

Full Length Research Article
Bacteriophage Endolysins Antibacterial Effect against S. mutans Isolated from Dental Caries: An In-Vitro Study
Huda Mohammad Hasan Ali1*, Adeeb Ahmed kadhim Al-Zubaidy2, Ahmed Sahib Abdulamir3
Adv. life sci., vol. 11, no. 2, pp. 444-448, May 2024
*- Corresponding Author: Huda Mohammad Hasan Ali (drhuda1984@hotmail.com)
Authors' Affiliations
2. Department of Pharmacology, College of Medicine, University of Warith Al-Anbiyaa, Karbala – Iraq
3. Department of Pharmacology, College of Medicine, Al-Nahrain University, Baghdad – Iraq
[Date Received: 17/09/2023; Date Revised: 01/02/2024; Date Available Online: 18/04/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Many bacterial infections including dental caries that caused by Streptococcus mutans (S. mutans) were difficult to be treated due to the unavailability of effective antibiotics. Therefore, it requires to focus the attention on endolysin as an antibacterial agent. This study was designed to evaluate the In-vitro effectiveness of specific bacteriophage endolysins against S. mutans isolated from dental caries.
Methods: Twenty-five isolates of S. mutans were collected from patients with dental caries attended to Al-Imamain Al-Kadhimain, Medical City Hospital, and AL-Ameen Health Center, during the period from December 2021 to May 2022. The S. mutans isolates were identified by culturing them on their specific mitis salivarius sucrose bacitracin tellurite media (MS-SBT agar), morphological characteristics, gram staining, biochemical tests and VITEK 2 compact system. Seven of these S. mutans isolates were used for the in-vitro study.
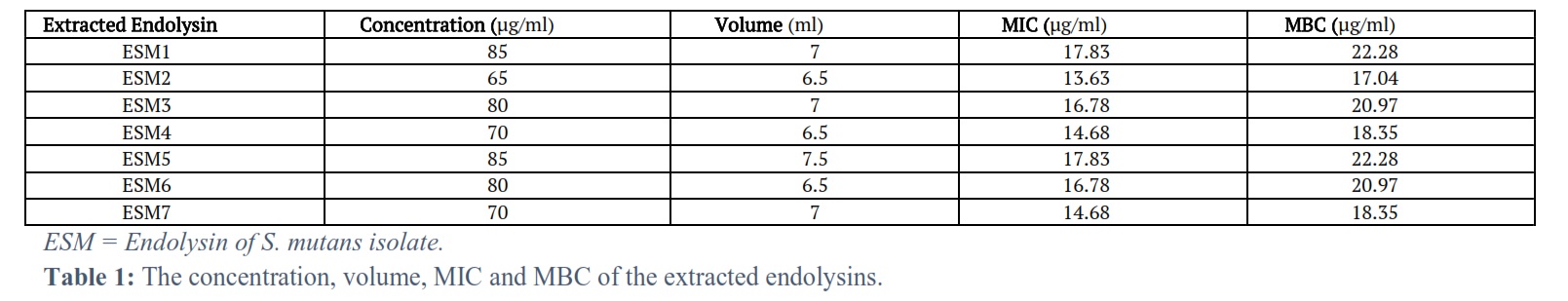
Results: The bacteriophage endolysin enzyme extracted from the corresponding S. mutans isolate was effective as In-vitro antibacterial agent with minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) ranging between (13.63 – 17.83) µg/ml and (17.04-22.28) µg/ml respectively.
Conclusion: The endolysin was effective as antibacterial agent against multi-drug resistant (MDR) S. mutans upon growing and testing on culture media.
Keywords: S. mutans; Bacteriophage; Endolysin; Dental caries
Introduction![]()
The gram-positive bacterial lysins have evolved to bind to their cell wall binding receptors at a high affinity to limit the release of free enzyme [1]. Streptococcus mutans (S. mutans) is a facultatively anaerobic, Gram-positive coccus commonly survived in the oral cavity; so, it is considered one of the primary causative agents of dental caries [2].
Bacteriophages are the most abundant entities on earth. They are bacterial viruses that have genetic material in the form of either RNA or DNA, encapsidated by a protein coat [3]. This capsid is attached to a tail which has fibers that are used for attachments to receptors present on the bacterial cell surface. Most of the bacteriophages have polyhedral capsid except filamentous phages [4]. Phages infect bacteria through two possible ways; lytic life cycle and lysogenic life cycle. When these viruses multiply, they kill their hosts, and the life cycle is called lytic life cycle. On the other hand, in the lysogenic life cycle, some phages (temperate phages) can grow vegetatively and can insert their genome into host chromosome replicating with their hosts for many generations [5].
Bacteriophage endolysins are a new and novel class of anti-bacterial agents. They are highly developed enzymes produced by bacteriophage to digest the cell wall by cleaving the essential bonds in the bacterial cell wall peptidoglycan for phage offspring release. In Gram-positive bacteria, small quantities of purified lysin added externally results in immediate lysis causing log-fold death of the target bacterium [6, 7].
As a mechanism of action of endolysins; basically, there is a three-step model system of the bacterial host lysis. This model consists of three proteins named endolysin, holin, and spanin. These proteins perform actions on the bacterial outer membrane, inner membrane, and peptidoglycan and can cause the lysis of the host. The degradation of the peptidoglycan is initiated when holin makes small pores in the host inner membrane; then, lysis is initiated by releasing endolysin inside the periplasm and degrading the peptidoglycan and according to the new findings, spanin, which is another lysis protein, is required for the degradation of the outer membrane. The lysis of the bacterial host cell occurs through this mechanism [8]. The release of phage progeny at the end of the lytic cycle is facilitated by holins and lysins [9].
The present research aims to assess the In-vitro efficacy of particular bacteriophage endolysins against S. mutans strains isolated from dental caries.
Methods![]()
Seven of these isolates that were used for the In-vitro study were collected from patients with dental caries attainted to Al-Imamain Al-Kadhimain, Medical City Hospital, and AL- Ameen Health Center, from December 2021 to May 2022. The identification and susceptibility tests were done to these isolates by VITEK 2 GP device system.
For the isolation of bacteriophage, a number of samples of sewage, feces of sheep, chicken litter and bird litter were collected. Infectious phages were separated by phage spot test using a nutrient-agar. The plaques were counted by top layer agar plaque assay to determine the phage titer, morphological characteristics, edge, clarity and size. Furthermore, top layer plaque assay is the best way used for getting highest titers of phage suspensions [10, 11]. The phage titer was calculated by using the following formula:
![]()
Extraction and purification of endolysin was done by precipitation by ammonium sulfate solution which was gradually added to crude sample to obtain a saturation ratio of 85 % at 4 ºC. The supernatant was removed, and the resultant precipitate was dissolved in a suitable volume of potassium phosphate buffer (50 mM, pH 7.0). Then dialysis of protein solution was done against distilled water (D.W.) for 24 hours [12]. Then, purification by gel filtration chromatography using Sephadex G-100 was done [13]. Finally, the molecular weight (M.W.), the concentration of endolysin, MIC and MBC were measured.
Assay of endolysin antibacterial activity: The bacterial broth titer was measured using serial dilution, then 10 µl (0.01 ml) of each dilution were spotted on nutrient agar, then incubated in 10 % CO2 at 37 °C for 48 hours. After incubation, the dilution plate that was showed countable separated colonies was used to calculate the bacterial titer (obtaining 0.5 McFarland) using the following equation [10, 14].
![]()
At the same time, 100 µl from this bacterial broth was incubated for 30, 60, 90 and 120 minutes with one MIC of endolysins with original concentrations of (85, 65, 80, 70, 85, 80, 70 µg/ml) according to the corresponding bacteria and their bacteriophage after extraction [15]. Also, one MIC of these endolysins were mixed with 100 µl bacterial broth and incubated in 10 % CO2 at 37 °C for 48 hours. The titer of bacterial broth plus endolysin was calculated using serial dilution and culture on the nutrient agar in 10% CO2 at 37 °C for 48 hours. The dilution plate that showed countable colonies was used to calculate the titer using the same above equation. The reduction in the titer of the bacterial broth with endolysin compared to the same broth without endolysin was measured. The endolysin antibacterial activity was quantified in log units decrease [10, 15].
Results![]()
Collection and isolation of S. mutans isolates: Seven isolates of S. mutans were isolated from the patients' oral cavity with dental caries. When they were cultured on MS-SBT agar plates, they were grown and taken the blue color of this medium. They were shown α-hemolysis upon growing on blood agar plates. The susceptibility tests for the isolates: Susceptibility tests were done by VITEK 2 GP device system. The results were shown that all the seven isolates that tested for the In-vitro study were MDR. The Bacteriophage characteristics: Seven bacteriophages that were active against S. mutans were produced detectable plaques. The diameter, margin cut, plaque shape and turbidity/clarity were the main characteristics that were determined. The plaque size was between 1.5 – 2.6 mm with oval and circular shape, clear and semi-clear (turbid) and regular and irregular margin cut. The titer of the isolated bacteriophages was between 109-1011 PFU/ml.
Characterization of the extracted endolysins
Endolysin molecular weight: The M.W. of the extracted endolysins ranged between 35 – 40 kDa. The concentration, volume, MIC, and MBC of endolysin: The concentration, volume, MIC and MBC of the extracted endolysins were ranging between (65-85) µg/ml, (6.5-7.5) ml, (13.63-17.83) µg/ml and (17.04-22.28) µg/ml respectively (Table 1).
The endolysin enzymatic activity assay
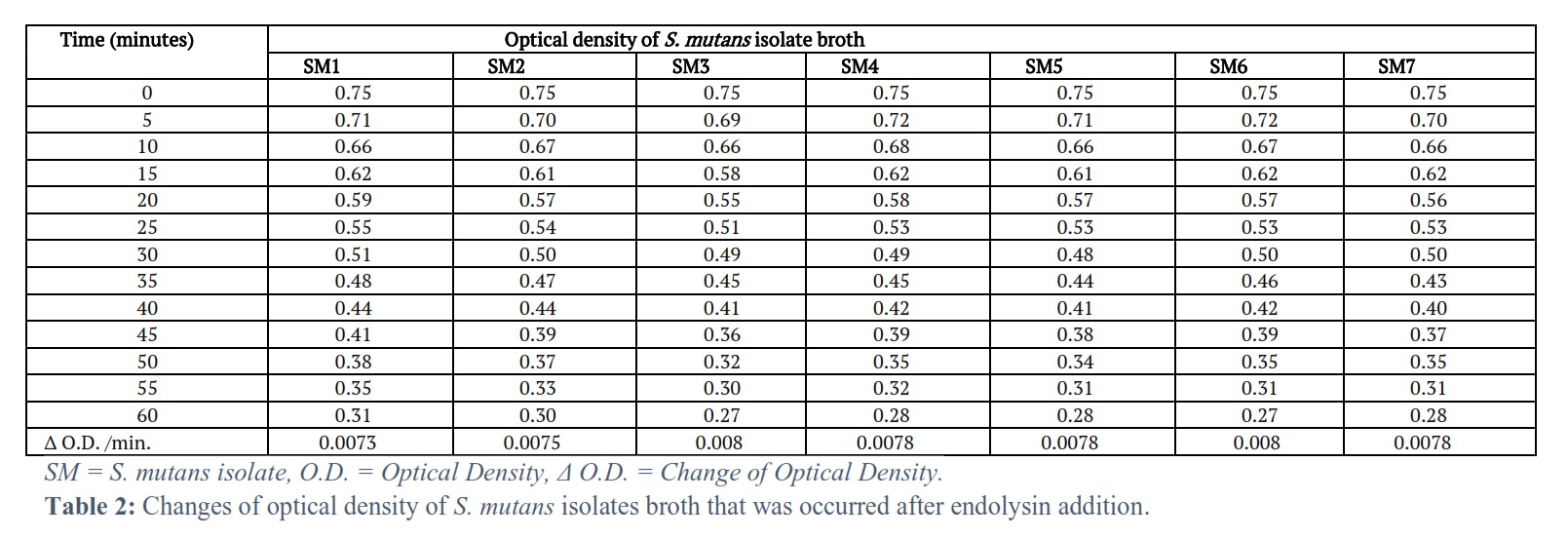
The optical density of the tested S. mutans isolate broth was clearly reducing with time (Tables 2 and 3).
Antibacterial activity assay of endolysin
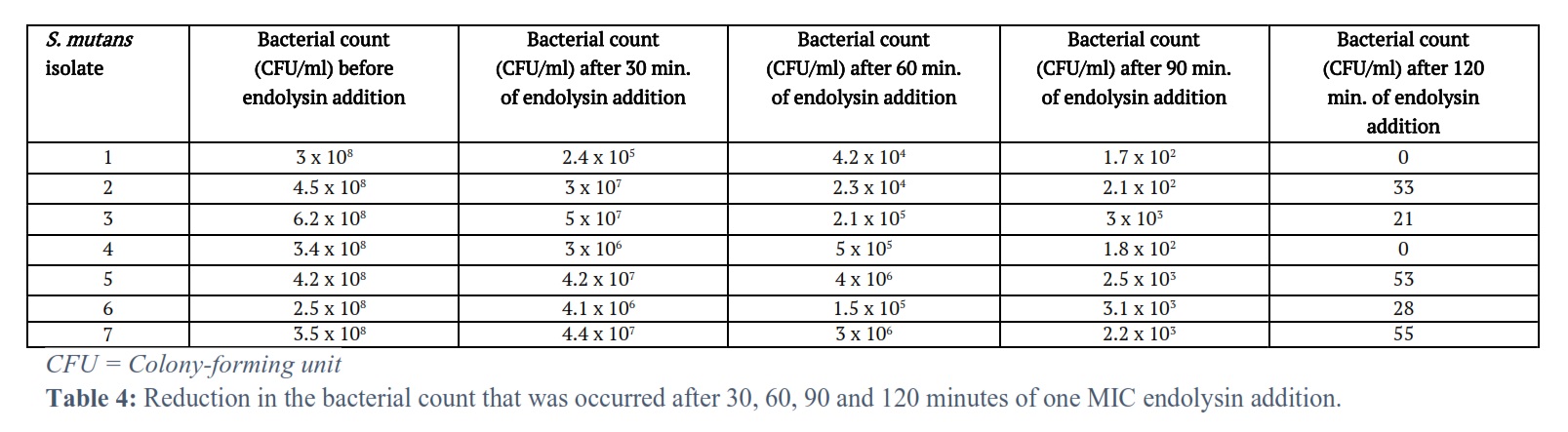
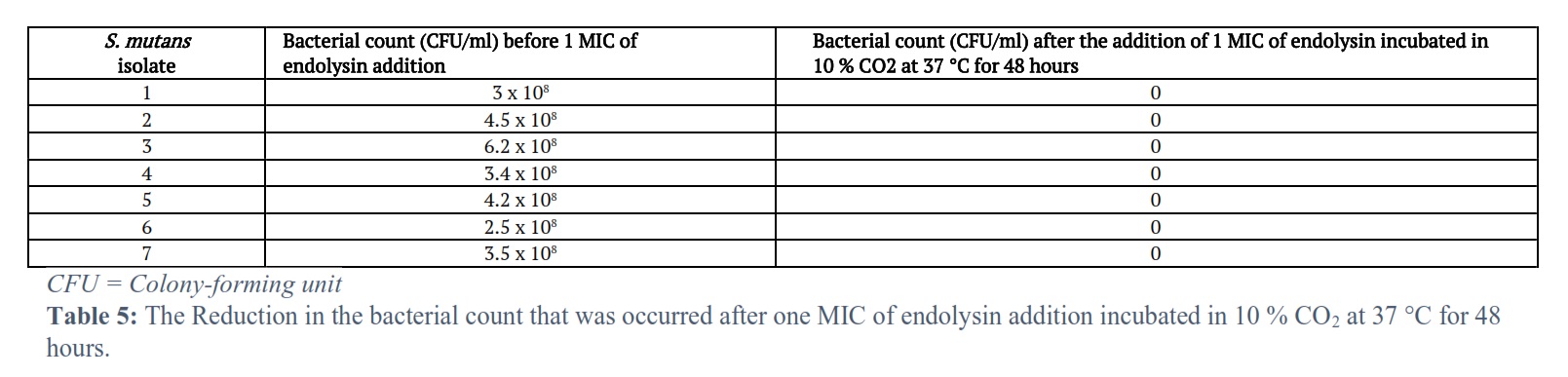
The results of the seven S. mutans isolates were showed a decrease in the bacterial count after 30, 60, 90 and 120 minutes of one MIC of endolysin addition (Table 4) and also after addition of one MIC of endolysin incubated in 10 % CO2 at 37 °C for 48 hours (Table 5).
Figures & Tables

Gram-positive endolysins have developed to bind to their binding receptors present in the cell wall at a high affinity to limit the release of free enzyme. Therefore, endolysins have been confirmed as effective antibacterial agents against most Gram-positive bacteria [1, 16].
The most widely occurrence of bacterial resistance to the available antibiotics was due to the use of inappropriate antibiotics for not suitable conditions and the use of antibiotics in high doses more than the required doses and for long period of time. All the seven extracted endolysins for the In-vitro study showed a high lytic activity whether by measuring the optical density of S. mutans isolates after the addition of their corresponding endolysins or by bacterial counting after one MIC of endolysin addition (bacterial counting after 30, 60, 90 and 120 minutes of the addition and after incubation of S. mutans isolates with their corresponding endolysins) in 10 % CO2 at 37 °C for 48 hours).
The optical density for all the seven tested S. mutans isolates was reduced with time, from time zero, just before the corresponding endolysin addition and after each 5 minutes after the addition of the corresponding endolysin for one hour. This resulted in optical density changes/min. 0.0073, 0.0075, 0.008, 0.0078, 0.0078, 0.008, 0.0078 for the S. mutans isolates 1, 2, 3, 4, 5, 6, 7 respectively. The bacterial count (CFU/ml) of all seven tested S. mutans isolates was reduced with time after 30, 60, 90 and 120 minutes of one MIC of endolysin addition. This resulted in a decrease in the bacterial count at the end of the 120 minutes 0, 33, 21, 0, 53, 28, 55 for the S. mutans isolates 1, 2, 3, 4, 5, 6, 7 respectively.
Also, the bacterial count (CFU/ml) of all seven tested S. mutans isolates was reduced with time after one MIC of endolysin addition incubated in 10 % CO2 at 37 °C for 48 hours. This resulted in a decrease in the bacterial count at the end of the incubation period to zero (no bacteria) for all the seven tested S. mutans isolates. The optical density changes and the bacterial count (CFU/ml) in this study are agreed with the results of study that was done by Ali et al. [15] with optical density changes/min. ranging between (0.013 – 0.015) and bacterial count (CFU/ml) at the end of 120 minutes (zero). The study of Hussain et al. [17] in Baghdad had results showing decrease in the bacterial growth when treated with endolysin in which the antibacterial effect of the gel formulation of endolysin was demonstrated by adding 10 ml of the gel onto the nutrient agar plates and incubating under aerobic conditions for 24 hours at 37°C. The plates were tested, and a clear zone indicating that the gel was active in bacterial killing.
Antibacterial effect of endolysins for the In-vitro study was dependent on many factors like pH range, buffers, temperature, and ionic strength [18]. Also there were many other factors that might be the cause of the difference from one study to others including: the volume of endolysin used in these studies which might differ from the volume of endolysin used in the current study which was one MIC, the degree of susceptibility of bacteria (sensitive intermediate or resistant), technical changes and the changes in the strength of the bacteriophages from which endolysins were extracted, the more potent bacteriophage was used, the more potent endolysin was obtained because the major effect of bacteriophage is attributed to the endolysin action.
Therefore, there is a difference in the results from one study to others; but almost the studies concerning endolysin In-vitro effect have the same effects of reducing the bacterial growth.
All S. mutans isolates were MDR. The seven extracted endolysins for the In-vitro study were showed a high lytic activity whether by measuring the optical density of S. mutans isolates after the addition of their corresponding endolysins or by bacterial counting after one MIC of endolysin addition (bacterial counting after 30, 60, 90 and 120 minutes of the addition and after incubation of S. mutans isolates with their corresponding endolysins in 10 % CO2 at 37 °C for 48 hours.
Conflict of Interest
The authors declare that there is no conflict of interest.
HA did all the lab work, and collected the data, while AALZ and AAB supervised all research steps, HA, AALZ and AAB participated equally in research design, data analysis and write the final version of the article.
![]() References
References
- Loessner MJ, Kramer K, Ebel F, Scherer S. C-terminal domains of Listeria monocytogenes bacteriophage murein hydrolases determine specific recognition and high-affinity binding to bacterial cell wall carbohydrates. Molecular Microbiology, (2002); 44(2): 335-349.
- Banas JA. Virulence properties of Streptococcus mutans. Frontiers in Biosciences, (2004); 9: 1267-1277.
- Clark JR, March JB. Bacteriophages and biotechnology: vaccines, gene therapy and antibacterials. Trends in Biotechnology, (2006). 24(5): 212-218.
- Ackermann HW. Tailed bacteriophages: the order caudovirales. Advances in Virus Research, (1998); 51: 135-201.
- Inal JM. Phage therapy: a reappraisal of bacteriophages as antibiotics. ArchivumImmunologiae et TherapiaeExperimentalis(Warsz), (2003); 51(4): 237-244. (PMID: 12956433).
- Fischetti VA. Lysin Therapy for Staphylococcus aureus and Other Bacterial Pathogens. Currrnt Topics in Microbiology and Immunology, (2017); 409: 529-540.
- Fischetti VA. Bacteriophage endolysins: a novel anti-infective to control Gram-positive pathogens. International Journal of Medical Microbiology, 2010; 300(6): 357-362.
- Cahill J, Young R. Phage Lysis: Multiple Genes for Multiple Barriers. Advces in Virus Research, (2019); 103: 33-70.
- Wang IN, Smith DL, Young R. Holins: the protein clocks of bacteriophage infections. Annual Reviews in Microbiology, (2000); 54: 799-825.
- Ezzeldin N, Elshishtawy H, Mohamed M. Molecular assessment of oral Streptococcus mutans isolated from patients with different ages and caries activity via selective media and protein pattern. Egyptian Dental Journal, (2021); 67: 780-800.
- Budzik JM, Rosche WA, Rietsch A, O'Toole GA. Isolation and characterization of a generalized transducing phage for Pseudomonas aeruginosa strains PAO1 and PA14. Journal of Bacteriology, (2004); 186(10): 3270-3273.
- Virgen-Ortíz JJ, Ibarra-Junquera V, Osuna-Castro JA, Escalante-Minakata P, Mancilla-Margalli NA, Ornelas-Paz Jde J. Method to concentrate protein solutions based on dialysis-freezing-centrifugation: enzyme applications. Analytical Biochemistry, (2012); 426(1): 4-12.
- Lakshmi BKM, Muni Kumar D, Hemalatha KPJ. Purification and characterization of alkaline protease with novel properties from Bacillus cereus strain S8. Journal of Genetic Engineering and Biotechnology, (2018); 16(2): 295-304.
- Harley JP. Colony Forming Unit and Calculation. Laboratory Exercises in Microbiology. McGraw-Hill Higher Education; 6th Edition; 1 April 2004.
- Ali MR, Abdulamir AS, Kadhim SR. Extraction, purification and therapeutic use of bacteriophage endolysin against multi-drug resistant Staphylococcus aureus: In vivo and In-vitro study. Journal of Contemporary Medical Sciences, (2018): 4 (1): 33– 38.
- Rashel M, Uchiyama J, Ujihara T, Uehara Y, Kuramoto S, Sugihara S, et al. Efficient elimination of multidrug-resistant Staphylococcus aureus by cloned lysin derived from bacteriophage phi MR11. Journal of Infectious Diseases, (2007); 196(8): 1237-1247.
- Hussain HE, Abdulamir AS, Farhood IG, Ali AR, Al-azzawy MA. In-vitro and in vivo study for antibacterial activity of endolysin-HEC gel and mixture gel on acne vulgaris caused by multidrug-resistant Staphylococcus aureus bacteria. Journal of Population Therapeutics and Clinical Pharmacology, (2023); 30(1): e242–e257.
- Oliveira H, São-José C, Azeredo J. Phage-Derived Peptidoglycan Degrading Enzymes: Challenges and Future Prospects for In Vivo Therapy. Viruses, (2018); 10(6):292.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


