

Full Length Research Article
Evaluation of Micro-RNA Expression Profiling Level as Biomarkers for Diagnosis and Gene Sequencing in Patients Suffering from Breast Cancer
Rasha H. Al-Rikabi1, Nagui H. Fares2, Mahmmad A. AL Faham3, Abdel Hady A. Abdel Wahab4
Adv. life sci., vol. 11, no. 3, pp. 641-647, August 2024
*– Corresponding Author: Rasha H. Al-Rikabi (rashahabeeb2021@gmail.com)
Authors' Affiliations
2. Cell Biology and Histology Faculty of Science, Ain Shams University – Egypt
3. Microbiology and Molecular Biology Faculty of medicine, Baghdad University – Iraq
4. Medical Biochemistry and Molecular Biology National cancer institute, Cairo University – Egypt
[Date Received: 13/11/2023; Date Revised: 21/06/2024; Date Published: 10/07/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Human bloodstream microRNAs (miRNAs) have emerged as a promising predictive and diagnostic biomarker for a range of cancers, including breast cancer. Our objective was to look into new miRNA biomarkers for diagnosis in the serum of patients with breast cancer and track the expression levels at different stages using miRNA profiling.
Methods: 53 breast cancer patients and 10 healthy controls had blood samples tested for three miRNAs. miRNAs were extracted from blood and evaluated using real-time quantitative polymerase chain reaction. After extracting genomic DNA, miRNA primer-produced PCR products were sequenced to discover point mutations that may contribute to the illness.
Results: After examining miR195, miR200b, and miR331, breast cancer patients had a significantly lower miR195 level than healthy persons. In addition, miR200b expression levels were significantly lower in breast cancer patients than in healthy individuals. In advanced stages, miR331 expression was substantially higher than in healthy people.
Conclusion: The findings of our study demonstrated a significant association between the expression of miRNAs and the prognosis of breast cancer. Additional research is necessary to study better the correlation between these circulating miRNAs and the stages of breast cancer.
Keywords: Breast cancer; Circulating miRNAs; miR195; miR200b; miR331 RT PCR; Gene Sequencing
Introduction![]()
Finding reliable and precise minimally invasive biomarkers that may identify early neoplastic alterations is a continuing search in breast cancer management. These biomarkers are crucial for early-stage breast cancer identification, disease progression tracking, and therapy efficacy assessment [1]. The current breast cancer detection tools and biomarkers have several serious shortcomings. The ionizing radiation it uses and the 8–10% false positive rate are two of the problems with the mammography apparatus, despite its status as the diagnostic gold standard [2].
Breast cancer therapy uses carcinoembryonic and carbohydrate antigens from circulating malignancies. Due to limited sensitivity, these markers are ineffective screening tools. Therapy has long employed markers as prognostic indicators to evaluate illness progression or relapse. An ideal biomarker would be widely accessible, collected with minimum interference, sensitive enough to identify early tumors in almost all patients, and uncommon or nonexistent in healthy persons without malignancies [3,4].
Hereditary mutations in the BRCA1 and BRCA2 genes cause 5–10% of breast neoplasms, according to Feng Y [5]. Additionally, gene expression profiling has shown the dysregulation of numerous genes in breast cancer samples and confirmed its genetic variability [5]. Several studies have shown microRNA dysregulation in breast cancer samples or plasma samples related to carcinogenesis. Two RNase III proteins—Drosha and Dicer—create miRNAs in many steps [6]. Numerous human gene expression levels are regulated by these tiny non-coding transcripts, which are primarily responsible for post-transcriptional regulation [7]. They have also been found to impact the onset of human disorders largely. On the other hand, miRNA metabolism and activities are regulated by sophisticated mechanisms [7].
This kind of control defines the micro-RNA expression in various tissues or at various developmental stages. Most significantly, miRNAs have a role in autoregulatory feedback pathways by suppressing the production of miRNAs that code for components involved in miRNA formation [7]. Some breast cancer samples show altered miRNA expression. Current study shows aberrant miR221 and miR222 expression in breast cancer [8].
Methods![]()
Cohort of research: This research was conducted with a group of 53 breast cancer patients. Specialists made the diagnoses for all patients, and we have access to their histopathology reports and immunohistochemistry data, which include markers for ER, PR, and Her2/neu. Participants' ages ranged from thirty to seventy-four, and all ten were healthy individuals free of breast cancer. This research took place at the Oncology Teaching Hospital in Medical City, Baghdad, Iraq (Record No. 51892 on July 12, 2022). All cases of breast cancer were histologically confirmed, and relevant clinicopathological characteristics were extracted from their medical records.
Blood collection: Blood samples were collected from patients and were subjected to miR195, miR200b, and miR331 gene analysis. The 2 ml of blood sample was right away kept at -80°C pending additional examination. An aliquot of blood samples was used to extract the serum, which was then chilled to -80°C for further examination.
DNA extraction from blood: Isolated DNA from whole blood using ReliaPrep™ Blood gDNA Miniprep System (Promega). Quantus Fluorometer purity and 260/280nm ratio estimation of isolated DNA.
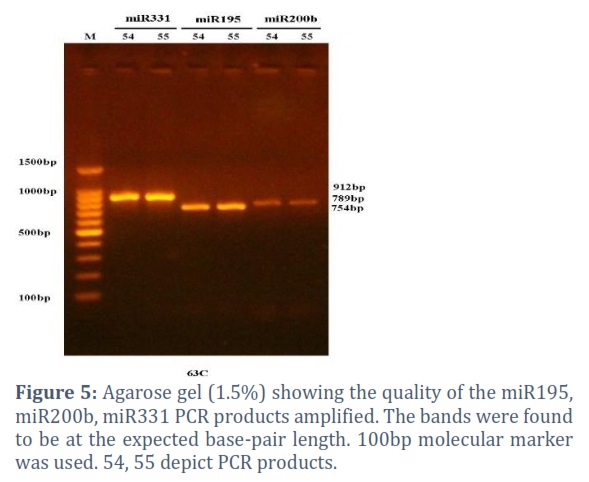
PCR: The Thermal Cycler from BioRad, USA, PCR Express, and the correct temperature program to cycle the PCR: treated with a denaturing heat of 94 °C for four minutes, then subjected to thirteen cycles of denaturation, annealing, and extension at temperatures of 55, 58, 60, 63, or 65 °C for 30 seconds each. In order to terminate the reaction, a last extended incubation of seven minutes at 72 °C was used, and then ten minutes of incubation at 4°C. Macrogen Corporation – Korea utilized an automated DNA sequencer, the ABI3730XL, to run Sanger sequencing on the purified PCR products after the 1.5arose gel was used to detect the product's quality. After receiving the findings by email, generous software was utilized to analyse them. Primers used in the PCR were miR195-F, miR195-R, miR200b-F, miR200b-R, miR331-F, and miR331-R. The primers used were 100pmol/µl.
RNA isolation: Following the instructions in the manual, total RNA was extracted from 0.4 ml of serum using TRIzol™ Reagent (Invitrogen, Carlsbad, CA, USA).
Primers design: The miRNA construct Tool, as suggested by [9], was used to build appropriate stem-loop RT-qPCR primers, and matured micro-RNA sequences were obtained from the miRNA database website (https://www.mirbase.org). This study made use of the oligonucleotides described in Table 1. We used miR195, miR200b, and miR331 micro-specific primers to create our real-time qRT-PCR studies. You may get these primers in lyophilized form from the Macrogen Company.
Differential expression
We contrasted two groups (cases and healthy persons as controls) separated by race and ethnicity for each of the 53 individuals. In a nutshell, a model of linear was fitted to the data for each comparison (cell means representing the various circumstances and a random effect for the array) [10]. We first retrieved the list of microRNAs that were differentially expressed for each comparison with a P<0.05, and we next looked for candidates that had at least a two-fold change in expression.
Sequencing analysis
In order to analyze PCR results, Applied Biosystems used a 16-capillary automated sequencer (ABI PRISM®3100 Genetic Analyzer).
Statistical analysis
We used Graph-Pad Prism Version 8 for our statistical research, which allowed us to correlate several factors. P < 0.05 indicates a statistically significant difference. micro RNAs have shown their worth as a diagnostic tool via the use of a Receiver Operating Characteristic Curve (ROC) study.
Results![]()
qRT-PCR of differentially expressed miRNAs:
Patients with breast cancer had lower circulating blood levels of miR195 compared to healthy controls. More advanced stages (III, IV) "showed a significant association with them (p=0.045, 0,009, respectively). Figure 1. Furthermore, serum miRNA expression levels were significantly higher in early-stage breast cancer patients compared to those in stages III and IV (p=0.0157 and p=0.0830, respectively)".
The findings also showed "that compared to the control group, patients in stage I–IV of breast cancer had significantly reduced miR200b expression (p = 0.0196, 0.049, 0.0081, and 0.005, respectively). Figure 2. Despite an increase in 331 expression data in stages III and IV, the status of breast cancer patients was two times greater than that of the healthy control group (p= 0.0031 and 0.0013, respectively). Fig 3.
Value of the miRNAs in breast cancer diagnosis
With the use of ROC curve analysis, we were able to determine the likelihood that miR-195, miR-331, and miR-200b might identify breast cancer in serum samples from patients. This study demonstrated the discriminating volume "between the breast cancer and healthy control samples by comparing the area under the curve (AUC) obtained by constructing the ROC curve using binary logistic regression."
The ROC curves of miRNAs indicated that miR-195 "had an area under the curve of 0.885, 87.4% sensitivity, and 90.0% specificity. While miR-200b AUC was 0.853, sensitivity was 88.7%, and specificity was 70.0%. For miR-331, AUC was 0.892, sensitivity was 84.9%, and specificity was 98.3%".
DNA isolation
The PCR products, as visualized on the 1.5% agarose gel, were at the expected band length. The band lengths for miR331, miR195, and miR20 b were 912, 789, and 754 bp, respectively.
We found that among the samples sequenced, at 586 positions, SNP was detected (p<0.05). In the samples 54 miR331 and 55 miR331 only we found T got substituted to G.
Highlights of the study
- Four downregulated and upregulated micro RNAs were recognized in the serum of blood in breast cancer patients.
- A point mutation was seen at 586 positions (T-G) in the miR331 samples (both 54 and 55) only.
Figures & Tables
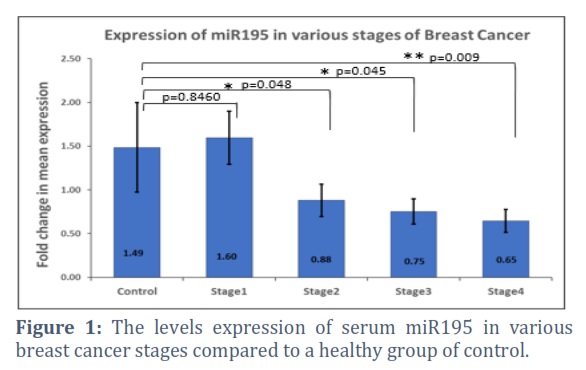
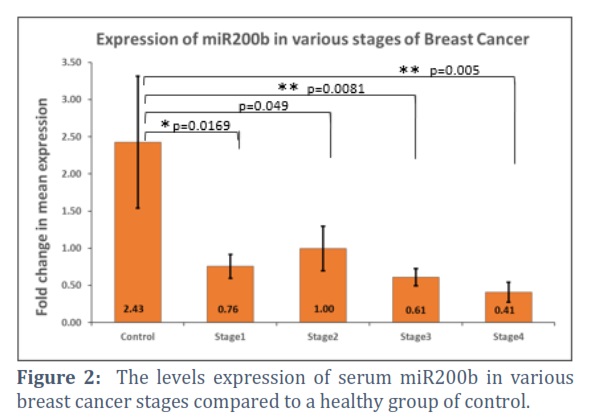
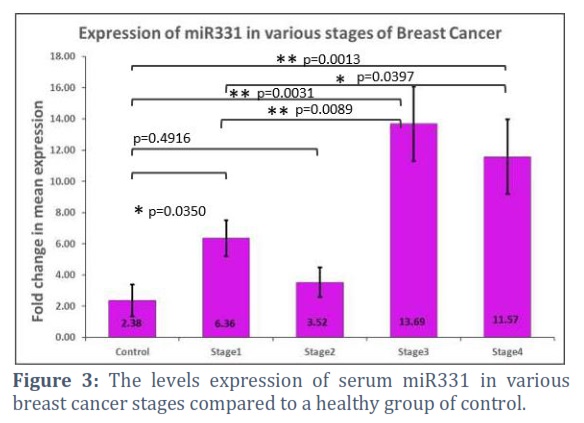

Diagnostic and prognostic indications for breast cancer include aberrant microRNA expression in early-stage and metastatic tumors [11]. Many studies connect miRNA expressions to tumor stage, subtype, and treatment effectiveness [12]. Micro RNA expression levels have linked breast cancer to cancer antigen (CA-125) and carcinoembryonic antigen (CEA) [15]. Much research has examined metastatic breast cancer’s up- or down-regulated miRNA expression to understand better its genesis, progression, and maintenance [13,14]. By targeting several genes, elevated micro RNAs may control a network of functional genes that contribute to tumor formation or metastasis [15].
This research examines whether qRT-PCR can differentiate between healthy controls and early-stage breast cancer patients by evaluating blood levels of cmiR-195, miR-200b, and miR-331. Advanced breast cancer patients had lower serum miR195 levels than healthy controls. These results supported a prior study [16] that found miRNA195 downregulated in breast cancer tissues and cell lines [17]. In another research, breast cancer patients exhibited lower miR-195 levels than healthy controls [18]. A study discovered that breast cancer patients had considerably lower miR195 and miR497 expression and that the same mechanism of CpG methylation upstream of both miR promoters influenced these levels [19, 20]. Such an expression pattern may make it easy to distinguish malignant tumors from benign or normal-state tumors [21]. There is an interacting relationship between tumor size and stage and miR195 levels in the blood, as stated in [22]. Nevertheless, Luo [25] showed that in plasma samples from breast cancer patients, miR195 expression was lower in those with a greater differentiation grade, later clinical stage, and lymph node metastases.
According to Shao et al. [23], 40 breast cancer samples had considerably lower miR-195-5p expression than nearby normal breast tissues. Overexpression of miR-195-5p inhibits cell proliferation and prolongs the G1 phase. [24] shows miR-195p suppresses breast cancer. They showed that miR195 might limit breast cancer cell proliferation and colonization in vitro by targeting CCNE1. This cyclin gene family member governs the cell cycle transition from the G1 to S phase and is an oncogene in various patient malignancies. However, miR-195 expression reduces cell proliferation and invasion in R-75-30 and MCF7 human breast cancer cells via targeting Raf-1 and Ccnd1. These findings suggested that serum miR-195 may be a potential breast cancer therapy target. In addition to breast cancer, miR-195 fights other cancers [25].
In this study, breast cancer patients had considerably lower miR-200b expression than controls. Researchers linked low miR200b expression to aggressive and progressed breast cancer. However, high miRNA-200b levels improved prognosis. Similar results link miR-200b reduction in breast cancer cell lines and tissues to advanced clinical stage and distant metastases [26]. They found a negative link between low miR200b expression and worse outcomes in breast cancer patients, indicating it suppresses tumor growth. miR-200b controls the epithelial-mesenchymal transition, which initiation of metastasis. Researchers in another investigation [27] discovered that transfecting MCF-7 cells with an inhibitor of miR200b significantly reduced miR200b levels, increased ERM expression, and improved cell motility and invasion. Cancers of the breast cancer [28], prostate [29], and non-small cell lung [31] have all been associated with miR200b, a tumor suppressor that regulates the epithelial-mesenchymal transition [30]. Researchers discovered reduced miR-200b levels in renal cell cancer patients [31]. Cancer miR200b dysregulation may result from epigenetic alterations such as DNA methylation, histone modifications, or transcriptional suppression [32,33]. Since it regulates epithelial-mesenchymal transition, the miR-200 family contributes to cancer formation. Since miR-200b is an established member of the miR-200 family, many researchers are interested in its function in cancer treatment and carcinogenesis [34].
This research found that breast cancer patients' blood samples had higher miR331 levels than healthy controls. In breast cancer patients, miR331 expression correlated with TNM stage and lymph node metastases. Previous studies have demonstrated that malignant breast tumors overexpress miR331. This and previous studies imply that miR331 levels may differentiate benign from malignant breast tumors [35]. Increased miR331 expression in hepatocellular carcinoma is related to worse clinicopathological features and worse survival [36]. miR331 interacts with the PH domain and leucine-rich repeat protein phosphatase to activate AKT, which promotes mesenchymal-to-epithelial transition, proliferation, and metastasis. An anti-miR331 vector in a xenograft mouse model greatly reduces proliferation and metastasis, making it a tumor-promoting miRNA [37]. We detected dysregulated miR331 in many cancers. In pancreatic and cervical malignancies, miR331 levels are greater [38,39]. CRC and ovarian cancers also downregulate miR-331 [40,41]. Due to elevated miR331 expression, acute myeloid leukemia patients had poor treatment response and survival rates [42].
Changes in the molecules' expression in bodily fluids due to illness or tissue injury can make them useful biomarkers for cancer diagnosis [43]. The onset of cancer is possible in the event of any dysregulation of miRNA expression. Only a subset of miRNAs linked to cancer should be worried about breast cancer, according to studies [44,45]. While reducing their levels in tumor tissue or cells that generate them, theoretically, the release of selective miRNA into the circulation can increase their levels in blood derivatives like plasma or serum [46]. Relationships between tissue expression and miRNAs in the blood that do not originate in cells remain murky. Scientists speculate that changes at the extracellular level might be due to environmental factors, non-specific effects of neoplasm formation, or changes in tumor cell expression [47]. Some have proposed that microvesicles or exosomes could be a means by which cells secrete these substances [48,49].
On the other hand, miRNAs in circulation are usually the leftovers from cells that have died or are no longer functional, and they are most often seen in plasma outside of exosomes and microvesicles [50,51]. Some miRNAs in circulation may have an immunological response to the tumor as their source or even come from blood cells [52]. Although several studies have shown that cancer alters miRNA levels in the blood, the exact mechanism by which tumor-related miRNAs reach the circulation remains unknown. By the way, [52] presented two theories: tumor miRNAs enter the circulation by tumor cell lysis or through newly formed blood arteries from the tumor microenvironment.
New studies show that miRNA patterns may detect breast cancer early. Thus, our pilot investigation suggests that blood miRNA expression levels may be diagnostic and predictive in early breast cancer patients.
Author Contributions
All authors contributed equally to designing, executing, analyses and drafting of this study.
The author declare that there is no conflict of interest regarding the publication of this paper.
![]() References
References
- Thompson A, Brennan K, Cox A, Harcourt D. Harris A, et al. Evaluation of the current knowledge limitations in breast cancer research: a gap analysis. Breast Cancer Research,(2008); 2(12): 348-365.
- Taplin S, Abraham L, Barlow W. Mammography facility characteristics associated with interpretive accuracy of screening mammography. Journal National Cancer Institute, (2008);100(12),876 – 887.
- Uehara M, Kinoshita T, Hojo T. Akashi-Tanaka S, Iwamoto E, et al. Long-term prognostic study of carcinoembryonic antigen (CEA) and carbohydrate antigen 15–3 (CA 15–3) in breast cancer. International Journal Clinical Oncology, (2007);13,447– 451.
- Kinoshita T, Hojo T. Long-term prognostic study of carcinoembryonic antigen (CEA) and carbohydrate antigen 15–3 (CA 15–3) in breast cancer. International Journal Clinical Oncology, (2007);13(9) :447– 451.
- Feng Y, Spezia M, Huang S, Yuan C, Zeng Z,et al. Breast Cancer Development and Progression: Risk Factors, Cancer Stem Cells, Signaling Pathways, Genomics, and Molecular Pathogenesis. Genes Disease (2018); 5(2):77–106.
- Elango R, Alsaleh K, Vishnubalaji R, Manikandan M, Ali A, et al. MicroRNA Expression Profiling on Paired Primary and Lymph Node Metastatic Breast Cancer Revealed Distinct microRNA Profile Associated With LNM. Frontiers in Oncology (2020);19;10:756.
- Treiber T, Treiber N, Meister G. Regulation of microRNA Biogenesis and its Crosstalk With Other Cellular Pathways. Nature Reviews Molecular Cell Biology, (2019); 20(1):5–20.
- Krol J, Loedige I, Filipowicz W. The Widespread Regulation of microRNA Biogenesis, Function and Decay. Nature Reviews Genetics, (2010);11(9):597–610.
- Czimmerer Z, Hulvely J, Simandi Z, Varallyay E, Havelda Z, et al. Versatile Method to Design StemLoop Primer-Based Quantitative PCR Assays for Detecting Small Regulatory RNA Molecules. PLoS One, (2013);8(1), e55168.
- Abak A, Amini S, Estiar MA, Montazeri V, Sakhinia E, et al. Analysis of miRNA-221 Expression Level in Tumors and Marginal Biopsies From Patients With Breast Cancer (Cross-Sectional Observational Study). Clinical Labortory,(2018); 64(1):169–175.
- Smyth GK. Linear models and empirical Bayes methods for assessing differential expression in microarray experiments. Statistical Applications in Genetics and Molecular Biology, (2004); 3: Article 3.
- Bahmanpour Z, Sheervalilou R, Choupani J, Shekari K, Montazeri V, et al. A new insight on serum microRNA expression as novel biomarkers in breast cancer patients. Journal Cell Physiology, (2019);234(11):19199-19211.
- Bartels C, Tsongalis G. MicroRNAs: novel biomarkers for human cancer. Clinical Chemistry, (2009); 55(4):623-631.
- Opstal-van W, Rodenburg W, Pennings J, van Oostrom C, Beijnen J, et al. A bead-based multiplexed immunoassay to evaluate breast cancer biomarkers for early detection in pre-diagnostic serum. International Journal Molecular Sciences,(2012); 13(10):13587-13604.
- Jang K, Ahn H, Sim J, Han H, Abdul R. et al. Loss of microRNA-200a expression correlates with tumor progression in breast cancer. Translational Research,(2014); 163(3), 242-251.
- Hafez M, Hassan Z, Zekri A, Gaber A, Al Rejaie S, et al. MicroRNAs and metastasis-related gene expression in Egyptian breast cancer patients. Asian Pacific Journal of Cancer Prevention,(2012);13(2), 591-598.
- Pencheva N, Tavazoie S. Control of metastatic progression by microRNA regulatory networks. Nature Cell Biology, (2013); 15: 546-554.
- Li D, Zhao Y, Liu C, Chen X, Qi, Y, et al. Analysis of MiR-195 and MiR-497 expression, regulation and role in breast cancer. Clinical Cancer Research,(2011); 17:1722–1730.
- Hannafon B, Sebastiani, P, de las Morenas, A, Lu, J, Rosenberg, C. Expression of microRNA and their gene targets are dysregulated in preinvasive breast cancer. Breast Cancer Research,(2011); 13 (2): R24.
- Zhao F, Dou, Y, Wang X, Han D, Lv Z, et al. Serum microRNA-195 is down-regulated in breast cancer: a potential marker for the diagnosis of breast cancer. Molecular Biology Report, (2014); 41(9):5913-5922.
- Deng H, Guo Y, Song H, Xiao B, Sun W, et al. MicroRNA-195 and microRNA-378 mediate tumor growth suppression by epigenetic regulation in gastric cancer. Gene,(2013); 518 (2);351–359.
- Yu W, Liang X, Li X, Zhang Y, Sun Z, et al. MicroRNA-195: a review of its role in cancers. Oncotargets and therapy,(2018); 11: 7109–7123.
- Shao Y, Sun X, He Y, Liu C, Liu H, et al. Elevated levels of serum tumor markers CEA and CA15-3 are prognostic parameters for different molecular subtypes of breast cancer. PLoS One,(2015); 10 (7):e133830.
- Nadeem F, Hanif M, Ahmed A, Jamal Q, Khan A. Clinicopathological features associated with MiRNA-195 expression in patients with breast cancer: evidence of a potential biomarker. Pakistan Journal of Medical Sciences, (2017); 33 (5), 1242–1247.
- Luo Q, Wei C, Li X, Li J, Chen L, et al. MicroRNA-195-5p is a potential diagnostic and therapeutic target for breast cancer. Oncology Report,(2014); 31 (3), 1096–1102.
- Kumar V, Abbas A.K, Fausto N, Aster J. Robbins, and Cotran Pathologic Basis of Disease Elsevier: health sciences, (2014); 9th ed.
- Ye F, Tang H, Liu Q. miR-200b as a prognostic factor in breast cancer targets multiple members of the RAB family. Journal of Translational Medicine,(2014); 12(17):1-10.
- Hong H, Yu H, Yuan J, Guo C, Cao H, et al. MicroRNA-200b Impacts Breast Cancer Cell Migration and Invasion by Regulating Ezrin-Radixin-Moesin. Medical Sciences Monitor,(2016); 8(22):1946-1952.
- He M, Liu Y, Deng X, Qi S, Sun X, et al . Down-regulation of miR-200b-3p by low p73 contributes to the androgen-independence of prostate cancer cells. Prostate (2013), 73(10):1048–1056.
- Cai Z, Zhang S, Zhang H, Zhou Y, Wu H, et al. Aberrant expression of microRNAs involved in epithelial-mesenchymal transition of HT-29 cell line. Cell Biology International,(2013).; 37(7):669–674.
- Pacurari M, Addison J, Bondalapati N, Wan Y, Luo D, et al. The microRNA-200 family targets multiple non-small cell lung cancer prognostic markers in H1299 cells and BEAS-2B cells. International Journal Oncology, (2013); 43:548–560.
- Yoshino H, Enokida H, Itesako T, Tatarano S, Kinoshita T, et al. Epithelial-mesenchymal transition-related microRNA-200s regulate molecular targets and pathways in renal cell carcinoma. Juornal Human Genet. (2013); 58:508–516.
- Castilla M, Díaz-Martín J, Sarrió D, Romero-Pérez L, López-García M, et al. MicroRNA-200 family modulation in distinct breast cancer phenotypes. PLoS One. (2012);7(10):e47709.
- Davalos V, Moutinho C, Villanueva A, Boque R, Silva P, et al. Dynamic epigenetic regulation of the microRNA-200 family mediates epithelial and mesenchymal transitions in human tumorigenesis. Oncogene, (2011); 31:2062–2074.
- Kang L, Mao J, Tao Y, Song B, Ma W, et al. MicroRNA-34a suppresses the breast cancer stem cell-like characteristics by downregulating Notch1 pathway. Cancer Sciences,(2015); 106:700–708.
- Papadopoulos E., Papachristopoulou G, Ardavanis A, Scorilas A. A comprehensive clinicopathological evaluation of the differential expression of microRNA-331 in breast tumors and its diagnostic significance. Clinical Biochemistry. (2018); 60:24-32.
- Chang R, Yang H, Fang F, Xu J, Yang L. MicroRNA-331-3p promotes proliferation and metastasis of hepatocellular carcinoma by targeting PH domain and leucine-rich repeat protein phosphatase. Hepatology,(2014);60: 1251 –63.
- Chen X, Luo H, Li X, Tian X, Peng B, et al. miR-331-3p functions as an oncogene by targeting ST7L in pancreatic cancer. Carcinogenesis (2018);39(8):1006–1015.
- Fujii T, Shimada K, Asano A, Tatsumi Y, Yamaguchi N, et al. MicroRNA331-3p Suppresses Cervical Cancer Cell Proliferation and E6/E7 Expression by Targeting NRP2. International Journal Molecular Sciences, (2016);17(8):13-51.
- Zhao D, Sui Y, Zheng X. MiR-331-3p inhibits proliferation and promotes apoptosis by targeting HER2 through the PI3K/Akt and ERK1/2 pathways in colorectal cancer. Oncology Report,(2016); 35(2):1075–1082.
- Buranjiang G, Kuerban R, Abuduwanke A, Li X, Kuerban G. MicroRNA-331-3p inhibits proliferation and metastasis of ovarian cancer by targeting RCC2. Archives of Medical Sciences,(2019); 15(6):1520–1529.
- Butrym A, Rybka J, Baczyńska D, Tukiendorf A, Kuliczkowski K, et al. Expression of microRNA-331 can be used as a predictor for response to therapy and survival in acute myeloid leukemia patients. Biomarkers in Medicine,(2018); 9(5):453-460.
- Schwarzenbach H, Nishida N, Calin G, Pantel K. Clinical relevance of circulating cell-free microRNAs in cancer. Nature Reviews Clinical Oncology,(2014);11(3): 145–156.
- Roth C, Rack B, Muller, V. Circulating microRNAs as blood-based markers for patients with primary and metastatic breast cancer. Breast Cancer Research,(2010);12:R90.
- Van Schooneveld E, Wouters M, van der Auwera I, Peeters D, Wildiers H, et al. Expression profiling of cancerous and normal breast tissues identifies microRNAs that are differentially expressed in serum from patients with (metastatic) breast cancer and healthy volunteers, Breast Cancer Research, (2012); 14 (1) R34.
- Pigati L, Yaddanapudi S, Iyengar R. Selective release of microRNA species from normal and malignant mammary epithelial cells. PLoS One,(2010);5: e13515.
- Witwer K. Circulating microRNA biomarker studies: pitfalls and potential solutions. Clinical Chemistry, (2015);61(1):56-63.
- Valadi H, Ekstrom K, Bossios A. Exosomemediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature Cell Biology, (2007); 9:654–659.
- Gallo A, Tandon M, Alevizos, I. The majority of microRNAs detectable in serum and saliva are concentrated in exosomes. PLoS One, (2012);7: e30679.
- Turchinovich A, Weiz L, Burwinkel B. Extracellular miRNAs: the mystery of their origin and function. Trends Biochemistry Sciences (2012); 37: 460–465.
- Arroyo J. Chevillet J. Kroh E. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proceedings of the National Academy of Sciences of the United States of America,(2011);108(12): 5003–5008.
- Pritchard C, Kroh E, Wood B. Blood cell origin of circulating microRNAs: a cautionary note for cancer biomarker studies. Cancer Prevention Research (Phila) (2012); 5(3):492–497.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0