

Full Length Research Article
Identification of phytochemicals as potential inhibitors of Candida albicans SAP3: An in-silico approach
Zain Alhindi
Adv. life sci., vol. 11, no. 3, pp. 679-683, August 2024
*– Corresponding Author: Zain Alhindi (zhhindi@uqu.edu.sa)
Authors' Affiliations
[Date Received: 05/03/2024; Date Revised: 01/06/2024; Date Published: 10/07/2024]
Editorial Expression of Concern:
Editorial Note:
28 May 2025: Reference to the above 'editorial expression of concern', this manuscript has been reanalysed through Turnitin for both similarity index and AI generated content. Similarity index was found well within permissible limit along with AI content having 'higher likelihood of false positive'. Therefore, the concern has been revoked.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Candida albicans is a major threat to human health, causing a variety of diseases. Candida albicans can overgrow and trigger infections like thrush, diaper rash, and vaginal yeast infections. Therefore, finding potential inhibitors of C. albicans from plant sources has become important, recently.
Methods: This study investigated the interaction of natural compounds (N = 450) from the ZINC database with the C. albicans SAP3 protein. PyRx tools were used to conduct molecular docking analyses, which included ligand and target receptor preparation, data analysis, and visualization.
Result: The binding energies between the natural compounds and SAP3 ranged from -8.8 to -7.8 kcal/mol, indicating that the interactions were predominantly strong. Among the compounds screened, ZINC000252509014, ZINC000252509722, ZINC000253389136, ZINC000251981944, and ZINC000252480871 were extensively explored in this study because they were found to be the best among others. Furthermore, these compounds showed favorable drug-like properties.
Conclusion: These findings lay the groundwork for future research aimed at developing potent SAP3 inhibitors to treat C. albicans infections.
Keywords: Candida albicans; SAP3 protein; Natural compounds; Virtual screening; Drug-likeness
Introduction![]()
Fungal infections kill an estimated 1.6 million people each year [1]. Candida albicans, a fungal pathogen, causes around 150 million mucosal infections and nearly 200,000 deaths each year, largely through invasive and disseminated illnesses in vulnerable populations. Candida infections have an economic impact of around $2 billion in healthcare expenditures in the United States each year [2], with comparable per capita costs found in the European Union. C. albicans is responsible for approximately 75% of all Candida infections, making it a huge worldwide health concern whose severity is increasing.
C. albicans frequently colonizes humans, with estimates indicating that between 20% and 80% of the global population is asymptomatically colonized at any given moment [3,4], albeit this incidence varies depending on host health state and dietary choices [5,6]. Colonization can occur at a variety of anatomical locations, including the mouth cavity, skin, gastrointestinal system, and vaginal mucosa [7,8]. These locales expose colonizing fungus to a variety of physiological challenges, including variations in pH, temperature, and oxygen levels, as well as nutritional scarcity and host immunological responses [9-11]. Although C. albicans' interactions with its human host are mostly commensal, the fungus can transform into an opportunistic pathogen, causing an estimated 400,000 serious bloodstream infections each year [12-14]. C. albicans can also cause less severe mucosal infections, including oral and vaginal thrush, as well as cutaneous infections [1]. Hence, C. albicans is the second most common fungal pathogen in people and the leading cause of healthcare-associated fungal infections [15].
A variety of factors and behaviors have been linked to C. albicans pathogenicity [16,17]. Secreted aspartic proteinases (SAP) 1-3 have been identified as key players in adhesion and tissue damage during local infections [18,19]. A full understanding of the SAP family's participation in C. albicans pathogenicity has been established by determining the involvement of distinct SAP variants in mucosal vs systemic infections; specifically, SAP 1-3 in mucosal infections and SAP 4-6 in systemic infections [18]. The diverse role of C. albicans SAP in the infection process highlights the potential of blocking SAP as a treatment method for a wide range of pathogen-caused illnesses [20].
Despite advances in basic life sciences and biotechnology, drug discovery and development still take 15 years and costs US$2 billion for a small-molecule drug. Clinical trials are the most expensive part of drug development, but discovery and preclinical research offer the most time and cost savings [21]. Preclinical research accounts for over 43% of pharmaceutical costs, with significant public funding, due to high attrition rates from target identification to lead optimization to clinical candidate selection. The 90% clinical trial failure rate is mostly due to early discovery challenges like target validation and ligand characteristics [22]. During the early stages of drug development, finding faster and more accessible methods to find more diverse arrays of high-quality chemical probes, hits, and leads with optimal ADMET profiles and pharmacokinetics properties would improve preclinical and clinical outcomes and help develop more efficacious, accessible, and safer therapies [21]. Increasing costs and attrition in conventional drug discovery and development have highlighted the need for computer-aided drug discovery methods [23]. This study aimed to find natural SAP3 inhibitor to combat the C. albicans infections.
Methods![]()
Protein preparation
The crystal structure of SAP3, indicated by the PDB ID: 2H6T, was obtained from the Protein Data Bank. The structure was cleaned in Discovery Studio Visualizer by removing molecules of pepstatin A, water molecules, and Zn. The cleaned and processed protein structure was subsequently saved in the .pdb format for further analysis.
Natural compound library preparation
A collection of 450 natural substances was compiled for this study, which were obtained from the ZINC database. The compounds underwent energy minimization utilizing the Universal Force Field (UFF) in the PyRx program. The compounds were converted to the pdbqt format after minimization to aid in further docking studies.
Virtual screening
Over the past ten years, docking—the main computational approach used in virtual screening (VS) has been the subject of intensive research and widespread use in the drug discovery industry [24]. To investigate the possibility of ligand binding in SAP3’s catalytic pocket, molecular docking studies were carried out with the PyRx0.8 tool [25]. These analyses were presented in 2D and 3D formats to visualize and examine the ligand-protein complex interaction profile. The pepstatin has used as the positive control.
Physicochemical, pharmacokinetics, and drug likeness properties estimation
The SwissADME tool was utilized to assess the physicochemical characteristics, pharmacokinetics, and drug-likeness of the five intriguing compounds. This web-based application allowed for a thorough assessment of each compound's feasibility for future drug development by analyzing significant variables crucial to their potential effectiveness and safety as therapeutic substances.
Results![]()
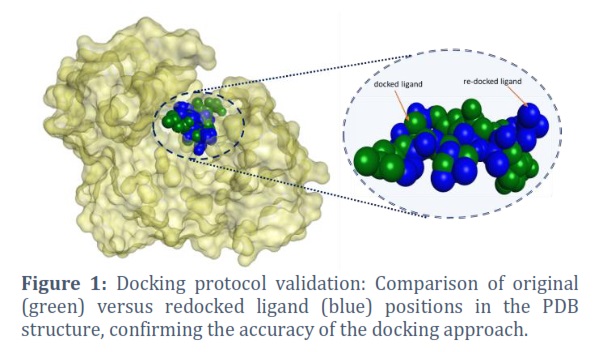
The study aimed to find natural inhibitors of C. albicans SAP3 by using computational screening to analyze 450 compounds from the ZINC database for potential inhibitory properties. Redocking analysis was conducted on the original PDB structure and ligand to validate the docking protocol. The preciseness of the docking approach was confirmed by comparing the position of the redocked ligand to its initial configuration (Figure 1).
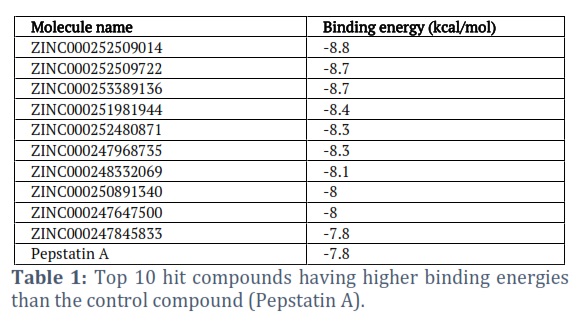
After optimizing the docking protocol, virtual screening was conducted using the PyRx program. The positive control chosen for this study was pepstatin A, which is a co-crystal ligand. The top 10 compounds identified in the screening process are presented in Table 1.
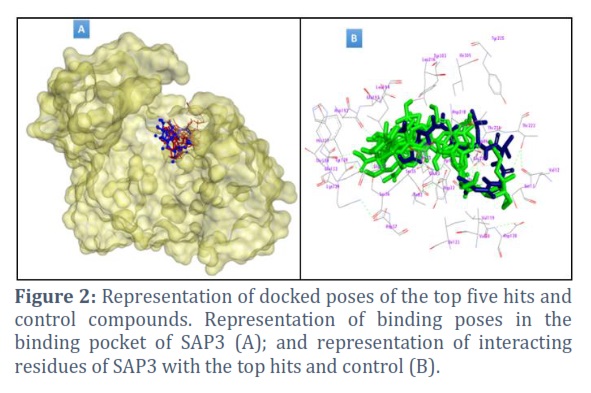
Even though several compounds showed comparable binding energy and interaction, the top five namely ZINC000252509014, ZINC000252509722, ZINC000253389136, ZINC000251981944, and ZINC000252480871 were extensively explored in this study because they were found to be the best among others. Figure 2 illustrates that the top five compounds and the control compound (blue) are binding in the same pocket and share most of the interacting residues of SAP3.
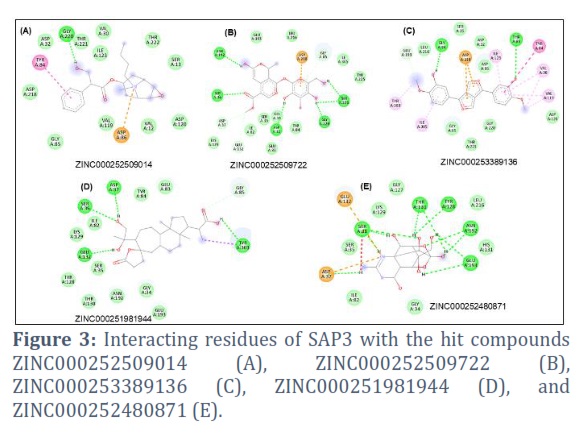
ZINC000252509014 interacted with Tyr84, Asp32, Gly220, Thr221, Val30, Ile123, Thr222, Ser13, Asp120, Val12, Asp86, Val119, Gly85, and Asp218 residues of SAP3. Among them, Asp32, Thr221, Val30, Ile123, Thr222, Ser13, Asp120, Val12, Val119, Gly85, and Asp218 residues make Van der Waals interactions with ZINC000252509014 (Figure 3A). ZINC000252509722 interacted with Asn192, Glu193, Leu216, Asp218, Gly85, Ile305, Tyr225, Thr221, Gly220, Tyr84, Gly34, Asp32, Glu83, Ser35, Ile82, Glu132, Lys129, Asp37, and Ser36 residues of SAP3 (Figure 3B). ZINC000253389136 interacted with Tyr303, Glu193, Gly34, Ser35, Asp218, Asp32, Ile123, Asp86, Thr88, Tyr84, Val30, Val119, Asp120, Gly220, Thr221, Gly85, and Ile305 residues of SAP3 (Figure 3C). ZINC000251981944 interacted with Tyr303, Glu193, Gly34, Asn192, Thr130, Tyr128, Ser35, Glu132, Lys129, Ile82, Ser36, Asp37, Tyr84, Glu83, and Gly85 residues of SAP3 (Figure 3D). Further, ZINC000252480871 interacted with Glu132, Lys129, Gly127, Thr130, Tyr128, Leu216, Asn192, His131, Glu193, Gly34, Ile82, Asp37, Ser35, and Ser36 residues of SAP3 (Figure 3E).
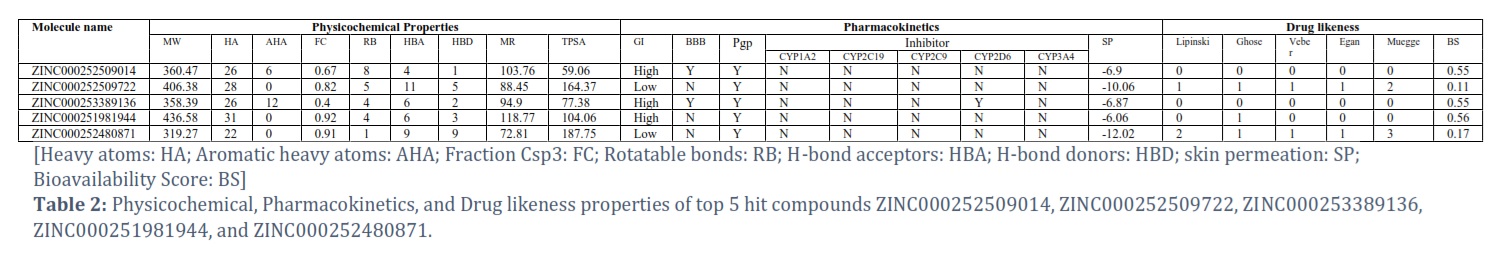
A compound must possess favorable physicochemical, pharmacokinetic, and drug-likeness properties to be deemed a potent drug candidate. In this context, the five chosen compounds were evaluated based on various parameters.
All compounds were found to have favorable properties and met the criteria for potential drug efficacy based on the assessments (Table 2).
Figures & Tables
Currently, the Candida genus has about 150 species, with approximately 20 recognized for their pathogenicity in humans. Candida albicans is the most common cause of candidiasis, a fungal illness that affects both adults and children [26,27]. It has been established that C. albicans contains genes encoding for SAP1 to SAP10 proteins, which are known as classical virulence factors and whose expression is modulated by a variety of factors such as pH, temperature, infection site, and physicochemical environmental conditions [20,28]. This work focused on SAP3, one of the SAP proteins, because it has previously crystallized with pepstatin [18]. The study examined 450 chemicals from the ZINC database for putative SAP3 inhibitory action. Among these compounds, the top five (ZINC000252509014, ZINC000252509722, ZINC000253389136, ZINC000251981944, and ZINC000252480871) were found to have a high affinity for SAP3.
The minimal binding energy obtained from docking simulations can be used to determine the ligand binding efficiency to the target protein [29-32]. When compared to the positive control, pepstatin, the hit compounds (ZINC000252509014, ZINC000252509722, ZINC000253389136, ZINC000251981944, and ZINC000252480871) show lower binding energies, suggesting a robust interaction between the hit compounds and SAP3.
H-bonding has an important role in mediating the ligand-protein interactions [33]. The hit compounds ZINC000252509014, ZINC000252509722, ZINC000252480871, and ZINC000253389136 demonstrated H-bonding interactions with several SAP3 protein residues.
Although antifungal medications with novel mechanisms of action are being developed in clinical trials, their number is rather modest in comparison to therapeutic treatments for other diseases [34]. Identification of further preclinical leads is critical, and natural product screening plays an important part in this effort. Natural products are distinguished by their significant structural variety, which allows for study of chemical regions that would be difficult to access only through synthetic substances [35,36]. Furthermore, natural products have been extensively evaluated for their biological functions, with multiple examples of approval as medicinal agents, either in their native form or as semisynthetic derivatives [37].
This study used the in-silico screening strategy to investigate the potential of compounds as SAP3 inhibitors. ZINC000252509014, ZINC000252509722, ZINC000252480871, and ZINC000253389136 were identified as having a high binding energy to the SAP3 protein. The favorable interactions observed were attributed to Van der Waals and H-bonding interactions. Furthermore, these compounds demonstrated promising drug-like characteristics. These findings lay the groundwork for future experimental research into the development of potent SAP3 inhibitors for C. albicans infections.
The author declare that there is no conflict of interest regarding the publication of this paper.
![]() References
References
- Brown GD, Denning DW, Gow NA, Levitz SM, Netea MG, White TC. Hidden killers: human fungal infections. Science Translational Medicine, (2012); 4(165): 165rv113.
- Benedict K, Jackson BR, Chiller T, Beer KD. Estimation of Direct Healthcare Costs of Fungal Diseases in the United States. Clinical Infectious Diseases, (2019); 68(11): 1791-1797.
- Mukherjee PK, Sendid B, Hoarau G, Colombel JF, Poulain D, Ghannoum MA. Mycobiota in gastrointestinal diseases. Nature Reviews Gastroenterology & Hepatology, (2015); 12(2): 77-87.
- Polvi EJ, Li X, O'Meara TR, Leach MD, Cowen LE. Opportunistic yeast pathogens: reservoirs, virulence mechanisms, and therapeutic strategies. Cellular and Molecular Life Sciences, (2015); 72(12): 2261-2287.
- Li J, Chen D, Yu B, He J, Zheng P, et al. Fungi in Gastrointestinal Tracts of Human and Mice: from Community to Functions. Microbial Ecology, (2018); 75(4): 821-829.
- Gunsalus KT, Tornberg-Belanger SN, Matthan NR, Lichtenstein AH, Kumamoto CA. Manipulation of Host Diet To Reduce Gastrointestinal Colonization by the Opportunistic Pathogen Candida albicans. mSphere, (2016); 1(1): e00020-15.
- Noble SM, Gianetti BA, Witchley JN. Candida albicans cell-type switching and functional plasticity in the mammalian host. Nature Reviews Microbiology, (2017); 15(2): 96-108.
- Limon JJ, Skalski JH, Underhill DM. Commensal Fungi in Health and Disease. Cell Host & Microbe, (2017); 22(2): 156-165.
- Alves R, Barata-Antunes C, Casal M, Brown AJP, Van Dijck P, Paiva S. Adapting to survive: How Candida overcomes host-imposed constraints during human colonization. PLOS Pathogens, (2020); 16(5): e1008478.
- Ene IV, Brunke S, Brown AJ, Hube B. Metabolism in fungal pathogenesis. Cold Spring Harbor Perspectives in Medicine, (2014); 4(12): a019695.
- Polke M, Hube B, Jacobsen ID. Candida survival strategies. Advances in Applied Microbiology, (2015); 91: 139-235.
- Zhai B, Ola M, Rolling T, Tosini NL, Joshowitz S, et al. High-resolution mycobiota analysis reveals dynamic intestinal translocation preceding invasive candidiasis. Nature Medicine, (2020); 26(1): 59-64.
- Yan L, Yang C, Tang J. Disruption of the intestinal mucosal barrier in Candida albicans infections. Microbiological Research, (2013); 168(7): 389-395.
- Alenazy H, Alghamdi A, Pinto R, Daneman N. Candida colonization as a predictor of invasive candidiasis in non-neutropenic ICU patients with sepsis: A systematic review and meta-analysis. International Journal of Infectious Diseases, (2021); 102: 357-362.
- Magill SS, O'Leary E, Janelle SJ, Thompson DL, Dumyati G, et al. Changes in Prevalence of Health Care-Associated Infections in U.S. Hospitals. The New England Journal of Medicine, (2018); 379(18): 1732-1744.
- Kumamoto CA, Gresnigt MS, Hube B. The gut, the bad and the harmless: Candida albicans as a commensal and opportunistic pathogen in the intestine. Current Opinion in Microbiology, (2020); 56: 7-15.
- Prieto D, Correia I, Pla J, Roman E. Adaptation of Candida albicans to commensalism in the gut. Future Microbiology, (2016); 11(4): 567-583.
- Borelli C, Ruge E, Schaller M, Monod M, Korting HC, et al. The crystal structure of the secreted aspartic proteinase 3 from Candida albicans and its complex with pepstatin A. Proteins, (2007); 68(3): 738-748.
- Calugi C, Guarna A, Trabocchi A. Insight into the structural similarity between HIV protease and secreted aspartic protease-2 and binding mode analysis of HIV-Candida albicans inhibitors. Journal of Enzyme Inhibition and Medicinal Chemistry, (2013); 28(5): 936-943.
- Santos ALS, Braga-Silva LA, Goncalves DS, Ramos LS, Oliveira SSC, et al. Repositioning Lopinavir, an HIV Protease Inhibitor, as a Promising Antifungal Drug: Lessons Learned from Candida albicans-In Silico, In Vitro and In Vivo Approaches. Journal of Fungi (Basel), (2021); 7(6): 424.
- Sadybekov AV, Katritch V. Computational approaches streamlining drug discovery. Nature, (2023); 616(7958): 673-685.
- Sun D, Gao W, Hu H, Zhou S. Why 90% of clinical drug development fails and how to improve it? Acta Pharmaceutica Sinica B, (2022); 12(7): 3049-3062.
- Schaduangrat N, Lampa S, Simeon S, Gleeson MP, Spjuth O, Nantasenamat C. Towards reproducible computational drug discovery. Journal of Cheminformatics, (2020); 12(1): 9.
- Kontoyianni M. Docking and Virtual Screening in Drug Discovery. Methods in Molecular Biology, (2017); 1647: 255-266.
- Dallakyan S, Olson AJ. Small-molecule library screening by docking with PyRx. Methods in Molecular Biology, (2015); 1263: 243-250.
- Pappas PG, Lionakis MS, Arendrup MC, Ostrosky-Zeichner L, Kullberg BJ. Invasive candidiasis. Nature Reviews Disease Primers, (2018); 4: 18026.
- Sardi JCO, Scorzoni L, Bernardi T, Fusco-Almeida AM, Mendes Giannini MJS. Candida species: current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options. Journal of Medical Microbiology, (2013); 62(Pt 1): 10-24.
- Naglik JR, Challacombe SJ, Hube B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbiology and Molecular Biology Reviews, (2003); 67(3): 400-428.
- Sayed Murad HA, M MR, Alqahtani SM, B SR, Alghamdi S, et al. Molecular docking analysis of AGTR1 antagonists. Bioinformation, (2023); 19(3): 284-289.
- Kamal MA, H MB, I JH, R SA, M SH, et al. Insights from the molecular docking analysis of EGFR antagonists. Bioinformation, (2023); 19(3): 260-265.
- Elaimi A, Hanadi MB, Almutairi A, Alniwaider RA, Abulkaliq MA, et al. Insights from the molecular docking analysis of GRP78 with natural compound inhibitors in the management of cancers. Bioinformation, (2023); 19(1): 39-42.
- I JH, Alsharif FH, Aljadani M, Fahad Alabbas I, Faqihi MS, et al. Molecular docking analysis of KRAS inhibitors for cancer management. Bioinformation, (2023); 19(4): 411-416.
- Chen D, Oezguen N, Urvil P, Ferguson C, Dann SM, Savidge TC. Regulation of protein-ligand binding affinity by hydrogen bond pairing. Science Advances, (2016); 2(3): e1501240.
- Perfect JR. The antifungal pipeline: a reality check. Nature Reviews Drug Discovery, (2017); 16(9): 603-616.
- Ortholand JY, Ganesan A. Natural products and combinatorial chemistry: back to the future. Current Opinion in Chemical Biology, (2004); 8(3): 271-280.
- Ganesan A. The impact of natural products upon modern drug discovery. Current Opinion in Chemical Biology, (2008); 12(3): 306-317.
- Newman DJ, Cragg GM. Natural Products as Sources of New Drugs from 1981 to 2014. Journal of Natural Products, (2016); 79(3): 629-661.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()