

Review Article
The Concept of Chloroplast Transformation; Its Relevance Towards Food Security
Bushra Tabassum1, Iqra Yousaf2, Olawale Samuel Adeyinka3, Rida Khalid1, Anwar Khan4
Adv. life sci., vol. 11, no. 1, pp. 28-39, February 2024
*- Corresponding Author: Bushra Tabassum (bushratabassum.sbs@pu.edu.pk)
Authors' Affiliations
2. Centre of Excellence in Molecular Biology, University of the Punjab, Lahore – Pakistan
3. Department of Chemistry, Physics and Atmospheric Sciences Jackson State University, Jackson – USA
4. Department of Microbiology, BUITEMS, Quetta – Pakistan
[Date Received: 09/11/2022; Date Revised: 25/07/2023; Date Published: 25/02/2024]
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
The world population is expected to reach an estimated 9.2 billion by 2050 which means up to 70% increase in food production would be needed to feed the world. Moreover, climate changes pose yet another challenge to global food security. Genetic engineering is playing its role in modern agriculture to overcome food security challenges but for better crop production more biotechnological advances are required. Chloroplast genetic engineering provides a better alternative to nuclear genetic engineering due to its properties of precise transgene insertion through homologous recombination, lack of epigenetic changes and high transgene expression. We reviewed the fundamentals behind chloroplast transformation and divulge its various applications in nutritional enhancement and agricultural stress management. Furthermore, we provide recent efforts of Ribonucleic Acid interference (RNAi) technology in the transplastomic transformation towards insect-specific and environmentally friendly control measures. We envisage that targeted chloroplast genome editing could be a game changer in the future application of chloroplast transformation.
Keywords: Agricultural stress management; Chloroplast transformation; Genome editing; RNA interference; Transgenic crops
Introduction![]()
Heritable genetic materials in plant cells are contained in three cellular compartments: the nucleus, plastids, and mitochondria. The huge success recorded in transgene plants has been through the incorporation of the targeted gene into the nucleus. However, transplastomic could offer a better alternative for sustainable food security. Plant cells and eukaryotic algae contain an organelle called chloroplast that is important for photosynthesis , the production of compounds like fatty acids, amino acids, nucleotides, phytohormones, vitamins, secondary metabolites, and the integration of sulfur and nitrogen [1,2]. Chloroplast has a highly polyploid circular genome (plastome) and specific genetic machinery for transcription/translation [3]. The machinery was supposed to have evolved from endosymbiotic cyanobacteria that were incorporated into an ancestral eukaryotic host cell [4]. The genome of a plastid contains about 120- 130 genes that are highly conserved and structured in a quadripartite manner. The structure comprises two (IRs: IRA and IRB) inverted repeat regions, that divide the genome into small single-copy (SSC) and large single-copy (LSC) sections. Despite inclusive conservation in structure, the genomic size of chloroplast differs between species, including (Cathaya argyrophylla) 107 kb to (Pelargonium) 218 kb [4, 5]. The variations within the inverted regions give rise to the various plastid species' varied genomic sizes yet tRNA genes and protein-coding genes share many similarities since IRs typically have identical nucleotide sequences [6].
Since 80’s when the tobacco (Nicotiana tabacum) chloroplast genome was first sequenced [7], there has been drastic progress in other plant’s chloroplast genomes. Currently, there is the availability of about 800 complete sequences of the chloroplast genome in the organelle genomic database of (NCBI). There is also an important contribution of chloroplast genomes made to the phylogenetic analysis of numerous plant families and aided the understanding of evolutionary relationships within phylogenetic clades [8]. The Genome sequences of chloroplast have revealed considerable sequence and structural variation in the interspecific and intraspecific plant species. Such variant information has been particularly valued for the knowledge of how climate affects cash crops, favoring the reproduction of closely related species and the discovery and preservation of immensely useful characteristics. Information on chloroplast genomes has been used to study the cultivation of various crops, particularly legumes [9].
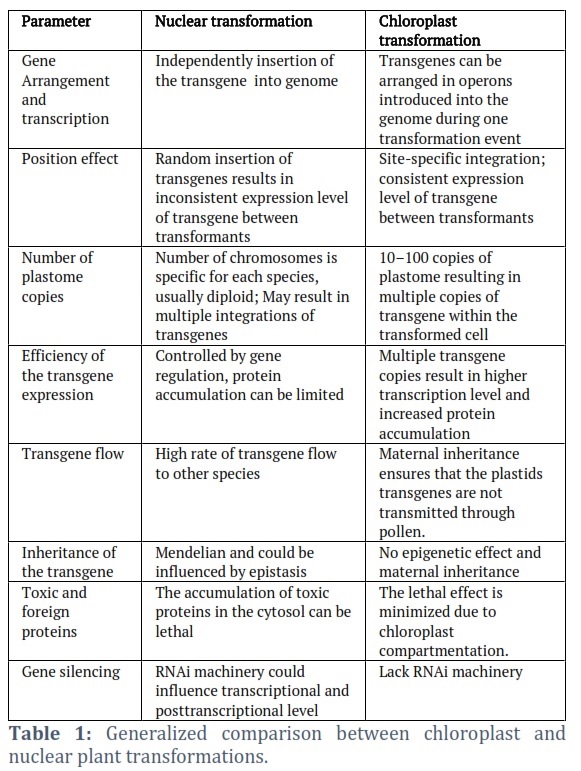
Breakthrough in the chloroplast genome integration to Chlamydomonas and tobacco [10, 11] has unlocked innovative opportunities to modify other crops' chloroplast genomes. The absence of epigenetic effects and maternal inheritance of plastid gives it an edge over nuclear transformation. Unlike nuclear transgene (Table 1); the transplastomic plant could express several external proteins from the chloroplast genome at very high levels.
This is a result of the plastids containing thousands of chloroplast genome copies per cell and the absence of gene silencing. Furthermore, it provides high recombinant protein levels with the viability of multiple genes expression from operons and the absence of position effects [12]. The chloroplast genes responsible for photosynthesis are of three categories. The first group consists of: the photosystem I and II (psaA, psaB), (psbA, psbB) respectively, ATP synthase (atpA, atpB), the large subunit of RubisCO (rbcL), cytochrome 6bf (petA, petB) and NAD(P)H dehydrogenase genes (ndhA, ndhB). The second consists of a group of RNA genes and accounts for the genetic machinery in the chloroplast like ribosomal RNA (rrn5, rrn16), RNA polymerase (rpoA, rpoB), transfer RNA (trnK, trnH) and ribosomal subunit genes (rps2, rps3). The third is mainly composed of open reading frames for preserved plastids (ORFs) and genes that code for proteins (cemA, matK,) [13]. IR regions have been presumed to be more suitable for transgene insertion because they occur in duplicate in most chloroplast genomes. The integration site plays an important role in the transfer of gene expression levels and in the enhancement of homoplasmy under antibiotics selection.
Plastid transformation occurs by the targeted gene integration at a predictable site via the plastid genome and the transformation vector recombining in a homologous manner. The transgenic cassette's inclusion into one copy of the IR aids incorporation into the other duplicates. Thus, exert more selection pressure to achieve homoplasmy via the plastid genome's replica repair system [14,15]. In addition, the highly conserved intergenic spacer region is the base concept of universal vector among the higher plant chloroplast genomes and is utilized to transform the plastomes of many related plant species [15] because the plastids have their own unique genetic machinery for transcription/translation. However, non-related species or species with significantly less homology to the said flanking region may result in low expression which might necessitate a species vector for each particular species to be transformed successfully. Due to positive advances in biotechnology, similar methods are employed to integrate foreign genes into both the nuclear genome and chloroplast genome. The effective delivery of foreign DNA across the double membrane of the plastid, the efficient transplastome selection as well as the integration of the heterologous DNA, are the-all marks of effective chloroplast transformation.
Methods![]()
Literature Search and Selection Criteria
A systematic search was carried out from Google Scholar, NCBI and Google Web Browser by providing key terms “chloroplast transformation”, vectors, promoters, plastid engineering, role of flanking regions in plastid transformation, homoplasmic transgenic lines, plant based edible vaccines, plastid and transgene expression, plant based Biopharmaceutical products, commercial application of chloroplast genetic engineering, plant protection through chloroplast transformation etc. The literature found was further screened for inclusion according to their contents. In this study, 80 peer reviewed research articles were selected.
Discussion![]()
Methods of transgene integration in the chloroplast genome
There are various reported techniques for introducing foreign DNA into the genome of the chloroplast. Among reported methods of delivering foreign DNA into the chloroplast genome, biolistic (particle bombardment) DNA delivery appears to be the most popular. However, there are reports of stable plastid transformation using a direct process based on polyethylene glycol (PEG)). Although the particle bombardment technique is very expensive, its benefits include high efficiency of transformation and quick regeneration of transformed tissues while polyethylene glycol (PEG) is less expensive but requires high technical know-how, especially during protoplast isolation.
Particle bombardment: This approach is centered on a basic concept that involves the rapid acceleration of DNA-coated particles (micro projectiles) into living tissues or cells directly. It can be a PDS-1000/HeTM gadget or a gunpowder model. In the gunpowder, a small aqueous suspension of DNA-coated tungsten powder is placed at the front of a plastic macro projectile that resembles a bullet. The macro projectiles are ejected through a small hole, while the micro projectiles are accelerated by the gunpowder charge. The PDS-1000/HeTM device will discharge a microcarrier that will be propelled by a burst of helium gas molecules coated with millions of dried DNA-coated micro carriers.
Since the 1980s, DNA delivery into cells using high-density macro projectiles has undergone several advancements since the first effective delivery of DNA to a plant was announced. This technique has undergone several improvements to reduce the destruction of target tissue samples and cell damage, and improve delivery efficacy. Hedtke et al. [16] suggested a more effective technique called Bio-Rad PDS-1000/He particle gun system which transports gold particles coated with a target gene and the selectable marker gene bar at 650 psi rupture pressure to modify the wheat immature embryos.
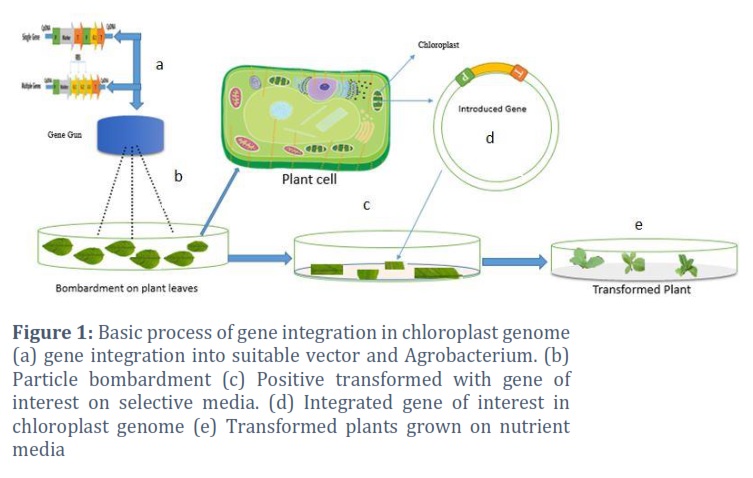
The advantages of this technique include its broad range of applications in the transformation of recalcitrant plants, monocotyledons, and dicotyledons. False positive results are minimal, and device operation is simple and only requires a small quantity of plasmid DNA. Particle bombardment has been used in plastid transformation (Figure 1).
Liere et al. [17] presented a replicable plastid transformation method using two distinct tobacco-specific plastid vectors, (Prrn/GFP/Trps16: PpsbA/aadA/TpsbA) and pZS197 (Prrn/aadA/TpsbA), designed for integration into the huge single copy, potato chloroplast genome with inverted repeat sections, and plant regeneration with uniformly modified plastids.
Polyethylene Glycol (PEG): This transformation technique requires protoplast extraction; the protoplasts are then given one more treatment with a solution that contains DNA, PEG, and different ions. In the process, the plasma membrane allows the DNA to pass through and enter the cytoplasm. The design vectors with sequences homologous to a particular target region in the plastid genome are subsequently incorporated into the target plastid genome. Antibiotics are used to separate homoplastomic cells from segregating modified plastome copies. B-tubulin with a hemagglutinin (HA) tag has been momentarily expressed in canidioschyzon merolae; the expression and the localization of the gene product were validated by immunocytochemistry 24 hours after PEG-mediated transformation [18]. Stoppel And Meurer [18] demonstrated an easy and economical stable plastid transformation in tobacco that was accomplished by treating leaf protoplasts with the transforming DNA, called polyethylene glycol (PEG) treatment.
The Essential components of chloroplast vector and putative function
Typical chloroplast vectors consist of a promoter; selective marker gene, 5′-UTR, and the gene of interest. The 5′ untranslated region within two chloroplast DNA flanking regions facilitates insertion by double homologous recombination into the chloroplast genome.
Flanking site vs Promoters: For an effective homologous recombination event and endogenous regulatory sequences for the targeted transgenic expression, particularly flanking DNA sequences are essential [19]. For the effective transfer of the desired gene into the plastid genome, there are following Flanking sites have been used include; rbcl/accD ,rps12/trnV and trnM/trnG. However, it was discovered that the chloroplast genome’s trnI/trnA genes, which are flanked by the ribosomal operon, are the most effective site for transgene incorporation over plastid transformation. Because trnA site has been demonstrated that the transgene’s location in the inverted repeat region, the presence of a replication origin, and the intron sequence are all linked to its increased copy number [20]. Studies have shown that the integration of the transgene into the transcriptionally active spacer region (trnl/trnA) resulted in the expression of transgenes about over 25-fold higher than the transcriptionally silent spacer region (rbcl/accD) [21]. This may be because multiple promoters present either heterologous or endogenous which increases transcription. Plastid transcription is accomplished mainly by two plastid RNA polymerases (RNAP); plastid-encoded plastid RNAP (PEP) and nucleus-encoded RNA polymerase (NEP). Various plastid promoters present which have both transcriptions start sites including PEP and NEP. The PEP core subunits are coded for within the plastome as rpoA, ropB, rpoC1,rpoC2 respectively The plastome encodes for the PEP core subunits ((α)2, β, β’, and β’’) as rpoA, ropB, rpoC1, and rpoC2, respectively While there is a link between T-type bacteriophages' RNA-synthesizing enzymes and single subunit nucleus-encoded RNA polymerase (NEP) [20,21].
Most plastid promoters contain the consensus sequences for sigma factors -35 (TTGaca) and -10 (TATaaT) which are nearly identical to bacterial RNAP [22]. Therefore proving the plastids’ cyanobacterial ancestry. The plastid transformation vector contains plastid promoters, often the 16S rRNA promoter (Prrn16) or the psbA promoter, which are used to insert transcriptionally transgenes into the chloroplast genome (PpsbA). While PpsbA’s transcription start site only has a PEP, Prrn’s transcription start site has both PEP and NEP [22]. It is crucial to remember that the main transcription products of both NEP and PEP activities are polycistronic. Thus, it needs substantial posttranscriptional modification like Intron removal, mRNA processing into monocistronic or oligocistronic forms, trimming of the 5′ and 3′ ends, and RNA editing [23].
Untranslated Regions (5’ and 3’): The interaction of RNA sequences and structures with other active elements in gene expression, stabilizes plastid mRNAs through RNA secondary structures together with their 5' and 3' UTRs. The synthesized proteins need protection from chloroplast proteases because they can be easily degraded by chloroplast proteases. Plastid transcripts' 5' untranslated region (UTR) has a significant impact on how much protein is produced when chimeric transgenes are expressed. There are various available 5’ UTR with various degrees of transcript longevity and protein stability in transgenic chloroplasts. For instance, the human serum albumin (hsa) coding sequence fused with 5' UTRs under light was controlled by chloroplast psbA, which boosted protein expression by 500-fold [24].
Although there are shreds of evidence of an increase in protein accumulation due to the structure that protects the foreign protein from degradation. Studies to determine whether the fusion of the C. reinhardtii 16S promoter with its 5'UTR regulatory elements contributed to the high protein synthesis levels. It has been found that larger amounts of luxCt (the marker gene) mRNA at steady-state and, subsequently, higher quantities of mRNA produced via the 16S promoter lead to greater protein accumulation from the atpA 5'UTR [25]. For the determination of complete expression levels for the chimeric constructs, RNA stability did not play any significant role. However, there is a 200-fold higher protein expression recorded having constructed with 5′ UTR of psbA as compared to constructs holding the rbcL 5′ UTR [26]. Prokaryotic messenger RNAs have Shine-Dalgarno (SD) sequences in their 5′ untranslated regions (5′ UTR) that enable translation activation. They function as ribosome-binding sites by recognizing a sequence motif at the 3′ end of the 30S ribosomal subunit's 16S ribosomal RNA [26]. There are pieces of evidence that suggested the translated mRNAs in a few bacteria deficient in the identifiable SD motif representing translation activation with lack of an SD sequence can happen with great efficacy [25]. Stern et al. [27] provided intriguing observations into the procedure of translation activation with a lack of a ribosome-binding site.
However, an In-vivo analysis of the projected RBS efficacy for the tobacco chloroplast atpI gene on translation suggested that for chloroplasts to effectively translate, there must be an element or elements upstream of the putative ribosome binding site [28]. Shine-Dalgarno (SD)-like sequences, which are generally GGAGG, are found in the 5′-untranslated sections of four tobacco chloroplast mRNAs, indicating that functional SD-like sequences are present about 10 nucleotides upstream from the translational initiation codon [28]. It has been claimed that the 5′-UTR is a binding site for mRNA-specific translational activator proteins and immediately directs the ribosomal 30S subunit to the initiation codon since not all SD sequences are conserved on 4–9 nucleotides upstream of the start codon. To manipulate translation initiation efficiency for reverse genetics and transgene expression in plastids, SD sequences generated from bacteriophages have been used. The 3’UTRs are vital in chloroplast mRNAs and it is located immediately downstream of the stop-codon. It contains inverted repeats (IRs) that can form stem-loop structures which aid in the maturation and stability of the transcript. And likewise, hinder 3’ to 5’exonucleases attack and block polyadenylation. However, Ribonuclease has been used to delete polyadenylation sites and protective stem-loop structures, which starts plastid mRNA breakdown [29]. The rps16, rbcL, psbA, and rpl32 3' UTRs are the loci that are typically employed to control foreign genes in plastids.
Marker genes: For precise reliable transformation detection, the chloroplasts and associated cell lines that carry the altered plastomes must contain only homoplasmic copies of the transformed plastomes. As a result, it's essential to fuse the marker gene to a plastid expression system that includes a plastid promoter, a 5' untranslated region (UTR) with a Shine Dalgarno sequence for ribosome binding, and a 3' UTR with cis-acting elements. Herbicide resistance markers like phosphinothricin acetyltransferase (pat/bar gene), Bromoxynil nitrilase (bxn gene), and Antibiotic resistance genes have been extensively used to identify transformants. neo gene and aphA6 are antibiotic resistance that confers kanamycin resistance while the popular antibiotic resistance gene aminoglycoside 3''-adenylyltransferase gene confers resistance to antibacterial. There are public in addition to biosafety concerns on the use of antiseptic or weedicide resistance genes as Specific markers of plant renovation. Therefore, the extinction of the antagonistic bacterial gene from cloned plants is crucial. The antibiotic or herbicide resistance genes avoidance includes; homology-based excision via directly repeated sequences, excision by phage site-specific recombinases, passing co-integration of the specific gene, and co-transformation separation. Staub et al. [30] investigated the bad gene in spinach (Spinacia oleracea), a plant with a particular marker gene for changing chloroplast genomes and they reported 25% chloroplast transformation efficiency in BA than streptomycin. The selection is centered on how the chloroplast site-specific gene-encoding enzyme BADH converts the toxic substance betaine aldehyde, which is only present in chloroplasts, to helpful Gly betaine. Recently, McCabe et al. [31] provided the viability of using dsdA gene as a specific marker gene in plastid and nuclear transformation systems and proposed that dsdA gene can also be used as a marker gene in plastid transformation of monocotyledons.
Application overview of chloroplast transformation
Biotechnology research has greatly explored the unique and significant advantages of chloroplast transformation toward the provision of affordable biopharmaceuticals like vaccines and antibodies. Similarly, the chloroplast genome has been modified to produce valuable enzymes, biomaterials, biofuels, and biomass augmentation as well as the sustainability of food production against factors that might influences production negatively.
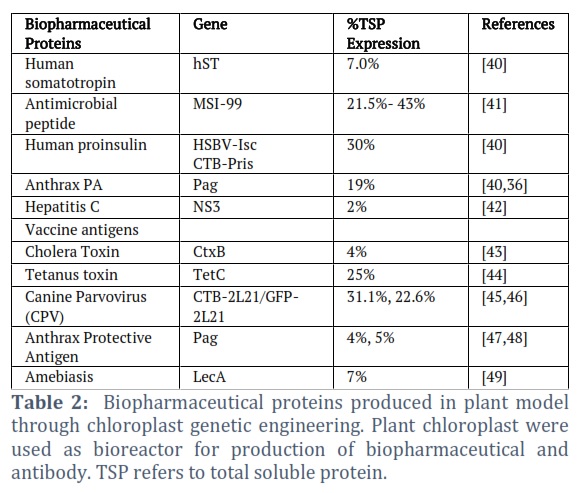
Pharmaceutical vaccine Production: Chloroplast transformation has attracted the pharmaceutical industries due to the DNA of the plastids' high copy number that enables bulk production of vaccine protein. Additionally, it decreases the restrictions based on the costs of the chemically manufactured vaccine's production and purification, thus could be a source of affordable vaccines for the public in developing nations. According to the idea, a plant's chloroplast genome incorporates the targeted genes that produce the bacterial and viral antigens (causing a particular disease) and is secreted as immunogenic proteins in edible tissues. However, only non-glycosylated antigens can be expressed in plastids due to the plastid's lack of a mechanism for glycosylation. There has been a significant advancement in the use of plant chloroplasts in bioreactors because it has been found that plants can significantly increase immunogenicity (Table 2). Arntzen [32] suggested that this newly inserted gene in plants might be replicated without any inherent constraints when they observed that transgenic plants generated HBsAg that was anti-genetically and the same holds for the HBsAg particles produced by recombinant yeast and human serum. Because HBsAg was detected in leaf extracts at levels that correlate with mRNA plenty by enzyme-linked immunoassays utilizing a monoclonal antibody exposed against HBsAg generated from human serum.
LecA, a potential amoebiasis-blocking target, was present in chloroplasts and produces 6.3% of TSP or 2.3 mg LecA/g leaf tissue [33]. Higher amounts of recombinant protein accumulation (more than 300-fold) were achieved compared to nuclear transformation when the secretory human protein somatotropin was expressed in the chloroplast [34]. HIV-1 p24 antigen has been synthesized in the high-biomass tobacco cultivar as well as a higher total soluble protein produced as a result of additional bacterial and viral antigens in different plant plastids [35].
Despite advancements in the plant-based pharmaceutical vaccine, it is not correlated with several clinical trials. However, a recent report on a clinical success in ZMapp—a tool developed by systematic selection of antibody components made by quick transient expression method in tobacco species—against the Ebola virus disease [30]. Currently, adjuvants are co-expressed with antigens to aid in antigen delivery and stimulate the immune response to a particular antigen. Chloroplasts from transgenic organisms are used as bioreactors to synthesize hybrid proteins. Koya et al. [36] evaluated a recombinant SOD produced by Withania somnifera a chloroplast transformation vector harboring a gene (encoding Cu/Zn SOD). They reported that transplastomic plants accumulated recombinant SOD up to 9% of the TSP in leaves and that pure chloroplast-expressed recombinant SOD had an estimated specific activity of 4600 U/mg. These findings demonstrate the possibility of chloroplast transformation for the commercial production of recombinant Superoxide dismutase in plants.
Antibodies bioreactor: Furthermore, the advance in biotechnology has enabled the expression of the antibody in the Chloroplast to ease the constantly increased demand for drugs to recover disorders. Human monoclonal antibodies are large single-chain antibodies used against HSV glycoprotein D and IgA heavy chain proteins imbedded in light chain protein by peptide linkage whose chloroplast was successfully able to express it [37]. Daniell et al. [38] explained the chloroplasts equipped with machinery to assemble complex human antibodies and for the first time, they show that assembled human IgG1 monoclonal antibodies can be produced and purified from chloroplasts of C. reinhardtii. The expression of embedded proteins that have an antibody domain that targets CD22, a B-cell surface epitope, and an enzyme domain from Pseudomonas aeruginosa resulted in the collection of mono and dimeric protein toxin in algal chloroplasts [39]. There are various evidence of monoclonal production in the plant (Table 3); based on the ability of chloroplast machinery for disulfide bond formation that enables large-scale product at less cost.
Application in sustainable food security
Chloroplast transformation has extensively been employed towards the achievement of affordable quantity and nutritious food. There have been reports of various transformations of the chloroplast genome for improvement in various agricultural traits; like herbicide, insecticide resistance, and tolerance deficiency of water and salt.
Chloroplast transformation in Nutritional enhancement: The breakthrough in chloroplast transformation has been explored for the improvement of nutritional biochemical pathways towards the incorporation and enablement of some lacking nutrients in non-green plant parts. In a study to improve carotenoid biosynthesis in tomatoes, the bacterial lycopene beta-cyclase gene triggers the changing of lycopene to beta-carotene and fourfold improvement of pro-vitamin A (fruits element) [50]. Similarly, lycopene beta-cyclase genes from Narcissus pseudonarcissus and Erwinia herbicola expressed in the tomato genome influenced the ability of the plant enzyme and efficiently converted lycopene into provitamin A (beta-carotene). Iordachescu and Imai [51] demonstrated more lycopene accumulation of xanthophyll cycle pigments. Consequently, it reduced alpha-branch xanthophyll lutein in transplastomic tomato plants’ green leaves while lycopene changed to beta-carotene with the provitamin A level reaching 1 mg/per dry weight in fruits. High-level expression of γ-TMT was reported to assess the conversion of most of the γ-tocopherol to α-tocopherol in transplastomic seeds (~10 fold higher) it enhances salt and heavy metal stresses by lowering ROS, ion leakage, and lipid peroxidation [51].
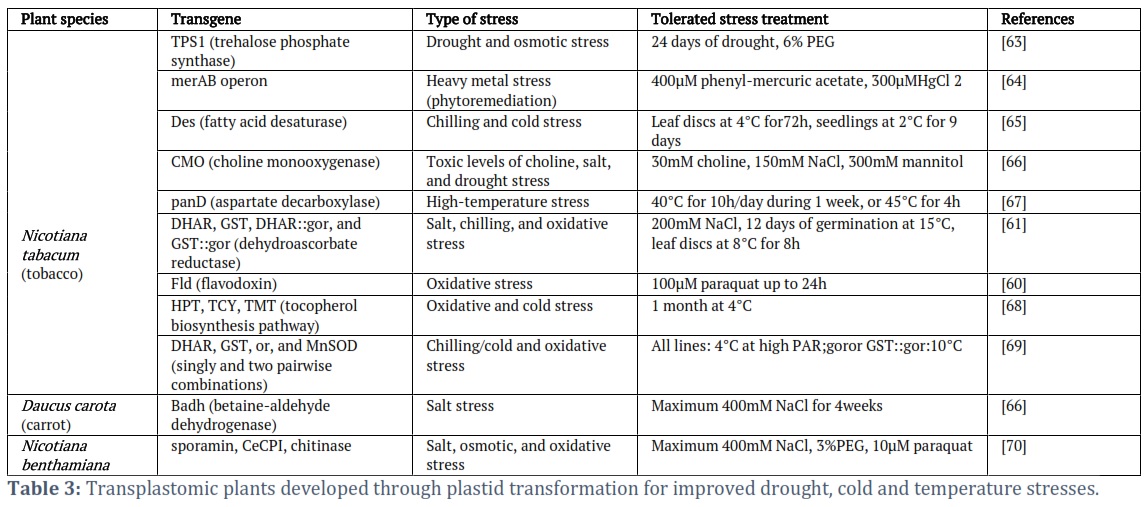
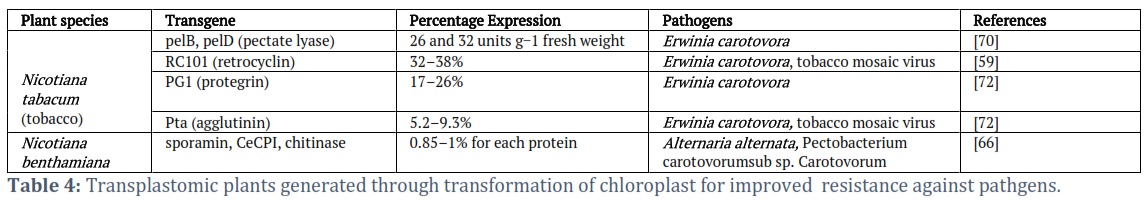
Chloroplast transformation in Agricultural stress management: Drought, salt, temperature (cold, chilling, or heat) stress which constitute abiotic stress, and biotic stress like disease (pathogen) and insect attack are the major factors that negatively influenced the availability of secure agricultural output. Biotechnology advancement in chloroplast transformation has proven to be a reliable technique for agricultural improvement in the present age of climatic change (Table 4); and the exponential increase of human population growth.
- Temperature stress management: Scientist over the years has studied the various function of the gene involved in environmental stress. The potential for genetic modification of the unsaturated fatty acids in plant lipids has ramifications for higher plants' capacity to withstand stress. Changed leaf fatty acid profiles of transplastomic plants result raise unsaturation levels in plant seeds and leaves and proposed that transplastomic would be more tolerant to cold. When the chloroplast genome of tobacco expressed Escherichia coli L-aspartate-alpha-decarboxylase (AspDC), a gene that catalyzes the decarboxylation of L-aspartate to produce beta-alanine and carbon dioxide. Badawi et al. [52] demonstrated E. coli panD expression improves the heat tolerance of photosynthesis and biomass production during high-temperature stress. Oxidative stress is a harmful factor for plants that are in direct contact with environmental stresses. Fang-Zheng et al. [53] improve the transgenic potato's resistance to oxidative and thermal stress by exhibiting superoxide dismutase and ascorbate peroxidase.
- Drought and salt stress management: Salt stress is another major and still progressively disorder in the agriculture sector. Saline soil affects 7% of the total area and 5% of the land utilized for agriculture, respectively. Consequently, poses a cost-effective problem due to stunted growth, slow development, and lesser output of crops on such soils. Several osmoprotectants, e.g., the sugar trehalose [54] and betaines [55], boost the cell’s ability to purge ROS and preserve macromolecules to bolster its ability to withstand salt, cold, and/or drought. Successful recombination of transplastomic plants by somatic cells and transformation of carrot cells. These plants express BADH in the presence of high NaCl concentrations, and transgenic carrot cell cultures of these plants developed seven times faster and accumulated 50 to 54 times more betaine than untransformed cells cultured in liquid media with 100 mM NaCl [55]. Zhang et al. [56] examine the significance of excessive Arabidopsis thaliana cytosolic APX synthesis in tobacco chloroplasts for protecting the plant from oxidative stressors brought on by Na2SO3 salt stress (NaCl). Fouad and Altpeter [48] evaluated the function of the vital antioxidant enzyme manganese superoxide dismutase (MnSOD). They suggest that since rice exhibits the capacity to scavenge reactive oxygen species (ROS) damage, it may contribute to the plant’s ability to withstand drought. It has been demonstrated that transgenic tobacco plants that overexpress the BvCMO gene from beet, a halophyte plant that naturally generates GlyBet, accumulate GlyBet and exhibit increased resistance to salt and drought stress. It increases the rate and yield of photosynthesis when exposed to salt stress (150 mM NaCl) [57]. Recently, Ceccoli et al. [58] reported that the expression of ArDH genes in the chloroplast metabolic pathway imparts tolerance to NaCl on tobacco chloroplasts (up to 400 mM).
- Improving tolerance towards biotic stressors: Biotic stress is another important factor that is currently affecting food sustainability. If not controlled it could result in serious endemic disease problems and limit the world's food availability. Accumulation of toxic compounds from chemical use in diseases and pest control necessitates alternative control measures.
- Pathogen resistance transplastomic plants: Plastid transformation has been proven to be an essential strategy to improve disease resistance to plant pathogenic bacteria and fungi because many of the target proteins store in a single region and release during the hypersensitive reaction [59]. There are several reports on transplastomic plants that are resistant to disease pathogens (Table 5). Insect and plant pathogen resistance is widespread in transgenic plants that express the agglutinin gene from Pinellia ternate [59]. Cry9Aa2 gene from Bacillus thuringiensis was inserted into tobacco plant plastids to withstand the potato tuber moth [60]. The transgenic tomato’s CeCPI and chitinase genes are expressed [61] and CeCPI and sporamin in tobacco showed resistance to insects and phytopathogens. The expression of several defense genes producing protease inhibitors and chitinase was later shown to be responsible for Nicotiana benthamiana's high level of broad-spectrum resistance to pests, diseases, and abiotic stressors [62].
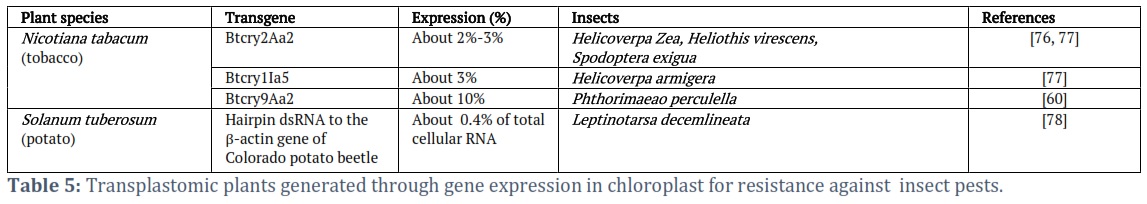
- Transplastomic plants with enhanced insect resistance: Transplastomic plants have been illustrated to show resistance against larvae of H. armigera, irrespective of the development stage. Reddy et al. [73] elaborated that the Cry1Ia5 protein collects up to 3% of the TSP in tissue which is 300 times more when likened to the express same protein in nuclear-transformed plants. Various transgenic plants have been commercialized and extensively grown expressing Bt crystal proteins (Cry1Ab in maize and Cry1Ac in cotton). Bacillus thuringiensis (Bt) crystal proteins are regarded as secure biological pesticides. However, there is serious contention on the transfer of such toxic protein to non-potential target vial pollen. Chloroplast transformation has been greatly investigated to combat this limitation since the transformation is not done in the nucleus and chloroplast is maternally transferred to the next generation. Various evidence has proved that the chloroplast genome could be targeted for high expression toward pest insect management and control. Chakrabarti et al. [60] reported a chance to produce insect-resistant transplastomic tobacco by up-regulation of insect pathogen defense mechanisms. Must recently, scientists are now employing RNAi technology in the transplastomic transformation toward an insect-specific and environmentally friendly control measure. This technology affects only the specific target insect, thus beneficial insects are not affected and do not produce any toxic protein in the plant like BT protein. Zhang et al. [74] reported that the amount of foreign protein, accumulated at the greatest level in mature leaves was 45.3% of the total soluble protein and 100% mortality of armyworms after the consumption of transgenic leaves. Three vital genes were targeted in Helicoverpa armigera by expression of the corresponding gene (V‐ATPase and Chi, P450 (CYP6AE14)) dsRNAs in the chloroplasts of tobacco. It was observed that increased transcript abundance greatly inhibited target gene transcription and slowed larval growth in the insect midgut. Consequently, the net weight of larvae, growth rate, and pupation rate were all dramatically reduced.
In a trial to observe whether the plant's inherent RNAi machinery can reduce this pesticide activity, dsRNA was diced into siRNAs. Sanz-Barrio et al. [75] identified full-length hpRNAs stored in transplastomic lines of N. benthamiana, which provided substantial protection against H. armigera herbivory. In contrast, hpRNAs of nuclear-transformed plants were converted into siRNAs, which provided a more reduced anti-feeding effect. According to this result, the chloroplast has little to no RNAi machinery or activity, and the hpRNAs produced there, do not transit the cytoplasm. It has been observed that the midgut tissue of beetles feeding on transplastomic potato leaves had siRNAs specific to the targeted genes and mRNA transcript levels of the targeted genes were lowered upon Colorado potato beetle feeding on transplastomic potato plants.
Figures & Tables
Over the years, the chloroplast genome had been an eye-catching target to improve photosynthesis and crop performance. Apart from photosynthesis pathways found in the chloroplast, there are other biosynthetic pathways with a low amount of regulatory proteins like NADPH-dependent thioredoxin reductase (NTRC), ferredoxin-thioredoxin reductase (FTR), and Thioredoxins (TRXs). Such protein overexpression may be investigated for plant development. For instance, increasing the activity of these TRXs may promote chloroplast biosynthetic processes and improve plant development [67]. Higher expression of caffeic acid O-methyltransferase I transgenic rice chloroplast was reported to enhance melatonin synthesis in chloroplasts via 5-methoxytryptamine pathway, indicating improved seedling development [1]. Additionally, it has been found that retrograde signals produced by chloroplasts can change the expression of nuclear genes [2]. To design a stress tolerance plant, this particular feature could be investigated by using targeted genome editing tools l like meganucleases (MNs), transcription activator-like effector nucleases (TALENs), zinc finger nucleases (ZFNs), Clustered Regularly Interspaced Short Palindromic Repeat (CRISPR). Targeted genome editing has been investigated to be a potent genetic tool for studying gene function or for altering genomes by introducing desired genes and fixing faulty genes. However, the use of such targeted editing tools to modify a multitude of known pathways in the chloroplast genome is either limited or absent. To our knowledge, no reports exist of the CRISPR/Cas9 system being used to induce stable targeted gene changes in the chloroplast genome of either the model species or any staple plant. The CRISPR/Cas9 technology developed from a type II bacterial immune system [79]. Since it has been evident that the endosymbiotic process gave rise to chloroplasts in which a type II bacterial immune system gave rise to a free-living cyanobacterium and the CRISPR/Cas9 technology. Considering the evolutionary link between both, we envisage that optimization of CRISPR/Cas9 technology in chloroplast would be a game changer in transformation of chloroplast and its usage in various applications.
Conflict of Interest
The authors declare no conflict of interest.
Acknowledgement
The author acknowledges the support from International Foundation for Science grant (No C/6257-1).
BT: Conceptulization and writing-Reviewing and editing, IY: Investigation and writing, OSA: literature and data curation, RK: Writing, AK: Resources and literature.
![]() References
References
- Martin W, Rujan T, Richly E, Hansen A, Cornelsen S, et al.. Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proceedings of the National Academy of Sciences, (2002); 99 (19): 12246–12251.
- del Campo E. Post-transcriptional control of chloroplast gene expression. Gene Regulation and Systems Biology, (2009); 3: 31–47.
- Chumley TW, Palmer JD, Mower JP, Fourcade HM, Calie PJ, Boore JL, Jansen RK. The complete chloroplast genome sequence of Pelargonium× hortorum: organization and evolution of the largest and most highly rearranged chloroplast genome of land plants. Molecular biology and evolution, (2006); 23(11): 2175-90.
- Gao L, SU YJ, Wang T. Plastid genome sequencing, comparative genomics, and phylogenomics: current status and prospects. Journal of Systematics and Evolution, (2010); 48(2): 77-93.
- Wambugu PW, Brozynska M, Furtado A, Waters DL, Henry RJ. Relationships of wild and domesticated rices (Oryza AA genome species) based upon whole chloroplast genome sequences. Scientific reports, (2015); 5(1): 13957.
- Chacón S MI, Pickersgill B, Debouck DG. Domestication patterns in common bean (Phaseolus vulgaris L.) and the origin of the Mesoamerican and Andean cultivated races. Theoretical and Applied Genetics, (2005); 110: 432-44.
- Svab Z, Maliga PA. High-frequency plastid transformation in tobacco by selection for a chimeric aadA gene. Proceedings of the National Academy of Sciences, (1993); 90(3): 913-7.
- Ahmad A, O Pereira E, J Conley A, S Richman A, Menassa R. Green biofactories: recombinant protein production in plants. Recent patents on biotechnology, (2010); 4(3): 242-59.
- Ravi V, Khurana JP, Tyagi AK, Khurana PJ. An update on chloroplast genomes. Plant Systematics and Evolution, (2008); 271: 101-22.
- Kavanagh TA, Thanh ND, Lao NT, McGrath N, Peter SO, Horváth EM, Dix PJ, Medgyesy P. Homeologous plastid DNA transformation in tobacco is mediated by multiple recombination events. Genetics, (1999); 152(3): 1111-22.
- Guda C, Lee SB, Daniell H. Stable expression of a biodegradable protein-based polymer in tobacco chloroplasts. Plant Cell Reports, (2000); 19: 257-62.
- Verma D, Daniell H. Chloroplast vector systems for biotechnology applications. Plant physiology, (2007); 145(4): 1129-43.
- De Block M, Schell J, Van Montagu M. Chloroplast transformation by Agrobacterium tumefaciens. The EMBO journal, (1985); 4(6): 1367-72.
- Chen Q, Lai H. Plant-derived virus-like particles as vaccines. Human vaccines & immunotherapeutics, (2013); 9(1): 26-49.
- Cañizares MC, Nicholson L, Lomonossoff GP. Use of viral vectors for vaccine production in plants. Immunology and Cell Biology, (2005); 83(3): 263-70.
- Hedtke B, Legen J, Weihe A, Herrmann RG, Börner T. Six active phage‐type RNA polymerase genes in Nicotiana tabacum. The Plant Journal, (2002);30(6):625-37.
- Liere K, Weihe A, Börner T. The transcription machineries of plant mitochondria and chloroplasts: composition, function, and regulation. Journal of plant physiology, (2011);168(12):1345-60.
- Stoppel R, Meurer J. The cutting crew–ribonucleases are key players in the control of plastid gene expression. Journal of experimental botany, (2012);63(4):1663-73.
- Fernández‐San Millán A, Mingo‐Castel A, Miller M, Daniell H. A chloroplast transgenic approach to hyper‐express and purify Human Serum Albumin, a protein highly susceptible to proteolytic degradation. Plant biotechnology journal, (2003);1(2):71-9.
- Kuroda H, Maliga P. Sequences downstream of the translation initiation codon are important determinants of translation efficiency in chloroplasts. Plant physiology, (2001);125(1):430-6.
- Rasala BA, Muto M, Sullivan J, Mayfield SP. Improved heterologous protein expression in the chloroplast of Chlamydomonas reinhardtii through promoter and 5′ untranslated region optimization. Plant biotechnology journal, (2011);9(6):674-83.
- Eibl C, Zou Z, Beck A, Kim M, Mullet J, Koop HU. In vivo analysis of plastid psbA, rbcL and rpl32 UTR elements by chloroplast transformation: tobacco plastid gene expression is controlled by modulation of transcript levels and translation efficiency. The Plant Journal, (1999); 19(3): 333-45.
- Skorski P, Leroy P, Fayet O, Dreyfus M, Hermann-Le Denmat S. The highly efficient translation initiation region from the Escherichia coli rpsA gene lacks a Shine-Dalgarno element. Journal of bacteriology, (2006); 188(17):6277-85.
- Scharff LB, Childs L, Walther D, Bock R. Local absence of secondary structure permits translation of mRNAs that lack ribosome-binding sites. PLoS genetics, (2011); 7(6): e1002155.
- Kuroda H, Suzuki H, Kusumegi T, Hirose T, Yukawa Y, Sugiura M. Translation of psbC mRNAs starts from the downstream GUG, not the upstream AUG, and requires the extended Shine–Dalgarno sequence in tobacco chloroplasts. Plant and cell physiology, (2007); 48(9):1374-8.
- Zhou F, Badillo‐Corona JA, Karcher D, Gonzalez‐Rabade N, Piepenburg K, Borchers AM, Maloney AP, Kavanagh TA, Gray JC, Bock R. High‐level expression of human immunodeficiency virus antigens from the tobacco and tomato plastid genomes. Plant biotechnology journal, (2008); 6(9): 897-913.
- Stern DB, Goldschmidt-Clermont M, Hanson MR. Chloroplast RNA metabolism. Annual review of plant biology, (2010); 61: 125-55.
- Dufourmantel N, Dubald M, Matringe M, Canard H, Garcon F, Job C, Kay E, Wisniewski JP, Ferullo JM, Pelissier B, Sailland A. Generation and characterization of soybean and marker‐free tobacco plastid transformants over‐expressing a bacterial 4‐hydroxyphenylpyruvate dioxygenase which provides strong herbicide tolerance. Plant biotechnology journal, (2007); 5(1):118-33.
- Ma JK, Drake PM, Christou P. The production of recombinant pharmaceutical proteins in plants. Nature reviews genetics, (2003); 4(10): 794-805.
- Staub JM, Garcia B, Graves J, Hajdukiewicz PT, Hunter P, Nehra N, Paradkar V, Schlittler M, Carroll JA, Spatola L, Ward D. High-yield production of a human therapeutic protein in tobacco chloroplasts. Nature biotechnology, (2000); 18(3): 333-8.
- McCabe MS, Klaas M, Gonzalez‐Rabade N, Poage M, Badillo‐Corona JA, Zhou F, Karcher D, Bock R, Gray JC, Dix PJ. Plastid transformation of high‐biomass tobacco variety Maryland Mammoth for production of human immunodeficiency virus type 1 (HIV‐1) p24 antigen. Plant biotechnology journal, (2008) ;6(9): 914-29.
- Arntzen C. Plant‐made pharmaceuticals: from ‘Edible Vaccines’ to Ebola therapeutics. Plant biotechnology journal, (2015); 13(8):1013.
- Mayfield SP, Franklin SE, Lerner RA. Expression and assembly of a fully active antibody in algae. Proceedings of the National Academy of Sciences, (2003); 100(2):438-42.
- Tran M, Zhou B, Pettersson PL, Gonzalez MJ, Mayfield SP. Synthesis and assembly of a full‐length human monoclonal antibody in algal chloroplasts. Biotechnology and bioengineering, (2009);104(4):663-73.
- Tran M, Van C, Barrera DJ, Pettersson PL, Peinado CD, Bui J, Mayfield SP. Production of unique immunotoxin cancer therapeutics in algal chloroplasts. Proceedings of the National Academy of Sciences, (2013); 110(1):E15-22.
- Koya V, Moayeri M, Leppla SH, Daniell H. Plant-based vaccine: mice immunized with chloroplast-derived anthrax protective antigen survive anthrax lethal toxin challenge. Infection and immunity, (2005); 73(12):8266-74.
- Watson J, Koya V, Leppla SH, Daniell H. Expression of Bacillus anthracis protective antigen in transgenic chloroplasts of tobacco, a non-food/feed crop. Vaccine, (2004); 22(31-32):4374-84.
- Daniell H, Chebolu S, Kumar S, Singleton M, Falconer R. Chloroplast-derived vaccine antigens and other therapeutic proteins. Vaccine, (2005); 23(15): 1779-83.
- Daniell H, Lee SB, Panchal T, Wiebe PO. Expression of the native cholera toxin B subunit gene and assembly as functional oligomers in transgenic tobacco chloroplasts. Journal of molecular biology, (2001); 311(5):1001-9.
- Tregoning JS, Nixon P, Kuroda H, Svab Z, Clare S, Bowe F, Fairweather N, Ytterberg J, Wijk KJ, Dougan G, Maliga P. Expression of tetanus toxin fragment C in tobacco chloroplasts. Nucleic acids research, (2003);31(4):1174-9.
- Molina A, Hervás‐Stubbs S, Daniell H, Mingo‐Castel AM, Veramendi J. High‐yield expression of a viral peptide animal vaccine in transgenic tobacco chloroplasts. Plant biotechnology journal, (2004);2(2):141-53.
- Chebolu S, Daniell H. Stable expression of Gal/GalNAc lectin of Entamoeba histolytica in transgenic chloroplasts and immunogenicity in mice towards vaccine development for amoebiasis. Plant biotechnology journal, (2007);5(2):230-9.
- Wurbs D, Ruf S, Bock R. Contained metabolic engineering in tomatoes by expression of carotenoid biosynthesis genes from the plastid genome. The Plant Journal, (2007); 49(2): 276-88.
- Apel W, Bock R. Enhancement of carotenoid biosynthesis in transplastomic tomatoes by induced lycopene-to-provitamin A conversion. Plant Physiology, (2009); 151(1): 59-66.
- Craig W, Lenzi P, Scotti N, De Palma M, Saggese P, Carbone V, McGrath Curran N, Magee AM, Medgyesy P, Kavanagh TA, Dix PJ. Transplastomic tobacco plants expressing a fatty acid desaturase gene exhibit altered fatty acid profiles and improved cold tolerance. Transgenic research, (2008); 17: 769-82.
- Jin S, Daniell H. Expression of γ‐tocopherol methyltransferase in chloroplasts results in massive proliferation of the inner envelope membrane and decreases susceptibility to salt and metal‐induced oxidative stresses by reducing reactive oxygen species. Plant biotechnology journal, (2014); 12(9):1274-85.
- Tang L, Kwon SY, Kim SH, Kim JS, Choi JS, Cho KY, Sung CK, Kwak SS, Lee HS. Enhanced tolerance of transgenic potato plants expressing both superoxide dismutase and ascorbate peroxidase in chloroplasts against oxidative stress and high temperature. Plant Cell Reports, (2006);25:1380-6.
- Fouad WM, Altpeter F. Transplastomic expression of bacterial L-aspartate-α-decarboxylase enhances photosynthesis and biomass production in response to high temperature stress. Transgenic research, (2009);18:707-18.
- Munns R, Tester M. Mechanisms of salinity tolerance. Mechanisms of Salinity Tolerance. Annual Review of Plant Biology, (2008); 59: 651–81.
- Karim S, Aronsson H, Ericson H, Pirhonen M, Leyman B, Welin B, Mäntylä E, Palva ET, Van Dijck P, Holmström KO. Improved drought tolerance without undesired side effects in transgenic plants producing trehalose. Plant Molecular Biology, (2007); 64:371-86.
- Iordachescu M, Imai R. Trehalose biosynthesis in response to abiotic stresses. Journal of integrative plant biology, (2008); 50(10):1223-9.
- Badawi GH, Kawano N, Yamauchi Y, Shimada E, Sasaki R, Kubo A, Tanaka K. Over‐expression of ascorbate peroxidase in tobacco chloroplasts enhances the tolerance to salt stress and water deficit. Physiologia Plantarum, (2004); 121(2):231-8.
- Wang FZ, Wang QB, Kwon SY, Kwak SS, Su WA. Enhanced drought tolerance of transgenic rice plants expressing a pea manganese superoxide dismutase. Journal of plant physiology, (2005); 162(4): 465-72.
- Khan MS, Kanwal B, Nazir S. Metabolic engineering of the chloroplast genome reveals that the yeast ArDH gene confers enhanced tolerance to salinity and drought in plants. Frontiers in plant science, (2015); 6: 725.
- Ruiz ON, Hussein HS, Terry N, Daniell H. Phytoremediation of organomercurial compounds via chloroplast genetic engineering. Plant Physiology, (2003); 132(3):1344-52.
- Zhang J, Tan W, Yang XH, Zhang HX. Plastid-expressed choline monooxygenase gene improves salt and drought tolerance through accumulation of glycine betaine in tobacco. Plant cell reports, (2008); 27: 1113-24.
- Le Martret B, Poage M, Shiel K, Nugent GD, Dix PJ. Tobacco chloroplast transformants expressing genes encoding dehydroascorbate reductase, glutathione reductase, and glutathione‐S‐transferase, exhibit altered anti‐oxidant metabolism and improved abiotic stress tolerance. Plant Biotechnology Journal, (2011); 9(6): 661-73.
- Ceccoli RD, Blanco NE, Segretin ME, Melzer M, Hanke GT, Scheibe R, Hajirezaei MR, Bravo-Almonacid FF, Carrillo N. Flavodoxin displays dose-dependent effects on photosynthesis and stress tolerance when expressed in transgenic tobacco plants. Planta, (2012); 236:1447-58.
- Lee SB, Li B, Jin S, Daniell H. Expression and characterization of antimicrobial peptides Retrocyclin‐101 and Protegrin‐1 in chloroplasts to control viral and bacterial infections. Plant biotechnology journal, (2011); 9(1):100-15.
- Chakrabarti SK, Lutz KA, Lertwiriyawong B, Svab Z, Maliga P. Expression of the cry9Aa2 Bt gene in tobacco chloroplasts confers resistance to potato tuber moth. Transgenic research, (2006); 15: 481-8.
- Jin S, Zhang X, Daniell H. Pinellia ternata agglutinin expression in chloroplasts confers broad spectrum resistance against aphid, whitefly, Lepidopteran insects, bacterial and viral pathogens. Plant biotechnology journal, (2012); 10(3): 313-27.
- Jin S, Kanagaraj A, Verma D, Lange T, Daniell H. Release of hormones from conjugates: chloroplast expression of β-glucosidase results in elevated phytohormone levels associated with significant increase in biomass and protection from aphids or whiteflies conferred by sucrose esters. Plant physiology, (2011); 155(1): 222-35.
- Lu Y, Rijzaani H, Karcher D, Ruf S, Bock R. Efficient metabolic pathway engineering in transgenic tobacco and tomato plastids with synthetic multigene operons. Proceedings of the National Academy of Sciences, (2013); 110(8): E623-32.
- Grant OM, Brennan DP, Mellisho Salas CD, Dix PJ. Impact of enhanced capacity to scavenge reactive oxygen species on cold tolerance of tobacco. International Journal of Plant Sciences, (2014); 175(5):544-54.
- Kumar S, Dhingra A, Daniell H. Plastid-expressed betaine aldehyde dehydrogenase gene in carrot cultured cells, roots, and leaves confers enhanced salt tolerance. Plant Physiology, (2004); 136(1): 2843-54.
- Chen PJ, Senthilkumar R, Jane WN, He Y, Tian Z, Yeh KW. Transplastomic Nicotiana benthamiana plants expressing multiple defence genes encoding protease inhibitors and chitinase display broad‐spectrum resistance against insects, pathogens and abiotic stresses. Plant Biotechnology Journal, (2014); 12(4): 503-15.
- Wani SH, Sah SK, Sági L, Solymosi K. Transplastomic plants for innovations in agriculture. A review. Agronomy for sustainable development, (2015); 35: 1391-430.
- Chan YL, Cai D, Taylor PW, Chan MT, Yeh KW. Adverse effect of the chitinolytic enzyme PjCHI‐1 in transgenic tomato on egg mass production and embryonic development of Meloidogyne incognita. Plant pathology, (2010); 59(5): 922-30.
- Senthilkumar R, Cheng CP, Yeh KW. Genetically pyramiding protease‐inhibitor genes for dual broad‐spectrum resistance against insect and phytopathogens in transgenic tobacco. Plant biotechnology journal, (2010); 8(1):65-75.
- Verma D, Kanagaraj A, Jin S, Singh ND, Kolattukudy PE, Daniell H. Chloroplast‐derived enzyme cocktails hydrolyse lignocellulosic biomass and release fermentable sugars. Plant biotechnology journal, (2010); 8(3): 332-50.
- Cosa BD, Moar W, Lee SB, Miller M, Daniell H. Overexpression of the Bt cry2Aa2 operon in chloroplasts leads to formation of insecticidal crystals. Nature biotechnology, (2001); 19(1):71-4.
- Yu Y, Yu PC, Chang WJ, Yu K, Lin CS. Plastid transformation: how does it work? Can it be applied to crops? What can it offer?. International Journal of Molecular Sciences, (2020); 21(14): 4854.
- Reddy VS, Leelavathi S, Selvapandiyan A, Raman R, Giovanni F, Shukla V, Bhatnagar RK. Analysis of chloroplast transformed tobacco plants with cry 1Ia5 under rice psbA transcriptional elements reveal high level expression of Bt toxin without imposing yield penalty and stable inheritance of transplastome. Molecular Breeding, (2002); 9: 259-69.
- Zhang J, Khan SA, Hasse C, Ruf S, Heckel DG, Bock R. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science, (2015); 347(6225): 991-4.
- Sanz‐Barrio R, Corral‐Martinez P, Ancin M, Segui‐Simarro JM, Farran I. Overexpression of plastidial thioredoxin f leads to enhanced starch accumulation in tobacco leaves. Plant Biotechnology Journal, (2013); 11(5):618-27.
- Adeyinka OS, Tabassum B, Koloko BL, Ogungbe IV. Enhancing the quality of staple food crops through CRISPR/Cas-mediated site-directed mutagenesis. Planta, (2023); 257(4): 78.
- Jin S, Singh ND, Li L, Zhang X, Daniell H. Engineered chloroplast dsRNA silences cytochrome p450 monooxygenase, V‐ATPase and chitin synthase genes in the insect gut and disrupts Helicoverpa armigera larval development and pupation. Plant biotechnology journal, (2015); 13(3): 435-46.
- He G. Engineering chloroplasts for insect pest control. Proceedings of the National Academy of Sciences, (2022);119(22):e2205125119.
- Kadoić Balaško M, Mikac KM, Bažok R, Lemic D. Modern techniques in Colorado potato beetle (Leptinotarsa decemlineata Say) control and resistance management: history review and future perspectives. Insects, (2020);11(9):581.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


