

Full Length Research Article
Metal Sulfates and Microbial Food Treatment Alleviate the Oxidative Damage Caused by PEG-Induced Osmotic Stress in Cotton Plants
Semra Almas1, Tijen Demiral Sert1,2*
Adv. life sci., vol. 11, no. 3, pp. 572-579, August 2024
*– Corresponding Author: Tijen Demiral Sert (tijensert@sdu.edu.tr)
Authors' Affiliations
2. Current address: Department of Biology, Faculty of Engineering and Natural Sciences, Süleyman Demirel University – Türkiye
[Date Received: 23/01/2023; Date Revised: 12/05/2024; Date Published: 10/07/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Drought stress restricts cotton production, which causes a considerable loss of cotton yields worldwide. Plant activators improve crop production and ameliorate abiotic stresses. Thus, the effects of plant activators on the genotype of cotton (Gossypium hirsutum L.) Candia, which is resistant to drought, were investigated.
Methods: Plants were treated with the plant activators, and then osmotic stress was initiated by 30% PEG 6000 treatment for 13 days. Antioxidant enzyme activities such as superoxide dismutase (SOD), ascorbate peroxidase (APX), and peroxidase (POX) were measured along with growth, relative water content (RWC), osmotic potential, photosynthetic pigments, ion leakage, proline accumulation, malondialdehyde (MDA) content, and reactive oxygen species (ROS).
Results: Osmotic stress reduced shoot fresh weight (FW) and osmotic potential but increased root length of Candia. Activator treatment alleviated these effects. Photosynthetic pigment contents and cell membrane permeability were not affected by osmotic stress and activator treatments; proline accumulation and lipid peroxidation levels were enhanced by osmotic stress, but activators alleviated these increments. Activity bands for the isozymes Mn-SOD1, SOD2, and SOD3, as well as the isozyme Fe-SOD, were found in Candia. Osmotic stress elevated APX activity in Candia leaves, while activator treatment decreased it. POX activity was not affected by osmotic stress, but activator treatment decreased POX activity. Osmotic stress-induced accumulation of ROS was lessened by activator treatment.
Conclusion: Plant activator treatment reduced the amount of oxidative damage (ROS buildup and lipid peroxidation) caused by drought stress in the Candia genotype.
Keywords: Antioxidant system; Cotton; Gossypium hirsutum L.; Osmotic stress; PEG 6000; Plant activators
Introduction![]()
One of the most vital crops in the world, cultivated cotton (Gossypium spp. Malvaceae family) is the second-most significant oil seed crop and the world's top producer of natural fiber used in the textile industry. One of the main causes of the nearly 10%–15% reduction in plant yield and slowdown of cotton growth is drought stress [1]. Moreover, regions subjected to severe water stress cultivate 57% of the world’s cotton [2]. Thus, developing drought-tolerant cotton cultivars, increasing water usage efficiency, and creating innovative biosafe technologies are crucial for a sustainable cotton sector.
Plants respond to drought by sensing changes in the lack of water status in the roots. Hydraulic signaling begins with the formation of a hydraulic signal as a result of changes in Ψw (water potential), which can be caused by variations in turgor, Ψs (osmotic potential), or water tension [3]. This is because the water potential of the leaves and the roots differ. When the stress signal reaches the leaves, the plant closes its stomata, going into a water-saving mode. Reduced NADP+ regeneration during the Calvin cycle resulting from stomatal closure under water deficit conditions will cause an over-reduction of the photosynthetic electron transport chain. Consequently, electrons from the photosystem I (PSI) acceptor side are transferred to O2 due to the leakage of photosynthetic electrons to the Mehler reaction, resulting in the production of hydrogen peroxide (H2O2) and superoxide anion (O2•−) [4]. These lethal ROS and its products, singlet oxygen (1O2), superoxide, H2O2, and hydroxyl radicals (•OH), react with lipids, proteins, and nucleic acids to cause lipid peroxidation, protein denaturation, DNA mutation, oxidative stress, and eventually cell death [5, 6]. By triggering their antioxidant defense system, plants respond to excess production of reactive oxygen species (ROS) by averting an oxidative explosion. Defensive enzymes such as glutathione reductase (GR), catalase (CAT), ascorbate peroxidase (APX), peroxidase (POX), and superoxide dismutase (SOD) as well as non-enzymatic antioxidants such as vitamin C, vitamin E, glutathione, and carotenoids are involved in the process [7].
The Food, Agriculture, and Health Organization states that low- and middle-income countries have lost 34% of their crops due to drought, costing $37 billion (FAO 2021). New methods must be developed to meet the population's 3 billion tons of grain needs, which will reach approximately 9 billion people by 2050 [8]. These methods should ensure the development of products that increase yield, provide resistance to abiotic and biotic stress factors, improve food quality, reduce environmental pollution, or have the most negligible impact on it [9]. A new approach in this area to strengthen the defense mechanism of plants has emerged by using plant biological stimulants or plant activators [10, 11]. To support plant growth, development, yield, defense against pathogens, and stress tolerance, this method externally applies small amounts of organic or inorganic compounds and/or microorganisms to the plant [11-14]. Because they are a practical and efficient tool for enhancing plant productivity and yield by maintaining the plant metabolism at its most productive level, researchers, farmers, and industrial companies have expressed great interest in using bio stimulants or plant activators based on natural substances. Many scientific studies have shown their novel aspects to improve crop production and ameliorate abiotic stresses [11-14].
We have decided to use two commercially available plant activators called Soil-Set® and Crop-Set® (Alltech) because they offer a level of standardization in terms of concentration, purity, and formulation, reducing variability and facilitating result interpretation. The bioactive ingredients of both plant activators were copper sulfate, iron sulfate, manganese sulfate, and zinc sulfate (only in Soil-Set®) besides bacterial fermentation media (microbial food) and plant extract (natural surfactant). To determine the impact of the use of the plant activators on improving the drought tolerance of cotton, we deployed a drought-tolerant cotton genotype, Candia. We mimicked osmotic stress by PEG-6000 treatment. Here, we hypothesize that the application of a plant activator treatment reduced the extent of oxidative damage caused by PEG-6000-induced osmotic stress in the Candia genotype.
Methods![]()
Plant material, growing conditions, and plant activator treatments
The seeds of the cotton (Gossypium hirsutum L.) Candia genotype were provided by the Şanlıurfa Agricultural Research Institute. Seedlings were grown in a fully automatic climate chamber, the temperature of which was set at an average of 27 ± 1 °C per day (30 °C in the light phase / 24 °C ± 1 °C in the dark phase). The growth chamber had a lighting cycle of 16 h light / 8 h dark, a light intensity of 350 µmol m-2 sec-1, and around 60% humidity level.
The perlite-filled pots were divided into two groups, the activator (A) and control (C), and the pots of the A group were treated with Soil-Set® dissolved in deionized water at the recommended dose by the manufacturer. In contrast, the pots of the C group were treated with deionized water alone until the seeds were germinated. Seedlings were grown until the 5-6 leaf stage (first stage), and then Crop-Set®, dissolved in deionized water at the recommended dose by the manufacturer, was started to be applied to the leaves and the perlite of cotton plants in the A group (second stage). Ten days after activator treatments, plants were harvested (Day 0), and osmotic stress was initialized (third stage) by applying PEG 6000 in ½ Hoagland's solution to half of the pots of both C and A groups, and thus four treatment groups were formed (C, A, PEG, A+PEG). Osmotic stress treatment was applied by gradually increasing the concentration of PEG 6000 from 10% at the first application to 20% at the second time and then finally to 30% and the final harvest was performed 13 days after the 30% PEG 6000 application.
Growth parameters and leaf relative water content (RWC)
The lengths, fresh and dry weights of the roots, and shoots of randomly selected ten plants from each experimental group were measured for growth parameters. For leaf RWC, 6 leaf samples from each group were selected, and the relative water content of the leaf samples (RWC) was calculated as % [15].
Leaf osmotic potential (Ψs)
The osmotic potential was measured by an osmometer (Wescor Vapor Pressure Osmometer 5600). Osmolarity (c) values were converted to megapascals (MPa) by placing them in the Van't Hoff equation below.
Ψs (MPa) = −c (mosmoles kg−1) × 2.58 × 10−3
Photosynthetic pigment concentrations
Chlorophyll a (Chl-a), chlorophyll b (Chl-b), and carotenoid (Car) contents were measured spectrophotometrically and calculated according to Lichtenthaler and Wellburn [16].
Proline determination
Bates et al.'s method [17] for spectrophotometric determination of free proline content was followed. Using the calibration curve, proline concentration was determined and reported as µmol proline g-1 FW.
Cell membrane permeability (Ion Leakage) and MDA content
The permeability of cell membranes was measured and computed per Dionisio-Sese and Tobita [18]. Quantification of MDA, the final product of lipid peroxidation, was performed according to Madhava-Rao and Sresty [19]. The content of MDA was estimated using the extinction coefficient (ε = 155 mM-1 cm-1).
Antioxidant enzyme activity assays
Identification of SOD isoenzymes
Using a mortar, pestle, and liquid nitrogen, leaf samples were ground into a powder. After that, they were homogenized using 50 mM Na-phosphate buffer (pH 7.8) containing 2% w/v polyvinylpolypyrrolidone (PVPP) and 1 mM EDTA. The homogenates were centrifuged for 30 minutes at 4 °C at 14,000 rpm, and the supernatants were utilized for enzyme activity tests and protein analysis. Using bovine serum albumin as a standard, the total protein content was determined following Bradford [20].
Native polyacrylamide gel electrophoresis (PAGE) was used to separate samples with equal protein concentrations (10 mg well-1) in 5% stacking and 12% separating gels at 4 °C with a constant current of 120 mA [21]. Using nitro blue tetrazolium (NBT) and riboflavin for photochemical labeling, SOD isozymes were identified. [22]. Each SOD isozyme’s unit activity was determined by calculating the SOD standard from bovine liver (Sigma Chemical Co.). Inhibitors were added to the staining solution in order to improve the characterization of the various types of SOD. According to Vitória et al., [23], these included 2 mM KCN, which inhibits Cu/Zn-SOD activity, and 3 mM H2O2, which inhibits both Cu/Zn-SOD and Fe-SOD activities. Both inhibitor treatments do not affect Mn-SOD activity. Four independent replicates were used for the experiments, and the SOD activities of the activator or control plants were assumed to be 100%. The percentage of control values for each treatment is displayed. The means having the same letter within each isoenzyme do not differ substantially at P < 0.05 as measured by LSD.
APX and POX activities
APX activity was assayed by monitoring the decrease in the assay medium at 290 nm for 180 seconds [24]. Ascorbate oxidized per minute given as mmol mL-1 represents the unit of APX activity.
Peroxidase (POX) enzyme activity was assayed according to Herzog and Fahimi [25]. For three minutes, the rise in absorbance at 465 nm was monitored, and μmol H2O2 decomposed mL-1 min-1 was used to express the POX activity.
Determination of H2O2 and superoxide (O2•−) accumulation levels
The H2O2 accumulation level was measured according to Bernt and Bergmeyer [26]. Superoxide (O2•−) buildup was measured in accordance with Xu et al., [27].
Statistical analysis
All of the data was subjected to a one-way analysis of variance (ANOVA), and the changes in the mean values were compared using the LSD (Lowest Standard Deviations) test. Each data point was the mean ± standard error of ten repeats (n = 10) for growth parameters and six (n = 6) for other parameters. Significant differences were detected between comparisons with a P < 0.05. The standard version of SPSS for Windows, Release 11.0-, was used to conduct statistical analyses.
Results![]()
Growth, leaf water status, and photosynthetic pigment contents
30% PEG 6000-induced osmotic stress increased root length by 27% while showing no significant effect on shoot length (Fig. 1a). Plant activator treatment in the presence of osmotic stress caused around 12% decrement in root length, thus lessening the osmotic stress-induced growth of roots. FW of shoots and roots showed 124% and 64% increments on day 0, whereas, on day 13, activator treatment alone caused 20% and 18% enhancements in shoot and root FW, respectively. Although osmotic stress treatment for 13 days caused a 37% decrease in shoot FW, it caused a 26% increase in root FW of Candia plants, and the decline in shoot FW was not changed significantly by activator treatment (Fig. 1b). However, enhancement in root FW induced by osmotic stress was hampered around 28% by activator treatment in the A+PEG group, almost reaching control level. Activator treatment alone caused an almost two-time increase in shoot DW while causing no change in root DW on day 0. Although osmotic stress caused a 2 times increase in root DW, shoot DW decreased around 8% on day 13. The root and shoot DW did not exhibit a statistically significant difference from the PEG-treated group alone in the presence of plant activators (Fig. 1c).
The activator treatment appears to mitigate some effects of osmotic stress on root biomass, reducing the increase in root FW and length induced by osmotic stress and bringing it closer to control levels. However, the activator does not significantly affect shoot biomass changes under osmotic stress, as the decline in shoot FW and DW remains largely unchanged. This suggests that while the plant activator has a moderating effect on root biomass under osmotic stress, it does not substantially alter the osmotic stress impacts on shoot biomass.
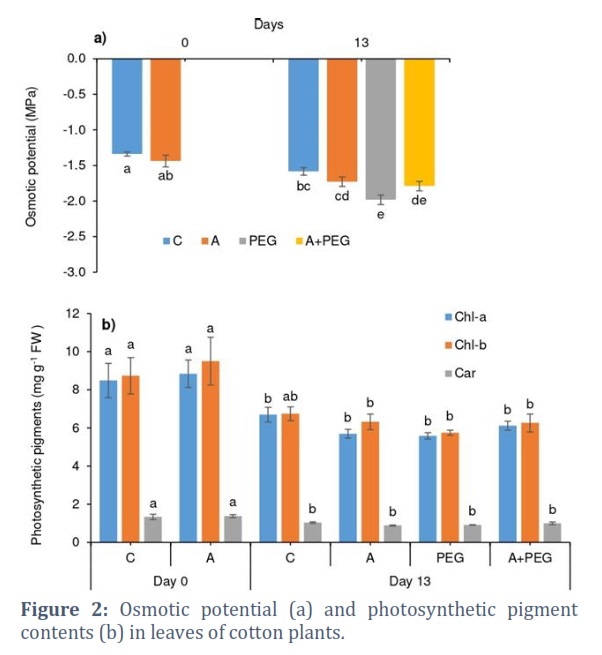
Osmotic stress diminished leaf RWC by 3%, and the activator treatment resulted in a 20% decrease in leaf RWC of Candia plants under osmotic stress (Fig. 1d). Likewise, PEG-induced osmotic stress decreased Ψs around 25% but activator treatment hampered this stress-induced decrement in Ψs (Fig. 2a). Photosynthetic pigment contents were affected by neither activator nor stress treatments except for a 15% decrease in Chl-b under PEG-induced osmotic stress (Fig. 2b).
Proline accumulation, ion leakage, and MDA content
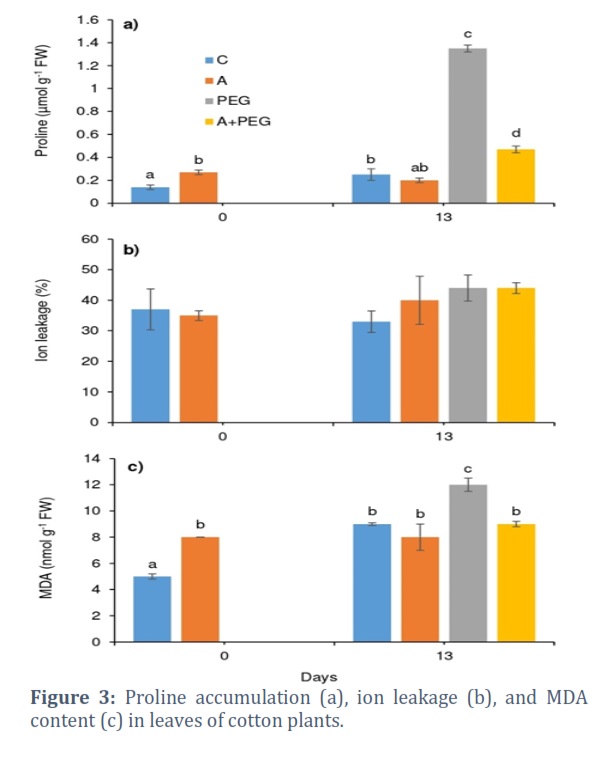
PEG-induced osmotic stress alone significantly increased free proline buildup, causing a 5.4 times increase compared to the control group (Fig. 3a). However, plant activator treatment remarkably decreased proline accumulation induced by osmotic stress, which was still 88% higher than in control plants. Osmotic stress did not significantly affect the ion leakage of Candia leaves, and plant activator treatment did not show any effect on ion leakage either (Fig. 3b). However, MDA content showed a 33% increase after osmotic stress and activator treatment completely prevented this increase in the A+PEG group (Fig. 3c).
Antioxidant enzyme activities, H2O2 and O2•− accumulation levels
Electrophoretic separation of SOD isoenzymes brought Mn-SOD1, Mn-SOD2, Mn-SOD3, and Fe-SOD isozyme activity bands into view (Fig. 4). Plant activator treatment stimulated the appearance of Mn-SOD1 and Mn-SOD2 isoenzymes on day 0 but Mn-SOD1 and Mn-SOD3 isoenzymes on day 13. Control plants exhibited Mn-SOD1 and Fe-SOD activity bands on day 13; the expression of Mn-SOD1 activity in the activator group was 77% higher than in the control group.
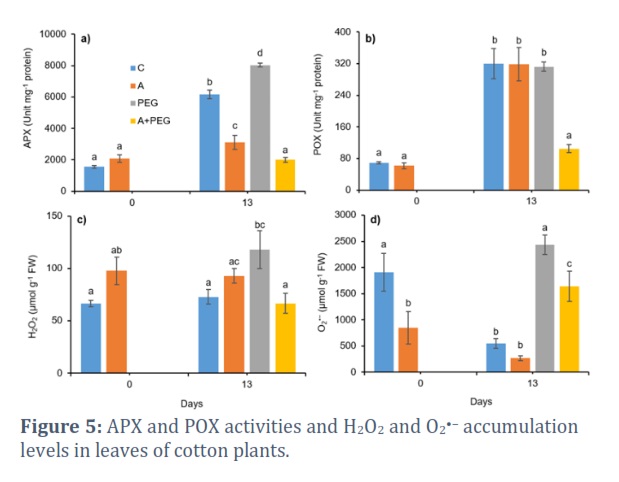
APX activity showed a 30% enhancement by PEG-induced osmotic stress, and this increase was lessened by 68% in the presence of activator treatment (Fig. 5a). APX activity of activator treatment alone was also 49% lower than the constitutive level of control conditions.
On the thirteenth day, POX activity in Candia plants demonstrated a 67% decrease in the A+PEG group in contrast to the control group (Fig. 5b).
PEG-induced osmotic stress caused 62% and 4 times increments in H2O2 and O2•− accumulations in leaves of Candia plants on day 13, respectively. However, activator treatment inhibited PEG-induced ROS accumulation, causing a 77% decrease in H2O2 and a 48% decrease in O2•− concentration when compared to PEG-induced osmotic stress treatment alone (Fig. 5c and 5d).
Figures & Tables
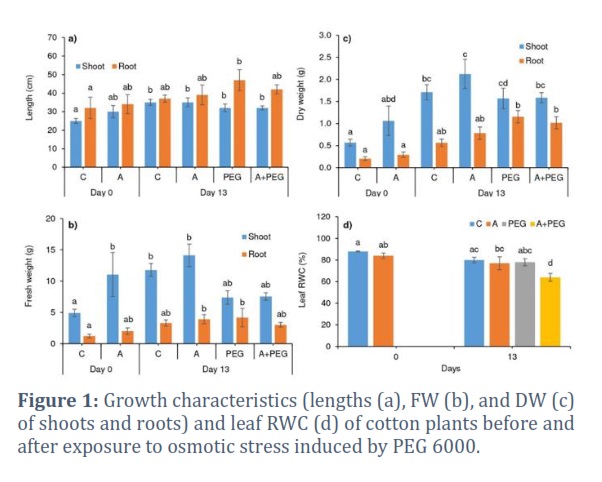

Morphological traits
The primary cause of the production decline is the plant's decreased fresh and dry weight as a result of the drought [28]. The delay of growth and reduction in plant height was identified as one of the most significant consequences of drought stress experienced by cotton plants during their growth phase [29]. Supporting this, osmotic stress decreased the FW and DW of shoots of Candia plants. In general, plants tend to be longer rooted during exposure to drought stress [30], and that was the case for Candia plants, the roots of which became longer under osmotic stress (Fig. 1a). However, plant activator treatment mitigated this stress-related symptom by preventing an osmotic stress-induced increase in root length.
Physiological traits
The leaf RWC and, consequently, the leaf osmotic potential (Ψs) decreases in higher plants during drought stress [31]. In comparison to low levels of drought stress (often about -0.2 MPa) observed in a range of plant species, an osmotic potential of -1.20 MPa was applied to Candia plants via PEG-mediated drought stress [32]. The plants adapted to arid conditions have often been found to have a high leaf RWC [33], and this was the case for drought-tolerant Candia plants that displayed high leaf RWC despite stress conditions. The production of organic solutes, including proline, to provide osmotic adjustment and ensure ongoing water absorption throughout the stress period may cause a lower value of Ψs in the leaves of Candia plants under osmotic stress [34]. Because cotton's natural growth environment regularly coexists with high temperatures and water stress, maintaining the leaf water status is essential for the plant to grow and survive.
Under a range of stress situations, free proline synthesis is a vital activity that helps balance the osmotic potency of the cytosol with that of the vacuole and outside the cell, increasing the plant's tolerance against stressors [35, 36]. Besides its role as a cytosolic organic osmolyte, proline functions as an antioxidant, chelator, energy, and nitrogen source and stabilizer for macromolecular structures and/or subcellular membranes [35, 37]. Confirming our results, Gomes et al., [38] also observed a proline accumulation in the leaves of coconut plants exposed to water shortage. Proline accumulation during water deficit has also been reported in the roots of maize [39] and in the roots and shoots of sunflower [40]. Enhanced accumulation of free proline in leaves of Candia plants under osmotic stress was remarkably prevented by the treatment with plant activators, which showed the stress-alleviating effect of plant activators. Similarly, proline concentration in tomato plants decreased under stressful conditions, as shown by Gedeon et al., [41], and increased proline accumulation occurred in the leaves of tomato plants in salinity conditions with the application of the bio stimulants. In situations where there is a water shortage, compatible osmolytes can support turgor and turgor-related functions by lowering water activity and the cell’s osmotic potential [35]. This is supported by the fact that proline accumulation in leaves of Candia plants was seen to occur in conjunction with decreasing Ψs, suggesting that proline was directly involved in osmotic adjustment in our investigation. This increased water potential provided tolerance against PEG-induced osmotic stress.
Gomes et al., [38] did not associate proline accumulation with osmoregulation when they considered growth reduction in two Brazilian Green Dwarf coconut ecotypes exposed to drought stress. However, they valued proline more for its ability to prevent membrane damage, as demonstrated by the electrolyte leakage method [38]. Supporting this, we also found an absence of membrane damage in the leaves of Candia plants after PEG-induced water deficit (Fig. 3b), while a significant accumulation of proline was observed (Fig. 3a). Still, enhanced peroxidation of lipid membranes after exposure to osmotic stress in Candia plants seems to be related to the enhanced production of ROS (Fig. 3c).
Antioxidant enzyme activities and ROS accumulation
Plants have evolutionarily deployed enzymatic and non-enzymatic antioxidants to tightly regulate ROS production/elimination to avoid the harms of extreme environmental conditions. SOD constitutes the first obstacle against ROS, and this enzyme dismutates superoxide anion radical (O2•−) into hydrogen peroxide (H2O2) [42]. Following the SOD reaction, H2O2 is enzymatically reduced into H2O and O2 by enzymes such as CAT, POX, and APX [4].
Plant activator treatment stimulated the occurrence of Mn-SOD1 and Mn-SOD2 izozymes on day 0, but Mn-SOD1 and Mn-SOD3 izozymes on day 13. The application of C. oxyacantha extract as a bio stimulant by Naboulsi et al., [43] resulted in a similar increase in the antioxidant activities of tomato plants, including SOD. Additionally, Abou-Sreea et al., [44] demonstrated that silymarin-enriched honeybee treatments and foliar feeding of honeybees increased SOD activity in chili pepper plants. No detection of SOD isoenzymes (which does not necessarily imply the absence of SOD activity) in PEG and A+PEG groups corroborated the enhanced accumulation of O2•− in leaves of Candia plants in both groups on both days (Fig. 5d). Still decreased accumulation of superoxide in the A+PEG group compared to the PEG group might imply the removal of superoxide radicals by other antioxidants rather than SOD (such as phenols). Decreased activities of APX in both A and A+PEG groups and of POX in A+PEG group on day 13 suggest that activator treatment might have prevented oxidative damage to major biomolecules, which in turn reduced the peroxidation of lipid membranes and rendered the activities of the antioxidant enzymes. A similar result was also obtained by Abdalla [45] that the treatment of rocket plants with a bio stimulant (the extract of Moringa oleifera) under water stress decreased the activities of CAT, POX, and SOD enzymes and they attributed this to the protection ability of the extract of Moringa oleifera against oxidative damage by favoring the accumulation of ascorbic acid and phenols. Reduced accumulation levels of H2O2 and O2•− under PEG-induced osmotic stress further support the protective effect of plant activator treatment. Our findings were validated by the fact that millet seedlings subjected to drought stress were more likely to accumulate ROS like O2•− and H2O2, and that treatment with 100 mg L−1 humic acid dramatically reduced the rate of ROS formation [30].
Here, we hypothesize that the application of plant activators reduced the amount of oxidative stress-induced damage to cotton plants. Thus, the application of plant activator to Candia plants reduced the degree of oxidative damage brought on by PEG-induced osmotic stress and consequently did not result in any increase in SOD, APX, or POX activities, nor did it raise the content of H2O2 or lower the level of O2•−. Similar findings were reported by Demiral and Turkan [46], who found that exogenous glycine betaine (GB) application reduced lipid peroxidation levels and the activities of SOD, APX, CAT, and GR enzymes in a salt-tolerant rice cultivar under salt stress. They also proposed that GB protection was ensured by its ability to prevent oxidative damage and preserve water homeostasis under salinity stress. Thus, plant activators may be used to improve agriculturally important plants’ ability to tolerate drought stress, which is particularly pertinent as we face the global challenge of food security in the context of a changing climate.
Acknowledgments
This study was financially supported by Harran University Scientific Research Center (HÜBAK), Project No. 14175, and the results obtained from S.A.'s MSc. thesis conducted in the Department of Biology, Faculty of Arts and Sciences, Harran University, Şanlıurfa, Türkiye. Thanks are due to Dr. Nimet Kılınçoğlu for this valuable discussion.
Conflict of Interest
The authors have no conflicts of interest to declare. The authors declare no affiliations or relationships with the commercial company whose products were used in this study that could be construed as a potential conflict of interest.
SA carried out the experiments and prepared the first draft of the manuscript. TDS conceived the study, supervised the experiments, acquired funding, performed statistical analysis, and wrote and edited the final version of the manuscript. Both authors have approved the final manuscript.
![]() References
References
- Pettigrew WT. Physiological Consequences of Moisture Deficit Stress in Cotton. Crop Science, (2004); 44:1265–1272.
- Kim JH, Castroverde CDM, Huang S, Li C, Hilleary R. Increasing the Resilience of Plant Immunity to a Warming Climate. Nature, (2022); 607(7918): 339-344
- Christmann A, Grill E, Huang J. Hydraulic Signals in Long-Distance Signaling. Current Opinion in Plant Biology, (2013); 16: 293–300.
- Dumanovic´ J, Nepovimova E, Natic´ M, Kucˇa K and Jac´evic´ V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Frontiers in Plant Science, (2021); 11: 552969.
- Turkan I, Demiral T. Recent Developments in Understanding Salinity Tolerance. Environmental and Experimental Botany, (2009); 67(1): 2-9.
- Demiral T, Turkan I, Sekmen AH. Signaling Strategies during Drought and Salinity, Recent News. Advances in Botanical Research, (2011); 57: 293-317.
- Zabalza A, Van Dongen JT, Froehlich A. Regulation of Respiration and Fermentation to Control the Plant Internal Oxygen Concentration. Plant Physiology, (2008); 149: 1087-1098.
- Le Mouël C, Forslund A. How Can We Feed the World in 2050? Review of Responses from Global Scenario Studies. European Review of Agricultural Economics, (2017); 44(4):541-591.
- Turkusay H, Tosun N, Yildiz S, Saygili H. Effects of Plant Activators on Physiological and Morphological Parameters of Processing Tomato. Acta Horticulturae, (2009); 808: 431-436.
- Turkan I, Demiral T, Sekmen AH, Tosun N. Abiotic Stress and the Role of Plant Activators. In: Lyons TP, Jacques KA, editors: Proceedings of Alltech's 20th Annual Symposium on Nutritional Biotechnology in the Feed and Food Industries. (2004); 387-390. Nottingham University Press, England.
- Yakhin OI, Lubyanov AA, Yakhin IA, Brown PH. Biostimulants in Plant Science: A Global Perspective. Frontiers in Plant Science, (2017); 7: 2049.
- Calvo P, Nelson L, Kloepper JW. Agricultural Uses of Plant Biostimulants. Plant and Soil, (2014); 383: 3–41.
- Sun T-J, Lu Y, Narusaka M, Shi C, Yang Y-B, Wu J-X, et al. A Novel Pyrimidin-Like Plant Activator Stimulates Plant Disease Resistance and Promotes Growth. PLoS One, (2015); 10(4): e0123227.
- Dayan A, Sari N. The Effects of Different Plant Activators on Fruit and Seed Yield and Properties of Snack Pumpkin (Cucurbita pepo L.). International Journal of Scientific and Technological Research, (2019); 5(10): 1-12.
- Smart RE, Bingham GE. Rapid Estimates of Relative Water Content. Plant Physiology, (1974); 53(2):258-260.
- Lichtenthaler HK, Wellburn AR. Determination of Total Carotenoids and Chlorophylls A and B of Leaf Extracts in Different Solvents. Biochemical Society Transactions, (1983); 11: 591–592.
- Bates LS, Waldren RP, Tear ID. Rapid Determination of Free Proline for Water-Stress Studies. Plant and Soil, (1973); 39:205-207.
- Dionisio-Sese ML, Tobita S. Antioxidant Responses of Rice Seedlings to Salinity Stress. Plant Science, (1998); 135(1): 1-9.
- Madhava-Rao KV, Sresty TVS. Antioxidative Parameters in the Seedlings of Pigeon Pea (Cajanus cajan L. Millspaugh) in Response to Zn and Ni Stresses. Plant Science, (2000); 157: 113-128.
- Bradford MM. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Analytical Biochemistry, (1976); 72: 248-254.
- Laemmli UK. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature, (1970); 227(5259): 680-685.
- Beauchamp C, Fridovich I. Superoxide Dismutase: Improved Assays and Applicable to Acrylamide Gels. Analytical Biochemistry, (1971); 44: 276-287.
- Vitória AP, Lea PJ, Azevedo RA. Antioxidant Enzymes Responses to Cadmium in Radish Tissues. Phytochemistry, (2001); 57(5): 701-710.
- Nakano Y, Asada K. Hydrogen Peroxide is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant and Cell Physiology (1981); 22(3): 867-880.
- Herzog V, Fahimi HD. A New Sensitive Colorimetric Assay for Peroxidase Using 3, 3'-Diaminobenzidine as Hydrogen Donor. Analytical Biochemistry, (1973); 55: 554.
- Bernt E, Bergmeyer HU. Inorganic Peroxidases. In: Bergmeyer HU, editor: Methods of Enzymatic Analysis. (1974); Vol. 4: 2246–2248. Academic Press, New York, USA.
- Xu S, Li J, Zhang X, Wei H, Cui L. Effects of Heat Acclimation Pretreatment on Changes of Membrane Lipid Peroxidation, Antioxidant Metabolites, and Ultrastructure of Chloroplasts in Two Cool-Season Turfgrass Species under Heat Stress. Environmental and Experimental Botany, (2006); 56: 274–285.
- Farooq M, Wahid A, Kobayashi NSMA, Fujita DBSMA, Basra SMA. Plant Drought Stress: Effects, Mechanisms, and Management: Sustainable Agriculture. (2009); 153-188. Springer, Dordrecht.
- Wrona AF. Factors Affecting Water Use: Newsletter of the Cotton Physiology Education Program. (2000); Vol: 1 No: 1. National Cotton Council.
- Ludlow MM, Muchow RC. A Critical Evaluation of Traits for Improving Crop Yields in Water-Limited Environments. Advances in Agronomy, (1990); 43: 107-153.
- Shen J, Guo M, Wang Y, Yuan X, Wen Y, Song X, Dong S, Guo P. Humic Acid Improves the Physiological and Photosynthetic Characteristics of Millet Seedlings under Drought Stress. Plant Signaling and Behavior, (2020); 15(8): e1774212 (13 pages).
- Michel BE, Kaufmann MR. The Osmotic Potential of Polyethylene Glycol 6000. Plant Physiology, (1973); 51 (5): 914–916.
- Carter JE Jr, Patterson RP. Use of Relative Water Content as a Selection Tool for Drought Tolerance in Soybean: 1985 Agronomy Abstract. (1985); ASA. Madison, WI.
- Nepomuceno A, Oosterhuis D, Stewart J. Physiological Responses of Cotton Leaves and Roots to Water Deficit Induced by Polyethylene Glycol. Environmental and Experimental Botany, (1998); 40(1), 29-41.
- Mansour MMF. Nitrogen-Containing Compounds and Adaptation of Plants to Salinity Stress. Biologia Plantarum, (2000); 43: 491-500.
- Farouk S, Al-Huqail AA. Sustainable Biochar and/or Melatonin Improve Salinity Tolerance in Borage Plants by Modulating Osmotic Adjustment, Antioxidants, and Ion Homeostasis. Plants, (2022); 11(6): 765.
- Samaras Y, Bressan RA, Csonca LN, Garcia-Rios MG, Paino D, Urzo M, Rhodes D. Proline Accumulation During Drought and Salinity. In: Smirnoff N, editor: Environment and Plant Metabolism. (1995); 161-187. Bios Scientific Publishers, Oxford, UK.
- Gomes FP, Oliva MA, Mielke MS, Almeida AAF, Aquino LA. Osmotic Adjustment, Proline Accumulation and Cell Membrane Stability in Leaves of Cocos nucifera Submitted to Drought Stress. Scientia Horticulturae, (2010); 126: 379-384.
- Yildiztugay E., Ozfidan-Konakci C., Kucukoduk M. Exogenous Nitric Oxide (as Sodium Nitroprusside) Ameliorates Polyethylene Glycol-Induced Osmotic Stress in Hydroponically Grown Maize Roots. Journal of Plant Growth Regulation, (2014); 33: 683-696.
- Baloğlu MC, Kavas M, Aydin G, Öktem HA, Yücel AM. Antioxidative and Physiological Responses of Two Sunflower (Helianthus Annuus) Cultivars under PEG-Mediated Drought Stress. Turkish Journal of Botany, (2012); 36: 707-714.
- Gedeon S, Ioannou A, Balestrini R, Fotopoulos V, Antoniou C. Application of Biostimulants in Tomato Plants (Solanum lycopersicum) to Enhance Plant Growth and Salt Stress Tolerance. Plants, (2022); 11: 3082.
- Wang Y, Branicky R, Noe A, Hekimi S. Superoxide Dismutases: Dual Roles in Controlling ROS Damage and Regulating ROS Signaling. Journal of Cell Biology, (2018); 217(6): 1915–1928.
- Naboulsi I, Ben Mrid R, Ennoury A, Zouaoui Z, Nhiri M, Ben Bakrim W, Yasri A, Aboulmouhajir A. Crataegus oxyacantha Extract as a Biostimulant to Enhance Tolerance to Salinity in Tomato Plants. Plants, (2022); 11(10): 1283.
- Abou-Sreea AIB, Azzam CR, Al-Taweel SK, Abdel-Aziz RM, Belal HEE, Rady MM, Abdel-Kader AAS, Majrashi A, Khaled KAM. Natural Biostimulant Attenuates Salinity Stress Effects in Chili Pepper by Remodeling Antioxidant, Ion, and Phytohormone Balances, and Augments Gene Expression. Plants (Basel), (2021); 10(11): 2316. PMID: 34834678; PMCID: PMC8617650.
- Abdalla MM. Boosting the Growth of Rocket Plants in Response to the Application of Moringa oleifera Extracts as a Biostimulant. Life Sciences, 11: (2014); 1097-8135. http://www.lifesciencesite.com. 189.
- Demiral T, Turkan I. Does Exogenous Glycinebetaine Affect Antioxidative System of Rice Seedlings under NaCl Treatment? Journal of Plant Physiology, (2004); 161: 1089–1100.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0