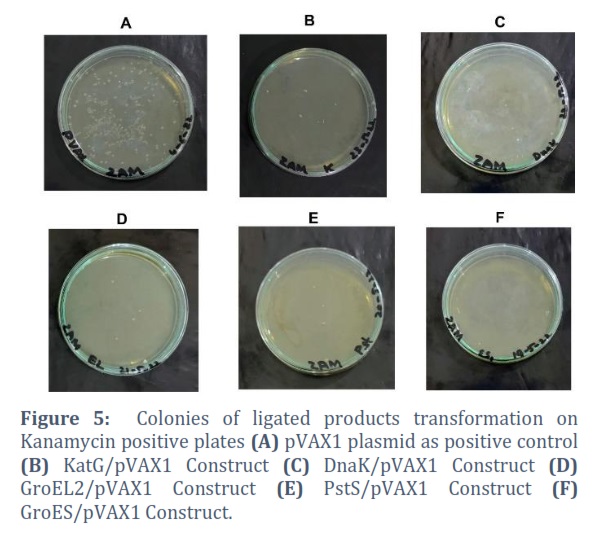
Reduction of tuberculosis incidence and development of modern and more effective drugs as well as vaccine against tuberculosis is the main target of WHO tuberculosis eradication program [21]. As the whole genome sequencing of M. tb is completed, more than hundred DNA vaccines have been designed and studied in various animal models but the protective antigen for TB is still not clear [22]. This deficit has created the limitation to the development of tuberculosis vaccines [23].
Formerly, various M. tb secreted proteins were regarded as suitable candidate vaccines but not all were effective. Some DNA vaccine encoding genes such as Mpt32, Bfrb, esat6 were produced and have been reported to stimulate the antibody response [24,25].
Another study including the development of subunit-based DNA vaccines was performed by using various genes of TB like cfp10, hspx, ag85a, ag85b and ag85c, which reports very strong immune response in mouse model in alone or combination with BCG [26]. Therefore, many gene encoding vaccine have shown very promising level of protection in different animal models and can be suitable candidate for TB vaccine.
In the present study, five genes related to mycobacterium were used for the designing and production of DNA vaccine including Rv1908c/KatG, Rv0440/GroEL2, Rv3418c/GroES, Rv0350/DnaK and Rv0934/PstS. All these genes were reported to have important biological functions and are structural parts of the cell, suggesting being very significant for survival of bacterium and also for taken to be as antigen [27-31]. In recent studies Rv1908c/KatG was selected and used by different groups and proved the importance of this gene in bacterial pathogenesis and DNA vaccine. Mycobacterial enoyl reductase inhibition as clinically evidenced by isoniazid, the most potent TB drug, KatG provides the most effective methods for eliminating M. tb [32]. In another study recombinant DnaK (Rv0350) and MPT83(Rv2873) gene constructs was tested on TB patients and found efficient in provoking the immune response against M. tb. Through genetic recombination and protein purification technology, these two antigenic proteins were cloned and purified. Blood samples were obtained from TB patients, non-TB patients with other pulmonary diseases and healthy subjects. In the immunological properties of stimulating humoral and cellular immunity, the results revealed that the sensitivity and specificity of both MPT83 and DnaK proteins were 76. 30% & 58. 95% and 77. 80% & 66. 67% respectively [33]. Another study presents cloning the genes of the immunodominant antigen of M. tb i.e. PstS and its T cell epitopes (PstS1p). This study revealed the immunity and immunogenicity of both PstS1 and its epitope PstS1p, which significantly produced higher antigen-specific IgG antibodies in mice serum, indicating increased antibody response. They also promoted proliferation of CD4+ T cell by stimulating the T helper 1 response, indicating the cruciality of this antigen as promising DNA vaccine candidate for controlling tuberculosis [34].
This study was victorious in cloning of five selected genes of Mycobacterium into mammalian expression vector pVAX1and making the very desirable candidate as DNA vaccines. These gene encoding constructs may be used in various animal models to verify the immunization and also for the challenge studies against TB in future experiments.
Conflict of Interest
The authors have no conflicts of interest to declare.
Muhammad Mohsin Zaman: Conceived idea, conducted the research, collected, analyzed and interpreted the data and wrote the article.
Mirza Imran Shahzad: Planned and supervised the research, edited and approved the final manuscript
Aeman Jilani: Helped in conducting research and writing of article, collected, analyzed and interpreted the data.
Areeba Yousaf: Helped in conducting research and writing of the article.
Ansah Bashir: Helped in conducting research and writing of article
Saher Riaz: Helped in conducting research and writing of article
Gildardo Rivera Sanchez: Financial support the research, edited and approved the final manuscript.
![]() References
References
- Whitlow E, Mustafa AS, Hanif SNM. An Overview of the Development of New Vaccines for Tuberculosis. Vaccines, (2020); 8(4): 586.
- Organization WH (2021) Global Tuberculosis Report 2020. Geneva, Switzerland; WHO; 2021. ISBN 978-92-4-003702-1.
- MacNeil A, Glaziou P, Sismanidis C, Date A, Maloney S, et al. Global epidemiology of tuberculosis and progress toward meeting global targets—worldwide, 2018. Morbidity and Mortality Weekly Report, (2020); 69(11): 281.
- Lu J, Wang C, Zhou Z, Zhang Y, Cao T, et al. Immunogenicity and protective efficacy against murine tuberculosis of a prime-boost regimen with BCG and a DNA vaccine expressing ESAT-6 and Ag85A fusion protein. Clinical and Developmental Immunology, (2011); 2011(1): 1-10.
- Tang D-c, DeVit M, Johnston SA. Genetic immunization is a simple method for eliciting an immune response. Nature, (1992); 356(6365): 152-154.
- Huygen K. Plasmid DNA vaccination. Microbes and infection, (2005); 7(5-6): 932-938.
- Sun R, Skeiky YA, Izzo A, Dheenadhayalan V, Imam Z, et al. Novel recombinant BCG expressing perfringolysin O and the over-expression of key immunodominant antigens; pre-clinical characterization, safety and protection against challenge with Mycobacterium tuberculosis. Vaccine, (2009); 27(33): 4412-4423.
- Montagnani C, Chiappini E, Galli L, de Martino M. Vaccine against tuberculosis: what’s new? BMC infectious diseases, (2014); 14(1): 1-9.
- Arroyo L, Marín D, Franken KL, Ottenhoff TH, Barrera LF. Potential of DosR and Rpf antigens from Mycobacterium tuberculosis to discriminate between latent and active tuberculosis in a tuberculosis endemic population of Medellin Colombia. BMC Infectious Diseases, (2018); 181-9.
- Hamada Y, Cirillo DM, Matteelli A, Penn-Nicholson A, Rangaka MX, et al. Tests for tuberculosis infection: landscape analysis. European Respiratory Journal, (2021); 58(5).
- Tameris M, Mearns H, Penn-Nicholson A, Gregg Y, Bilek N, et al. Live-attenuated Mycobacterium tuberculosis vaccine MTBVAC versus BCG in adults and neonates: a randomised controlled, double-blind dose-escalation trial. The lancet Respiratory medicine, (2019); 7(9): 757-770.
- Spertini F, Audran R, Chakour R, Karoui O, Steiner-Monard V, et al. Safety of human immunisation with a live-attenuated Mycobacterium tuberculosis vaccine: a randomised, double-blind, controlled phase I trial. The lancet Respiratory medicine, (2015); 3(12): 953-962.
- Bello S, Afolabi RF, Ajayi DT, Sharma T, Owoeye DO, et al. Empirical evidence of delays in diagnosis and treatment of pulmonary tuberculosis: systematic review and meta-regression analysis. BMC public health, (2019); 19(1): 1-11.
- Mu J, Jeyanathan M, Small C-L, Zhang X, Roediger E, et al. Immunization with a bivalent adenovirus-vectored tuberculosis vaccine provides markedly improved protection over its monovalent counterpart against pulmonary tuberculosis. Molecular Therapy, (2009); 17(6): 1093-1100.
- Mobed A. DNA Based vaccines against Mycobacterium tuberculosis: recent progress in vaccine development and delivery system. Iranian Journal of Immunology, (2020); 17(4): 255-274.
- Kowalewicz-Kulbat M, Locht C. Recombinant BCG to enhance its immunomodulatory activities. Vaccines, (2022); 10(5): 827.
- Arlehamn CSL, Lewinsohn D, Sette A, Lewinsohn D. Antigens for CD4 and CD8 T cells in tuberculosis. Cold Spring Harbor perspectives in medicine, (2014); 4(7): a018465.
- Buzitskaya Z, Stosman K, Khairullin B, Kassenov M, Nurpeisova A, et al. A new intranasal influenza vector-based vaccine TB/FLU-04L against tuberculosis: preclinical safety studies. Drug Research, (2022); 72(05): 255-258.
- Fomsgaard A, Liu MA. The key role of nucleic acid vaccines for one health. Viruses, (2021); 13(2): 258.
- Chung C, Niemela SL, Miller RH. One-step preparation of competent Escherichia coli: transformation and storage of bacterial cells in the same solution. Proceedings of the National Academy of Sciences, (1989); 86(7): 2172-2175.
- Sefidi-Heris Y, Jahangiri A, Mokhtarzadeh A, Shahbazi M-A, Khalili S, et al. Recent progress in the design of DNA vaccines against tuberculosis. Drug Discovery Today, (2020); 25(11): 1971-1987.
- Fan X, Gao Q, Fu R. Differential immunogenicity and protective efficacy of DNA vaccines expressing proteins of Mycobacterium tuberculosis in a mouse model. Microbiological research, (2009); 164(4): 374-382.
- Ilghari D, Lightbody KL, Veverka V, Waters LC, Muskett FW, et al. Solution structure of the Mycobacterium tuberculosis EsxG· EsxH complex: functional implications and comparisons with other M. tuberculosis Esx family complexes. Journal of Biological Chemistry, (2011); 286(34): 29993-30002.
- Shahzad MI, Shahid N, Sial N, Hasanat A, Khalid M, et al. Evaluation of DNA vaccines encoding M. Tb gene Bfrb and Mpt32 in mice model. Pakistan journal of pharmaceutical sciences, (2017); 30(5 (Supplementary)): 2025-2029.
- Shahzad MI, Khan IH, Luciw PA, Gulfraz M, Khanum A. Cloning, Expression and Genetic Immunization Studies of Mycobacterium tuberculosis Gene esat6. Pakistan Journal of Zoology, (2013); 45(3).
- Shahzad M. DNA vaccine against tuberculosis by using five Mycobacterium specific genes. International Journal of Mycobacteriology, (2021); 10(5): 4-4.
- Munir A, Wilson MT, Hardwick SW, Chirgadze DY, Worrall JA, et al. Using cryo-EM to understand antimycobacterial resistance in the catalase-peroxidase (KatG) from Mycobacterium tuberculosis. Structure, (2021); 29(8): 899-912. e894.
- Lopes RL, Borges TJ, Araújo JF, Pinho NG, Bergamin LS, et al. Extracellular mycobacterial DnaK polarizes macrophages to the M2-like phenotype. PLoS One, (2014); 9(11): e113441.
- Vinod V, Pushkaran AC, Kumar A, Mohan CG, Biswas R. Interaction mechanism of Mycobacterium tuberculosis GroEL2 protein with macrophage Lectin-like, oxidized low-density lipoprotein receptor-1: An integrated computational and experimental study. Biochimica et Biophysica Acta (BBA)-General Subjects, (2021); 1865(1): 129758.
- Esparza M, Palomares B, García T, Espinosa P, Zenteno E, et al. PstS‐1, the 38‐kDa M ycobacterium tuberculosis Glycoprotein, is an Adhesin, Which Binds the Macrophage Mannose Receptor and Promotes Phagocytosis. Scandinavian journal of immunology, (2015); 81(1): 46-55.
- Maurya VK, Singh K, Sinha S. Suppression of Eis and expression of Wag31 and GroES in Mycobacterium tuberculosis cytosol under anaerobic culture conditions, (2014); 52(08): 773-780.
- Ofori-Anyinam N, Hamblin M, Coldren ML, Li B, Mereddy G, et al. KatG catalase deficiency confers bedaquiline hyper-susceptibility to isoniazid resistant Mycobacterium tuberculosis. bioRxiv, (2023); 2023.2010. 2017.562707.
- LI X, XIAO T, LI M, LIU H, LI S, et al. Evaluation of the antigenicity of Dnak and MPT83 proteins of Mycobacterium tuberculosis. Chinese Journal of Microbiology and Immunology, (2019); 106-113.
- an X, Li X, Wan K, Zhao X, Deng Y, et al. Construction and immunogenicity of a T cell epitope-based subunit vaccine candidate against Mycobacterium tuberculosis. Vaccine, (2021); 39(47): 6860-6865.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0