

Full Length Research Article
Extraction and Partial Purification of Glucoamylase from Rhizopus oryzae by Solid State Fermentation using Agro-Industrial Residues
Qays Majeed Issa
Adv. life sci., vol. 11, no. 3, pp. 607-612, August 2024
*- Corresponding Author: Qays Majeed Issa (ojasim79@yahoo.com)
Authors' Affiliations
[Date Received: 10/07/2023; Date Revised: 25/04/2024; Date Published: 10/07/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Filamentous fungi are used to produce many industrial enzymes because these fungi have a great ability to produce such enzymes, especially when using solid substrates that are usually low in cost. The widespread use of this enzyme has made it very important to work on improving its production to achieve the maximum possible benefit.
Methods: Five fungal isolates were obtained from soil and decaying fruits and vegetables. All isolates were identified as Rhizopus spp. and cultured on Potato Dextrose Agar (PDA) plates escorted by starch 1%. The appearance of clearance zones around the fungal colonies represents the ability of fungi to produce Glucoamylase. Different solid substrates, nitrogen sources, and temperatures were used to improve enzyme production.
Results: Wheat bran gave the highest enzyme production with specific activity 6.25 U/mg, and yeast extract was the potent inducer for enzyme production with specific activity 9.28 U/mg. The optimal temperature for enzyme production was 30°C with specific activity10.27U/mg. Maximum specific activity 18.40U/mg was recorded at 55% saturation with ammonium sulfate precipitation. The result showed increased specific activity 24.84U/mg with dialysis. Partial purification of the enzyme revealed that pH5 was optimum for enzyme-specific activity 9.2U/ml; higher enzyme-specific activity was found with MnSO4 (11.83U/ml) as the best enzyme inducer.
Conclusion: This investigation attempted to examine R. oryzae as an effective maker of GA. Innovative work into the nearby creation of GA for industrial use utilizing local resources has demonstrated financial effectiveness. It requires improvement into consistency as needed by worldwide associations dealing with industrial enzymes.
Keywords: Glucoamylase; Production; Partial purification; Rhizopus oryzae; Solid state fermentation
Introduction![]()
Glucoamylase (GA) is an enzyme that is secreted from the outside of living cells. The non-reduced ends of the sequence of glycogen and amylose are used to make it by breaking down the α-1,4 bond with water [1]. This enzyme has many applications in biotechnology, as it is involved in brewing beer, alcoholic beverages, the food industry, and many medical applications. Many microorganisms, such as bacteria and fungi, produce this enzyme. Filamentous fungi(molds), however, are the better source among all microorganisms [2, 3]. In recent years, solid-state fermentation (SSF) procedures have been applied more and more for the production of GA. SSF is a low-cost technology with high productivity per reactor volume and easier procedures. SSF is particularly important in using the raw fermented product directly as an enzyme source [4, 5].
In the present study, process optimization of GA production from R. oryzae was carried out using different agro-industrial residues as substrates.
Methods![]()
Fungal Isolates
Fungal isolates were obtained from soil and decaying fruits and vegetables. Five isolates were identified as Rhizopus sp. according to [6] and propagated on slants of Sabouraud agar medium. Mycelial cultures were incubated at 30°Cfor 5 days, then they were kept in the refrigerator at 4°C until use.
Screening of the Most GA-Producing Strain
A piece of each of the five mold strain colonies of Rhizopus sp. was taken by cork-borer (5mm) and cultured on Potato Dextrose Agar medium (PDA) plates, escorted by starch 1%. All dishes were incubated for 28 hours at a temperature of 30°C; fungal cultures were flooded with iodine reagent. A measuring ruler measured the clearance zones areas around the fungal colonies. The appearance of clearance zones around the fungal colony represents the ability of fungus to produce GA [7,8]. Colonies of fungal strains and a clear zone formed around them were measured in diameter and further defined by the GA Index (GI):
GI = diameter of clear zone(mm) ‒ diameter of colony(mm)
Preparation of Spore Suspension
The isolate was chosen to produce the enzyme, and the largest diameter of the clearance zone appeared. The fungus was cultivated on (PDA). It was incubated for 120 hours at 30°C. After half an hour, the fungal culture was immersed in a sodium chloride solution 0.085% mixed with a solution of Tween 80 (1%), filtered using sterile gauze, where spores were obtained, and the required concentrations were determined using a hemocytometer.
Substrates
Commercially available wheat bran, rice husk, ground nut peel, and corn cobs were desiccated in an electric oven at 60°C even with weight stability and ground and used as catalyst substrates for enzyme production [9].
GA production by SSF
The fermentation was carried out in a 250 ml Erlenmeyer flask containing 10g of wheat bran, rice husk, ground nut peel, and corn cobs as substrate with 1% starch (w/w). Flasks were moistened with 30ml (3:1 V/W) of aqueous mineral salts solution: MgSO4.7H2O (1%); KH2PO4 (0.1%); CaCl2 (0.1%); FeSO4.7H2O (0.05%); and (NH4)2SO4 (0.1%). All flasks were autoclaved at 121°C for 15min. Adding 1 ml of R. oryzae spore suspension (2×107 spores/ml) started the fermentation process. The spores were mixed with the substrates well, then the flasks were incubated at 30°C for 120 hours [10].
Extraction of Crude GA
Fifty mM of citrate buffer solution with pH5 at a ratio of 10:1 was added to the fermented dough. A shaking incubator homogenized the mixture for two hours at 25°C. The mixture was filtered using Whatman filter paper No.1, and a centrifugation process was carried out at a speed of 6000 rpm. for 15 min., the precipitate was discarded, and the filtrate was used as a source of the GA to measure its activity and conduct other studies.
Measure of GA activity
One ml of soluble starch solution 2% was mixed with 0.1 M sodium acetate buffer with pH 5 and 1ml of enzyme solution. The reaction mixture was incubated for 20 min. at a temperature of 40°C. The liberation glucose was calculated with DNSA detector [11] utilizing glucose acting as a standard. GA activity unit is the amount of enzyme liberation one micromole of glucose /min / ml.
Estimate the amount of protein
The samples of protein were calculated by the technique of [12].
Optimization of the production process
Effect of Different Agro-Residues as Carbon Source
Four different solid substrates were used: wheat bran(WB), rice husk(RH), ground nut peel(GNP), and corn cobs(CC), each separately, with 1% starch(w/w). The fermentation was done in a conical flask (250 ml) containing 10 g of a dry solid substrate. The moisture ratio was adjusted to 3:1(V: W) with mineral salts and autoclaved at 121ºC and 15 psi for 20 min. After cooling, each flask was inoculated with 1ml of 2х107 spore/ml and incubated at 30ºC for 120 h.
Effect of Different Nitrogen Sources
Organic nitrogen sources such as (urea, peptone, yeast extract, tryptone, and soya bean meal) were investigated. These sources were added to the solid substrate at a concentration of 1%, and then sterilized. After cooling, each flask was inoculated with 1ml of 2х107 spore/ml and incubated at 30ºC for 120 h.
Effect of temperature
Five different temperatures (20-40ºC) were used. Each flask was inoculated with 1ml of 2х107spore/ml and incubated at a specific temperature for 120 h.
Partial purification of GA enzyme
Ammonium sulfate precipitation
Under refrigerated conditions, the crude enzyme solution was precipitated by different concentrations of (NH4)2SO4 (20-80%), and the precipitate was separated using a centrifuge at 10,000 rpm for 30 min. The precipitate was taken and dissolved in 5 ml of 0.1 M sodium acetate buffer (pH5). The activity of enzyme, the concentration of protein, and the specific activity were measured.
Dialysis
The dialysis process of the enzyme solution was carried out by using a 7000 (MWCo) dialysis bag against sodium acetate buffer 0.1 M, pH5 at 4°C for 24h and re-concentrated by sucrose. The activity of enzyme, the protein concentration, and the specific activity were measured. The enzyme solution was stored in the refrigerator at 4ºC until use.
Characterization of purified GA enzyme
The optimal pH on GA activity
Different pH ranges 2-8 were prepared. GA activity was assessed using starch as a substrate.
The metal ions effect on GA activity
Five mM concentration for each solution was prepared for MgSO4, MnSO4, FeSO4, and CaCl2 using phosphate buffer (pH5). 1ml of the enzyme was added to the previous concentrations. All test tubes were incubated at 30°C for 10 min. GA activity for all test tubes were measured.
Results![]()
Screening for hyper-producing strains of Rhizopus sp.
Test results for the isolate that produced the highest productivity of enzyme among the Rhizopus spp. isolates 1, 2, 3, 4, and 5 under study showed that the Glucoamylase index(GI) for these isolates was 1.4, 0.6, 1.2, 3.4, and 2.2, respectively. According to these results, the fungal isolate that showed largest clear zone, or GI, was selected to produce GA enzyme. The fungal isolate was later diagnosed as R. oryzae according to [8].
Optimization of the production process
Effect of Various Agro-Residues as Carbon Source
The results in figure1 revealed that the substrates varied in their ability to induce GA production. All the substrates (WB, RH, GNP, and CC) screened as carbon sources can support the growth and stimulate the production of GA by R. oryzae. Wheat bran produced the highest enzyme with specific activity 6.25 U/mg, followed by CC With specific activity 4.87 U/mg. At the same time, enzyme production was lower with GNP with specific activity 2.53 U/mg.
Effect of different nitrogen sources
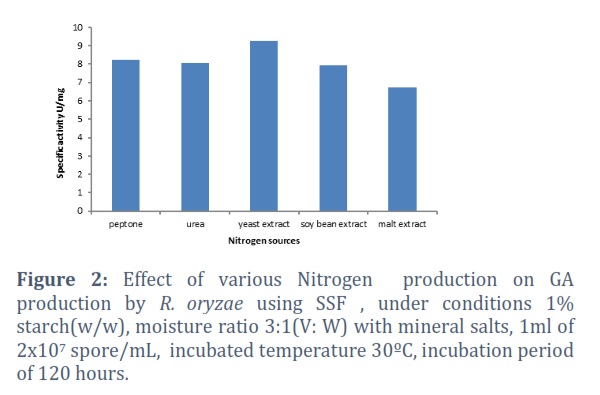
Data obtained in figure 2 revealed that yeast extract was the potent inducer for GA production by R. oryzae, and the specific activity was 9.28 U/mg. On the other hand, malt extract was the least effective inducer for enzyme production with specific activity 6.75 U/mg.
Effect of various temperatures
One of the factors that affect enzyme production is temperature. GA production was achieved at various temperatures (20, 25, 30, 35, and 40ºC). The most favorable temperature for GA production by R. oryzae was 30°C with specific activity 10.27U/mg. Nevertheless, the decrease or increase in the incubation temperature reduces enzyme production, as revealed in fig 3.
Partial purification of GA
After producing the crude enzyme from the R. oryzae fungus under the previous optimum conditions, the crude enzyme was purified using a series of steps, including precipitation and dialysis.
Ammonium sulphate (NH4)2SO4 precipitation
Gradient concentration 20-80 % saturation of (NH4)2SO4 was used for the precipitation of GA. Maximum specific activity 18.40U/mg was recorded at 55% saturation of (NH4)2SO4. This step achieved a high fold of purification and success in reducing the volume of the crude enzyme from 100 ml to 24 ml which facilitated working with it. The results showed an increase in the enzyme’s specific activity 11.80U/mg compared to the crude enzyme.
Dialysis
The results show an increase in specific activity 24.86 U/mg because of the increased concentration of the enzyme as a result of getting rid of the excess buffer solution.
Characterization of purified GA enzyme
The optimal pH of GA activity
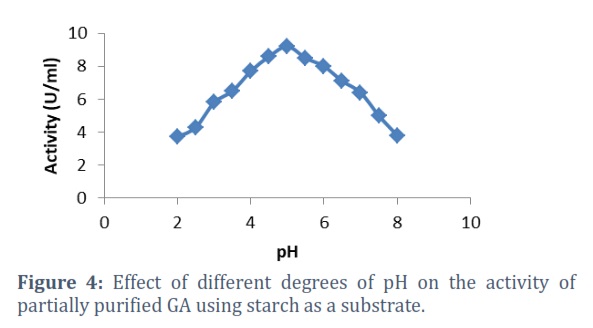
The results demonstrate in fig 4 that R. oryzae GA is active in a wide range of pH2-8 and it is not sensitive to pH change, but it is more active at pH5 (9.2U/ml) than other values.
Effect of metal ions on GA activity
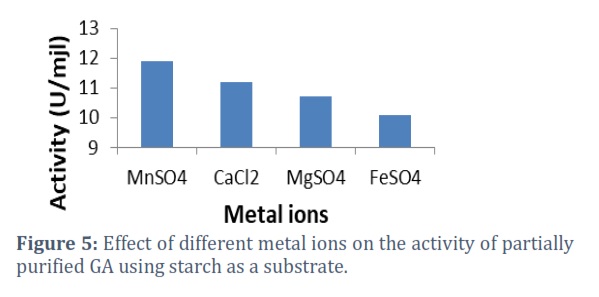
The results revealed that the highest enzyme activity was established with Mn2+ ions, followed by Ca2+,and then Mg2+. Thus, GA was established highly steady with MnSO4 (fig 5).
Figures & Tables
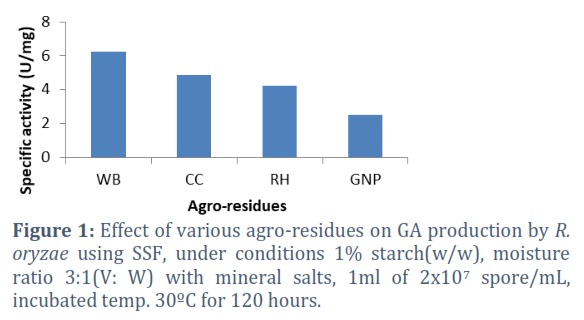

Starch is one of the rich sources of carbon that fungi need for the growth and production of GA [13]. Quantitative analysis of wheat bran indicates that it contain a large amount of starch, fibers, nitrogen and inorganic elements, so wheat bran is considered one of the important substrates that widely used in the field of scientific research, especially those concerned with the growth of fungi and the production of many enzymes, including GA [14]. The selection of an appropriate substrate for SSF has primarily been based on the more efficient use of various agro-industrial residues for enzyme production, as well as the potential benefits for the filamentous fungi, which can permeate into solid substrates with the assistance by the attendance of turgor pressure at the filament’s end [15,16].
Nitrogen sources have an extraordinary impact on microbial growth and the production of extracellular enzymes. The results in Figure 3 agree with [17], who revealed that adding peptone, and yeast extract to the culture medium invigorated GA formation by A. terreus. A complicated nitrogen source shows are necessary for production of a large amount of GA by R. oryzae. The prerequisite might be clarified by microorganisms' tendency to use proteins as nitrogen for growth, just as for extracellular enzyme formation. The GA synthesis by various microbes has been associated with the absence of different amino acids and complicated nitrogen sources in the growth medium [18].
All vital events in the cell are affected by temperature directly through the influence on enzymes and lipids in the cell membrane, which influences the quantity and speed of growth. When temperature increases to more than the optimum degree, this will lead to a rapid decline in the velocity of growth due to the denaturation of the enzyme; this is because of the rupture of weak bonds in the secondary and tertiary of enzyme construction [19].
One of the methods of concentrating and purifying enzymes is salting out proteins using (NH4)2SO4. Increases in the ionic strength of the solution create a decrease in the repulsion impact of like charges among undifferentiated particles of a protein [20]. It also reduces the forces around the protein molecules. When these forces are adequately reduced, the protein will precipitate. (NH4)2SO4 is appropriate and effective due to its high solubility, cheapness, and lack of toxicity to most enzymes [21].
Dialysis is the diffusion of molecules from high to low concentration via a permeable layer. Exclusively those molecules those are small sufficiently to able via the membrane's pores of are capable of moving via the permeable membrane and arriving to balance with the whole volume of solution. To remove further useless substances, it is necessary to displace the dialysis buffer. When the buffer changes, particles' movement from high to low concentration will recommence till balance [22].
Most enzymes undergo irreversible denaturation in strong acidic or alkaline conditions. pH can influence enzyme activity in several ways and with different mechanisms: groups ionization in the active site of the enzyme, groups ionization in the substrate, or influencing the shape of either the enzyme or the substrate [23], which could clarify the decrease in the enzyme activity at the great value of alkalinity (pH8) and acidity (pH2).
GA is known to be metal ions relying on enzymes [24]. The effect of metal ions on GA activity reported that Mn2+, Ca2+, Mg2+, and Fe2+developed GA. At the same time, GA is used with α-amylase which demands Ca2+ ions in the decomposition process; motivation of GA activity by Ca2+, Fe2+, and Mn2+ ions can make it extra appropriate for use in manufacturing starch biotransformation process.
The author declare that there is no conflict of interest regarding the publication of this paper.
![]() References
References
- Marlida T, Saari N, Hassan Z, baker j. Purification and characterization of sago starch degrading glucoamylase from Acremonium species endophytic fungus. Food Chemistry, (2000); 71(2): 221-7.
- Karim KMR, Tasnim T. Fungal glucoamylase production and characterization: A Review.Bioresearch Communications, (2018); 4(2):591-605
- Lam WC, Pleissner D, Lin CSK. Production of fungal glucoamylase for glucose production from food waste. Biomolecules, (2013); 3: 651-61.
- De Castro RJS, Sato HH. Enzyme production by solid state fermentation: general aspects and an analysis of the physicochemical characteristics of substrates for agro-industrial wastes valorization. Waste and Biomass Valorization, (2015); 6(6):1085-93.
- Jones EB, Pang KL, et al. An online resource for marine fungi. Fungal Diversity, (2019); 96(1): 347-433.
- Lawal A, Banjoko A, Olatpe S, Orji F. Production and partial purification of glucoamylase from Aspergillus niger isolated from Cassava peel soil in Nigeria. African Journal of Biotechnology, (2014); 13(21): 2154-8.
- Leisle JF, Summerell BA. The Fusarium laboratory manual. Wiley-Blackwell Publishing, (2006); 388.
- Zambare V. Solid state fermentation of Aspergillus oryzae for glucoamylase production on agro-residues. International Journal of Life Sciences, (2010); 4: 16-25.
- Ruann JS, Helia HS. Enzyme production by solid state fermentation: general aspects and an analysis of the physicochemical characteristics of substrates for agro-industrial wastes . Waste and Biomass Valorization, (2015); 6(6):1085-93.
- Miller GL. Use of dinitrosalicyclic acid reagent for determination of reducing sugar. Analytical Chemistry, (1959); 31(3): 426-8.
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Chemistry, (1976);72: 248-54.
- Imran M, Asad M, Gulfraz M, Qureshi R, Manzoor N, Choudhary A. Glucoamylase production from Aspergillus niger by using solid state fermentation process. Pakistan Journal of Botany, (2012); 44(6): 2103-10.
- Pandey A. Solid-state fermentation. Biochemical Engineering Journal, (2003); 13(2-3), 81-4.
- Sadh PK, Kumar S, Chawla P, Duhan JS. Fermentation: A Boon for production of bioactive compounds by processing of food industries wastes (by-products). Molecules, (2018); 23:1-33.
- El-Gendy MMA, Alzahrani NH. Solid state fermentation of Agro-industrial residues for glucoamylase production from Endophytic Fungi Penicillium javanicum of Solanum tuberosum L. Journal of Microbial and Biochemical Technology, (2020); 12(1):1-9.
- Ali S, Mahmood S, Alam R, Hossain Z. Culture condition for production of glucoamylase from rice bran by Aspergillus terreus. MIRCEN journal of applied microbiology and biotechnology, (1989); 5: 525–32.
- Kumar p, Satyanarayana T. Microbial glucoamylases: characteristics and applications. Critical Reviews in Biotechnology, (2009); 29(3):225-55.
- Adefisoye SA, Sakariyau AO. Production of glucoamylase by Aspergillus niger in solid state fermentation. Advances in Biological Research, (2018); 12 (1):7-11.
- Labrou N. Protein Purification : An Overview. Methods in molecular biology, (2014); 1129:3-10.
- Mothe T, Sultanpuram VR. Production, purification and characterization of a thermotolerant alkaline serine protease from a novel species Bacillus caseinilyticus. 3 Biotech, (2016); 6(1):53.
- Kavya P, Bhat Sk, Siddappa S, Rao AG, Kadappu KB, Marathe Gk. When to Avoid Dialysis during Protein Purification. Advances in Industrial Biotechnology, (2019); 2(1): 9-23.
- Daniel RM, Danson MJ, Eisenthal R, Lee KC, Peterson ME. The effect of temperature on enzyme activity : New insights and their implications. Extremophiles, (2008) ; 12(1):51-9.
- Deb P, Talukdar SA, Mohsina K, Sarker PK, Abusayem SM . Production and partial characterization of extracellular amylase enzyme from Bacillus amyloliquefaciens-P-001. SpringerPlus,(2013); 2(1):154.
- Kareem SO, Akpan I, Popoola S, Sanni LO. Purification and characterization of thermostable glucoamylase from Rhizopus oligosporus SK5 mutant obtained through UV radiation and chemical mutagenesis. International Journal of Biotechnology, (2014); 26(1):19-24.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


