

Full Length Research Article
Single Nucleotide Polymorphisms in HIF-1A, VEGFa, and VHL Genes: A Comparative Study between Sea Level and High-Altitude Residents in West Saudi Arabia
Siraj B. Alharthi1, 3*, Ijtihed Kilani2, Abdullah Alsoliman3, Romar P. Pascual2, Mohamed H. Zainy1, Ahmed S Albariqi4, Yahya M. Alzahrani4, Ahmed Alshehri5, Ahmed M. El-Shehawi6, Hashim M. Borayk7, Mohamed Morsi M. Ahmed1, 8
Adv. life sci., vol. 11, no. 1, pp. 153-161, February 2024
*– Corresponding Author: Siraj B. Alharthi (sirajj1411@gmail.com)
Authors' Affiliations
2. Science Department, Shorouq Al Mamlakah International School · Taif – Saudi Arabia
3. Molecular Diagnostic Unit, Al Hada Armed Forces Hospital · Taif – Saudi Arabia
4. Genetics Department, Al Hada Armed Forces Hospital · Taif – Saudi Arabia
5. Genetics Department, King Faisal Medical Complex · Taif – Saudi Arabia
6. Department of Biotechnology, College of Science, Taif University · Taif – Saudi Arabia
7. Medical Laboratory, King Fahad Armed Forces Hospital · Jeddah – Saudi Arabia
8. Department of Nucleic Acids Research, Genetic Engineering and Biotechnology, Research Institute (GEBRI) · Alexandria – Egypt
[Date Received: 04/10/2023; Date Revised: 25/01/2024; Date Published: 25/02/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: This comprehensive molecular study investigates the genetic variations in individuals residing at different altitudes in West Saudi Arabia, specifically contrasting sea-level residents in Jeddah (SL) with high-altitude dwellers in Taif (HA). The study focuses on the significant role of genetic adaptations in response to environmental challenges, particularly hypoxia. Understanding these genetic differences according to geographical location is crucial for multiple reasons. It not only sheds light on the adaptive mechanisms that humans develop in response to environmental stressors like reduced oxygen levels but also has broader implications for medical, anthropological, and evolutionary studies. By exploring how distinct populations adapt to their environments, this research provides valuable insights into the complex interplay between genetics, health, and disease, highlighting the importance of considering genetic diversity in medical and genetic research.
Methods: The research involved amplifying and sequencing three pivotal genes associated with hypoxia adaptation: Hif-1a, VEGFa, and VHL. Advanced genomic techniques were utilized to analyze samples from 22 volunteers, 10 from the sea level region of Jeddah and 12 from the high-altitude region of Taif. The study aimed to identify and analyze single nucleotide polymorphisms (SNPs) in these genes and their implications in altitude adaptation.
Results: Distinct SNPs were identified in the Hif-1a, VEGFa, and VHL genes, with high-altitude residents displaying more pronounced variations. The phylogenetic analysis demonstrated clear genetic clustering based on altitude, highlighting the molecular adaptations induced by altitude variations. The study reveals a complex interplay between genetics and environmental conditions, significantly contributing to our understanding of human adaptation to high altitudes.
Conclusion: This research underscores the profound impact of altitude on human genetics. The findings provide critical insights into how genetic variations facilitate adaptation to challenging environmental conditions, such as hypoxia, encountered at high altitudes. These insights have broad implications, potentially informing medical research related to altitude sickness and other altitude-related health issues.
Keywords: High-Altitude Adaptation; Single Nucleotide Polymorphisms (SNPs); Hif-1a Gene; VEGFa Gene; VHL Gene; Phylogenetic Analysis; Saudi Arabian Populations
Introduction![]()
Organisms living at high altitude (HA) are exposed to a stressful environment because of hypoxia (low oxygen pressure), low temperature, and higher radiation. More than 140 million individuals live at high HA around the World [1, 2]. Sudden change in altitude causes different negative impacts on organs including physiological, biochemical, hematological, and molecular changes to adapt to the extreme environmental conditions [1].
Elevation above sea level is defined as high altitude (HA) (2000 to 4000 m), very high altitude (4000 to 5500 m), and extremely high altitude (>5500 m). These altitudes affect the normal physiology and health due to the low partial pressure of oxygen. Hypoxia develops as a result of this low oxygen pressure [3, 4], therefore the body starts to adapt to the HA conditions through several physiological and molecular strategies. The blood pressure is increased to provide more oxygen to the tissues [5]. On the other hand, fast change in blood flow causes fluid leakage from the capillaries leading to serious conditions including high-altitude pulmonary edema (HAPE) and high-altitude cerebral edema (HACE) [6]. Also, cells can sense hypoxia by the oxygen sensor prolyl hydroxylases (PHDs) [5] which activates the transcription factor hypoxia-inducible factor-1 (HIF-1) [7]. The HIF-1 can relay the hypoxia signal and activate the hypoxia adaptive responses throughout the body [5]. Living at high altitude has various different effects on living systems including hypoxia [8], change in transcriptome [9], and changes in gut microbiota (metagenome) of microorganisms in the gut of living organisms.
HA-associated hypoxia has been considered as one of the associated factors in developing IBD (Crohn’s disease and ulcerative colitis) and changes in gut microbiota (dysbiosis) [10]. Low air pressure at HA (hypoxia) lowers the oxygen pressure which affects different organs including lungs, brain, heart, and blood. This causes a wide range of disorders such as HAPE, acute mountain sickness, and HACE [11, 12, 13, 14, 2].
Investigating the genetic differences between human populations from varied geographical locations offers invaluable insights into our evolutionary history and adaptive mechanisms. The significance of this study lies in its examination of specific genetic markers – HIF-1A, VEGFa, and VHL genes – across populations residing at different altitudes. By comparing these genetic variations between sea level and high-altitude populations, this research aims to unravel the intricate ways in which human genetics adapt to environmental pressures, such as varied oxygen levels. Understanding these adaptations is not only crucial for comprehending human evolutionary biology but also has potential implications in medical and genetic research, especially in areas related to altitude sickness, cardiovascular health, and metabolic adaptations [11].
Twenty-two volunteers contributed to this study; 10 from Jeddah as a sea level region and 12 from Taif as a high-altitude region. Volunteers had been chosen as old and original dwellers from each region. Table (1) summarizes the basic information of the 22 volunteers for samples used in this study. A consent was signed by each individual volunteer before contribution in the study. Blood samples (2 ml) were collected from each individual and used in DNA isolation.
Adaptation with life at HA requires rapid change in gene expression and genetic changes in rapidly evolving genes which contribute to the phenotype changes of animals living at HA. Many genomic studies have reported the association of genetic variants to life at HA, especially in the Tibetan native populations [15, 16, 17]. Among the prominent variations is the SNPs of endothelial PAS domain protein 1 (EPAS1) [18]. EPAS1 is an oxygen sensor that regulates hypoxia-inducible transcription factor pathways known as hypoxia-inducible factor-2α (HIF-2α). It plays a vital role in adaptation to HA and is also correlated with decreased hemoglobin levels [15]. SNPs in the EPAS1 were positively correlated with adaptation of the Tibetan to hypoxia at HA. Also, it was reported that three SNPs (rs6756667, rs13419896 and rs4953354) contributed to the physiological effects of acute mountain sickness (AMS) and hypoxia-related disorders in humans [15]. Novel genetic variants in the angiotensin converting enzyme (ACE) and adenosine monophosphate deaminase 1(AMPD1) enhanced various parameters of mountaineers including heart rate, oxygen uptake, and blood pressure. This performance was reported in genotypes carrying variant alleles ACTN3 XX and AMPD1 TT [21]. Also, protein variants in the EGLN1 (rs186996510, rs12097901) were correlated to adaptation of Andean population living at HA [19]. Recent variant of EPAS1 was correlated to susceptibility to acute mountain sickness in Han Chinese human subjects [4].
Several studies showed that high altitude is linked to SNPs in some genes [3]. SNPs were detected in coding, noncoding, exons, and introns, with high frequency in the intergenic regions of the genome.
Taif region is located about 1700-2500 m above sea level and represents the resort for National and International tourists. The impact of high altitude on immunity, SNPs frequency, and global gene expression in the Taif region have not been investigated. Therefore, the main objective of the current study is to investigate the impact of high altitude (Taif region) on the immunological and genetic parameters in human subjects.
Methods![]()
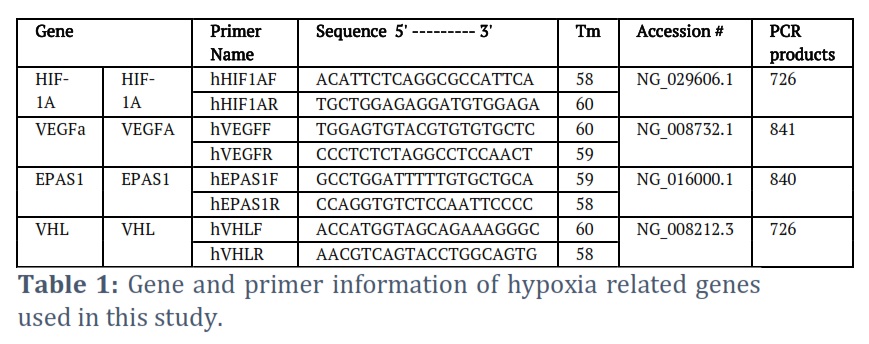
Primers
Genomic clones of human hypoxia-related genes were selected from the NCBI nucleotide database (https://www.ncbi.nlm.nih.gov/nucleotide). These clones contained the complete gene sequences including exons, intron, 5' flanking region, and 3' flanking region. Primers were designed based on the sequence of human genomic clones for every gene of high-altitude related genes (Table 1). Primers were designed to amplify noncoding regions because it has more SNPs frequency. Primers were synthesized at Macrogen company (https://www.macrogen.com/en/main). Table (1) summarizes gene and primer information used in this study. Specimens were procured from fresh EDTA-treated whole blood samples. The DNA extraction was meticulously executed utilizing the QIAamp Blood Mini Kit, in conjunction with the QIAcube Connect Analyzer. Delving into the extraction procedure, the process commences with the addition of 200 µl of the sample to 20 µl of Proteinase K solution. Subsequently, the lyse buffer is introduced to the aforementioned solution, which is then incubated in an orbital shaker at a temperature of 56 °C. This mixture is then channeled into a spin column situated within a rotor adapter, facilitating the binding of the nucleic acid to the silica membrane of the spin column. To purge contaminants, 500 µl of Washing Solution 1 is amalgamated with the lysate, followed by a centrifugation phase lasting 1 minute at 8000 rpm. The procedure is furthered by the addition of 500 µl of Washing Solution 2 to the filtrate obtained from the prior step, with centrifugation executed at 14000 rpm for a duration of 3 minutes. An additional centrifugation at maximum speed is subsequently performed. Culminating the process, the spin column is relocated to a 2 ml microcentrifuge tube, where 100 µl of elution buffer is introduced to the purified nucleic acid. The resultant nucleic acid product is preserved at a temperature of -20 °C, primed for subsequent testing.
Quantification and quantitation of DNA
Post-extraction, it is imperative to ascertain the quality and quantity of the DNA to ensure its suitability for subsequent applications. A widely adopted method for this assessment is spectrophotometry, specifically utilizing the "Nanodrop" instrument. The first metric of interest is DNA concentration, which serves as an indicator of the yield from the extraction process. For the DNA sample to be deemed adequate, its concentration should surpass 25 ng. Beyond mere concentration, the purity of the DNA is of paramount importance. This is gauged by measuring the optical density at 260 nm, a wavelength where nucleic acids absorb light most efficiently. However, the mere optical density at this wavelength doesn't provide a holistic view of the sample's purity. To further refine this assessment, the A260/A280 ratio is calculated. This ratio is instrumental in detecting the presence of potential contaminants, such as organic chemicals and proteins. An acceptable A260/A280 ratio, indicative of a relatively uncontaminated sample, typically ranges between 1.7 and 1.9. Any deviation from this range might suggest the presence of impurities, warranting further purification or investigation.
PCR amplification
PCR amplification was performed on DNA samples to target the four genes: Hif-1α, Vegfa, Epas, and Vhl, using specific primers listed in Table 2. The reactions utilized a simplified protocol with a focus on the annealing temperature specific to each gene. For each, the annealing was conducted at 58⁰C for 30 seconds as part of a broader thermal cycle. This approach builds upon established methods, adapting where necessary to accommodate gene-specific requirements. Further details are streamlined in favor of referencing pertinent studies that outline similar PCR conditions and methodologies.
Sequencing and sequence analysis
PCR products were sequenced at Macrogen company (https://www.macrogen.com/en/main) using the specific primers for each gene. The obtained sequences were trimmed and used for alignment to detect the single nucleotide polymorphism (SNPs). Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo) at the EMBL’s European Bioinformatics Institute (EMBL-EBI) (https://www.ebi.ac.uk/services) was used for multiple alignment of the obtained sequences for each gene. Alignment data was used to construct a phylogenetic tree among the aligned sequences.
Ethical considerations
Adhering to the highest standards of academic research, the study protocol was crafted with a keen emphasis on ethical considerations. It received the esteemed approval from the ethical and research committee of King Fahad Armed Forces Hospital, as evidenced by Reference Ethical Number: REC 504. Every step, from data collection to analysis, was conducted with an unwavering commitment to upholding the principles of ethical research.
Inclusion criteria
The study was discerning in its volunteer selection, focusing on individuals living in high altitude or sea level for at least 20 years. Their health and medical status were rigorously verified through an exhaustive review of medical files, detailed clinical histories, and other pertinent medical documentation. For thorough and reliable research results, race was taken into consideration when the study participants were selected, as it has been linked to a number of factors, including health and genetics, which are shared by the groups being studied. More than one technologist was involved in performing the experiment; however, the biases were minimized by having standardized specimen collection protocols and analyzing specimens on an automated analyzer as a part of a total lab automation unit without human handling in the pre-analytical, analytical, and post-analytical phases of testing.
Exclusion criteria
To maintain the integrity and purity of the data, volunteers who had undergone blood transfusions recently, individuals with known chronic illnesses, those on medications that could influence genetic parameters, and anyone with a family history of genetic disorders were systematically excluded. Furthermore, any participant deemed unhealthy based on their medical records or those with conditions that could potentially skew the results were also excluded from the study. More than 100 volunteers for data selection were checked, and only 22 were matched with our inclusion and exclusion criteria.
Results![]()
Hypoxia-Inducible Factor (HIF-1A)
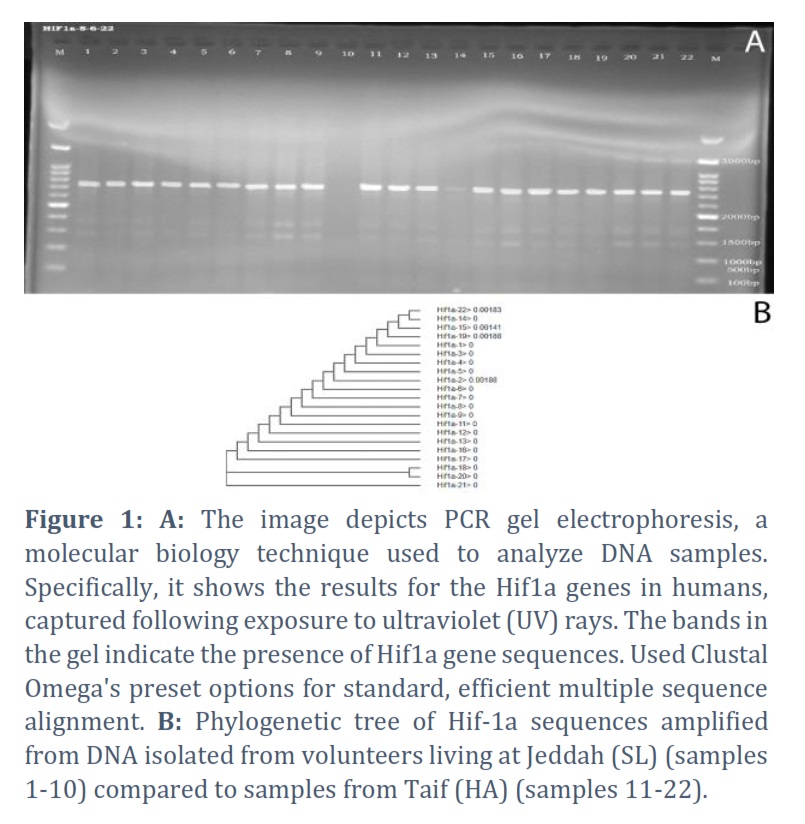
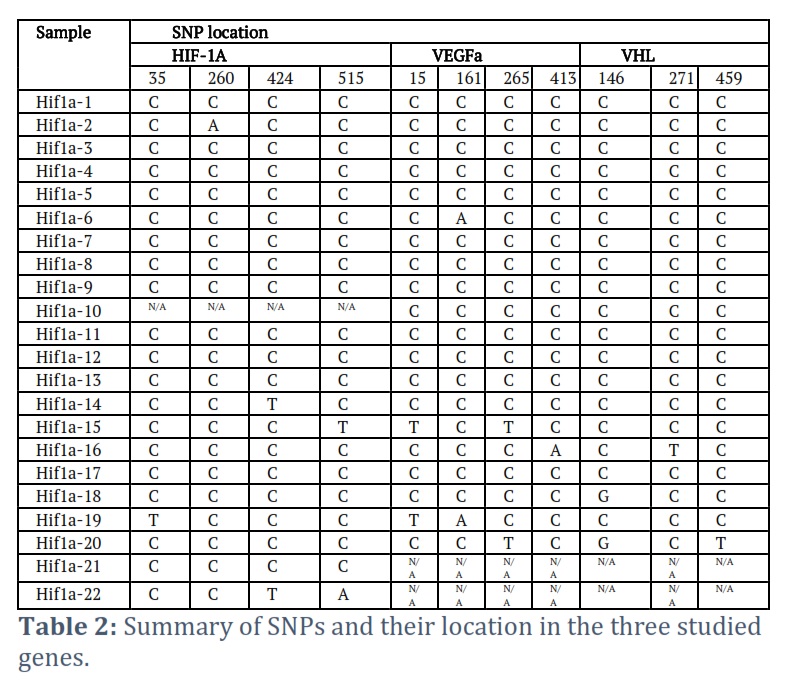
The obtained sequences were cleaned and trimmed. For the Hif1a gene, 9 final sequences for the sea level (SL) group and 12 final sequences for the high-altitude group (HA) were used for multiple alignment. Alignment of the 21 sequences resulted in the detection of 4 SNPs at position 35, 260, 424, and 515 of the clean sequence (Fig. 3). The 4 SNPs were detected in sample number 2 of the SL group and sample number 14, 15, 19, and 22 of the HA group (Fig. 2, Table 1). Sample #2 had one C>A SNP at 226. This sample is for a 50-year-old male. Sample # 14 (65 years old female) had C>T SNP at 424, sample #15 (74 years old female) had a C>T SNP at 515, sample 19 (79 years old male) had a C>T SNP at 35, and sample 22 (81 years old male) had a C>T SNP at 424 and a C>A SNP at 515 (Table 2). The reference alignment that showcases the SNPs in each gene is in the appendix referenced as the first.
The aligned 20 Hif1a sequences were used to construct the phylogenetic tree among sequences. The detected SNPs in samples 2 (SL), 14, 15, 19, and 22 (HA) separated them from other samples with different distances shown on the right of each sample (Fig. 2). Samples 18 and 20 were separated on one clade, samples 14 and 22 were also separated on one clade, and all other samples were separated on independent branches.
Vascular endothelial growth factor A (VEGFa)
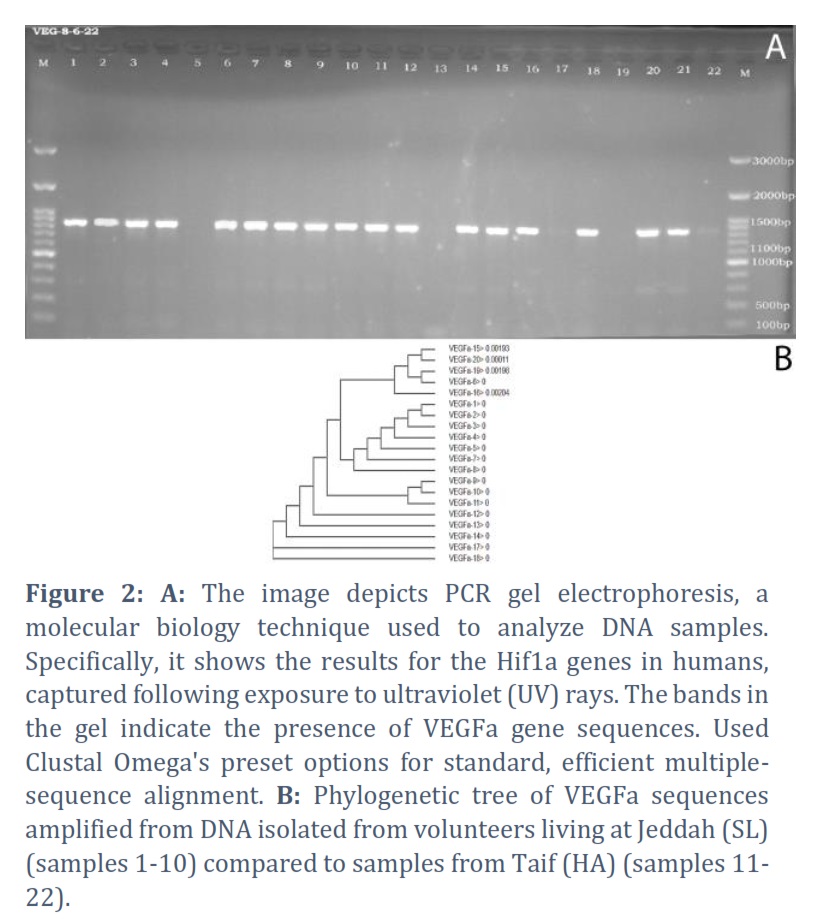
For the VEGFa gene, a clean sequence of 490 bp was obtained for multiple alignment of 20 DNA sequences, 10 sequences for SL and 10 for HA, resulting in the detection of 4 SNPs in individuals 6, 15, 16, 19, and 20. Individual 6 from the SL group had one C>T SNP at position 15. This is a 70-year-old female. Individual 15 is a 74-year-old female which had two C>T SNP at 15 and at 265 positions from the start of the aligned sequence. Individual 16 is 65 years old female who showed one C>A SNP 413 position. Individual 19 is 79 years old male. It had one C>T SNP at position 15 and G>A SNP at position 161. Individual 20 is 67 years old male who showed one C>T SNP at position 265. The reference alignment that showcases the SNPs in each gene is in the appendix referenced as the second.
The phylogenetic tree was constructed using the aligned 20 sequences of VEGFa. The main feature of the phylogenetic tree is the separation of samples with SNPs including sample 6 (SL) and samples 15, 16, 19, 20 (HA) on one sub-clade of the tree. One other sub-clade included samples 1, 2, 3, 4, 5, 7, and 8 (SL). The remaining samples were distributed on 8 sub-clades 5 of them with only one sample each.
The phylogenetic analysis reveals relationships between individuals who exhibited genetic changes, specifically those from the high-altitude regions. Notably, the slight numerical variations in the identified Single Nucleotide Polymorphisms (SNPs) suggest that the genetic differences among these high-altitude residents are minimal.
von-Hippel-Lindau (VHL)-ubiquitin ligase (VHL)
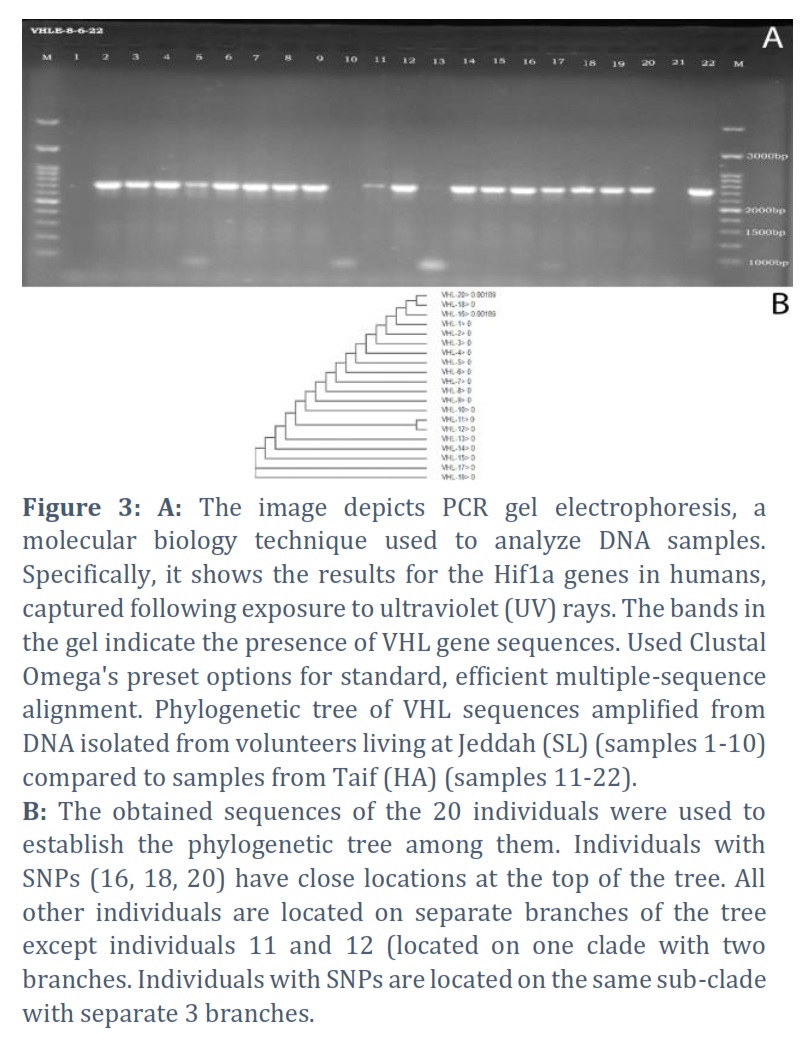
Multiple alignment of 20 DNA sequences (10 SL, 10 HA) resulted in the detection of 3 SNPs in individuals 16, 18, and 20. Individual 16 has one C>T SNP at 271. This is a 65-year-old female. Individual 18 is a 63-year-old male which had one A>G SNP at 146 from the start of the aligned sequence. Individual 20 is a 67-year-old male who showed two SNPs; A>G at 146 and C>T at 459. The reference alignment that showcases the SNPs in each gene is in the appendix referenced as the third.
Amplified from DNA isolated from volunteers living at Jeddah (Sea Level) (samples 1-10) compared to samples from Taif (High Altitude) (samples 11-22).
1. Sample 2 (Sea Level): SNP1-35 shows a C to A change.
2. Sample 14 (High Altitude): SNP3-424 shows a C to T change.
3. Sample 15 (High Altitude): SNP3-424 shows a C to T change.
4. Sample 19 (High Altitude): SNP1-35 shows a C to T change.
5. Sample 22 (High Altitude): SNP3-424 shows a C to T change and SNP4-515 shows a C to A change.
The information above indicates specific SNP changes at different locations within the Hif-1a gene, comparing samples from sea level (Jeddah) to those from a high altitude (Taif).
Amplified from DNA isolated from volunteers living at Jeddah (Sea Level) (samples 1-10) compared to samples from Taif (High Altitude) (samples 11-22).
1. Sample 6 (Sea Level): SNP1-15 shows a G to A change.
2. Sample 15 (High Altitude): SNP1-15 shows a C to T change, and SNP3-265 also shows a C to T change.
3. Sample 16 (High Altitude): SNP3-265 shows a C to A change.
4. Sample 19 (High Altitude): SNP1-15 shows a C to T change, and SNP2-161 shows a G to A change.
5. Sample 20 (High Altitude): SNP3-265 shows a C to T change.
The results indicate specific SNP changes at different locations within the VEGFa gene, comparing samples from sea level (Jeddah) to those from a high altitude (Taif).
The information presented (in the tables) specific SNP changes in the VHL gene among individuals from different altitudes, highlighting genetic variations between populations at sea level and high altitude.
1. Sample 16 (High Altitude): SNP2-271 shows a C to T change.
2. Sample 18 (High Altitude): SNP1-146 shows an A to G change.
3. Sample 20 (High Altitude): SNP1-146 shows an A to G change, and SNP3-459 shows a C to T change.
Occurrence of SNPs in Three Different Genes at Sea Level (SL) and High Altitude (HA):
1. Hif-1a Gene:
– SL: 1 occurrence out of 9 (11.11%)
– HA: 3 occurrences out of 12 (25%)
2. VEGFa Gene:
– SL: 1 occurrence out of 10 (10%)
– HA: 4 occurrences out of 10 (40%)
3. VHL Gene:
– SL: 0 occurrences out of 10 (0%)
– HA: 3 occurrences out of 10 (30%)
The information indicates the frequency of SNP occurrences in the Hif-1a, VEGFa, and VHL genes among individuals at sea level and high altitude.
The phylogenetic analysis reveals relationships between individuals who exhibited genetic changes, specifically those from the high-altitude regions. Notably, the slight numerical variations in the identified Single Nucleotide Polymorphisms (SNPs) suggest that the genetic differences among these high-altitude residents are minimal.
Summary of SNPs in Hif-1a Gene from Sea Level and High-Altitude Samples:
1. Sample 2 (Sea Level): SNP1-35, C>A change.
2. Sample 14 (High Altitude): SNP3-424, C>T change.
3. Sample 15 (High Altitude): SNP3-424, C>T change.
4. Sample 19 (High Altitude): SNP1-35, C>T change.
5. Sample 22 (High Altitude): SNP3-424, C>T change; SNP4-515, C>A change.
The information highlights specific changes in the Hif-1a gene among samples from different altitudes.
The information indicates specific SNP changes in the VEGFa gene among samples from different altitudes:
1. Sample 6 (Sea Level): SNP1-15, G>A change.
2. Sample 15 (High Altitude): SNP1-15, C>T change; SNP3-265, C>T change.
3. Sample 16 (High Altitude): SNP3-265, C>A change.
4. Sample 19 (High Altitude): SNP1-15, C>T change; SNP2-161, G>A change.
5. Sample 20 (High Altitude): SNP3-265, C>T change.
Summary of SNPs in VHL Gene from Sea Level and High-Altitude Samples
1. Sample 16 (High Altitude): SNP2-271, C>T change.
2. Sample 18 (High Altitude): SNP1-146, A>G change.
3. Sample 20 (High Altitude): SNP1-146, A>G change; SNP3-459, C>T change.
This information reflects specific SNP changes in the VHL gene among high-altitude samples.
Our results also indicate the frequency of SNP occurrences in the Hif-1a, VEGFa, and VHL genes among individuals at sea level and high altitude:
1. Hif-1a Gene:
– SL: 1 occurrence out of 9 (11.11%)
– HA: 3 occurrences out of 12 (25%)
2. VEGFa Gene:
– SL: 1 occurrence out of 10 (10%)
– HA: 4 occurrences out of 10 (40%)
3. VHL Gene:
– SL: 0 occurrences out of 10 (0%)
– HA: 3 occurrences out of 10 (30%)
Figures & Tables
The adaptation of human populations to varied environmental conditions, particularly altitude, has been a focal point of genetic and physiological research [2]. This study, conducted in the distinct altitudinal regions of West Saudia Arabia, provides a comprehensive analysis of the genetic variations associated with high-altitude adaptation.
Central to our investigation were the genes Hif-1a, VEGFa, and VHL. The Hif-1a gene, known for its regulatory role in oxygen homeostasis, exhibited specific variations that suggest a modulated cellular response to hypoxia as seen in the figures and tables that were provided above. This aligns with previous studies that have highlighted the significance of Hif-1a in cellular adaptations to hypoxic conditions [1]. The VEGFa gene, integral to angiogenesis and vascular permeability, presents variations that may indicate differential vascular responses in the high-altitude (HA) group. Given the established role of VEGFa in vascular adaptations to hypoxic environments, these variations could be indicative of a refined vascular adaptation mechanism in populations residing at higher altitudes.
The VHL gene, however, presented the most intriguing findings. With single nucleotide polymorphisms (SNPs) observed predominantly in the HA group, the VHL gene, a key component of the oxygen-sensing pathway, emerges as a potential marker for high-altitude adaptation. Its regulatory role in the degradation of HIFs suggests that variations in this gene might offer a selective advantage to individuals residing at higher altitudes. This observation is consistent with recent studies that have identified the role of VHL in high-altitude adaptations in various global populations [15]
The subsequent phylogenetic tree analysis further substantiated these genetic findings. Mostly, we have found results in individuals from the HA group bearing SNPs suggests a shared evolutionary trajectory, potentially indicative of the selective pressures of high-altitude environments. Such genetic clustering, previously observed in high-altitude populations such as Tibetans and Andeans, supports the hypothesis of convergent evolutionary strategies across different high-altitude populations [17].
Transcriptome analysis has been employed in numerous studies to disclose the underlying changes in gene expression as differentially expressed genes (DEGs) in response to biotic and abiotic changes including HA. The molecular adaptations to living at HA and its associated disorders including hypoxia are controlled by changes in gene expression of so many genes (transcriptome) [9, 20, 22]. For example, transcriptome analysis of 6 tissues of 5 Tibetan high-altitude vertebrates showed transcriptomic variations among species as they adapt to living at HA [22]. Also, transcriptome and proteome analysis of human platelets under normoxia compared to hypobaric conditions revealed changes in gene expression including membrane glycoproteins, integrin subunits, and alpha-granule chemokines. The results of this study concluded that living at HA triggered higher response to invoke thrombosis [20]. In addition, transcriptome analysis of lung and gluteus tissues of two species of cattle (living at sea level), Tibetan cattle (living at a moderate altitude), and yak (living at a high altitude) resulted in transcriptomic differences. The recorded DEGs included CD36 antigen (up-regulated) and CD59 antigen (down-regulated) in yak in comparison to the other animals. These and other gene expression profiles suggested that these differences could inhibit blood coagulation and reduce the risk of pulmonary edema.
Proteome is the functional interpretation of a transcriptome, therefore changes in transcriptome are reflected in the proteome profile. Proteome analysis of Yak during adaptation to high altitude showed upregulation of proteins and downregulation of proteins compared to Tibetan, Holstein, and Sanjiang cattle consecutively. The differentially expressed proteins included oxidative phosphorylation and electron transport chain proteins. Most of these proteins were increased in yak indicating that yak might enhance mitochondrial activity to maintain metabolic rates at high altitude. This could be one adaptive mechanism to high altitude. Also, yak showed high level of thioredoxin reductase 2 for protection from oxidative damages caused by living at HA.
In this study, we employed gel electrophoresis as a robust analytical technique to separate and visualize DNA fragments, thereby facilitating the identification of Single Nucleotide Polymorphisms (SNPs). Gel electrophoresis enabled us to achieve high-resolution separation of DNA fragments, thereby providing a clear snapshot of the genetic variations present in the samples.
Interestingly, our findings revealed a significant geographical disparity in the distribution of SNPs. A majority of the SNPs were predominantly found in the Taif region, characterized by its high-altitude (HA) environment, as opposed to the Jeddah region, which is at sea level (SL). This observation could be attributed to the unique environmental pressures exerted by high-altitude conditions, potentially driving genetic adaptations in the population residing in the Taif region.
When compared with existing literature, the genetic adaptations observed in West Saudi Arabia align with broader patterns of high-altitude adaptability. However, the unique genetic markers identified in this study, especially concerning the VHL gene, emphasize the region-specific nature of these adaptations. While overarching patterns of adaptability may be consistent across populations, regional environmental and evolutionary pressures contribute to the nuanced genetic variations observed.
In summation, this study offers a detailed overview of the genetic underpinnings of high-altitude adaptation in West Saudi Arabia. The identified genetic variations, while consistent with broader patterns of high-altitude adaptability, present unique regional signatures and confirmed SNPs changing with those individuals who are living at high altitude. Further research is warranted to elucidate the physiological implications of these genetic variations and to identify additional adaptive genes that may play a role in high-altitude adaptation.
Our in-depth analysis has unveiled the nuanced genetic adaptations that high-altitude residents in Taif have undergone in comparison to their sea-level counterparts in Jeddah. The presence of specific SNPs in the Hif-1a, VEGFa, and VHL genes among the HA group underscores the genetic predispositions and evolutionary adaptations to cope with hypoxic conditions. The phylogenetic trees further corroborate these findings, with SNP-bearing individuals from the HA group clustering distinctly, suggesting a shared evolutionary trajectory. Notably, the VHL gene exhibited SNPs exclusively in the HA group, emphasizing its potential role in high-altitude adaptation. This research not only enhances our understanding of human adaptability to altitude-induced hypoxia but also sets the stage for future studies exploring therapeutic avenues for altitude-related ailments. The intricate correlation between genetics and environment, as evidenced by our findings, expands on the knowledge of resilience and adaptability of human genetics.
Conflict of Interest
The authors declare that there is no conflict of interest.
All authors contributed extensively to the work presented in this paper. Siraj B. Alharthi, Ijtihed Kilani, Abdullah Alsoliman, Romar P. Pascual, Mohamed H. Zainy, Ahmed S Albariqi ,Yahya M. Alzahrani, Ahmed Alshehri, Ahmed M. El-Shehawi, Hashim M. Borayk, and Mohamed Morsi M. Ahmed were actively involved in every stage of the research process, including conceptualization, methodology development, fieldwork, sample collection, laboratory analysis, data analysis, and interpretation of the results. Their efforts encompassed DNA extraction, sequencing, PCR amplifications, biochemical assays, statistical and bioinformatics analysis, phylogenetic analysis, and preparation and revision of the manuscript. Each author played a critical role in ensuring the accuracy, reliability, and scientific integrity of the study, demonstrating a collaborative approach that was fundamental to the success of the research.
![]() References
References
- Khurana P, Gupta A, Sugadev R, Sharma YK, Kumar BH. HAHmiR.DB: a server platform for high-altitude human miRNA-gene regulatory networks and associated regulatory circuits. Europe PMC (2021); 10(10), 1000406.
- Alharthi S, Alsulaiman A, Mutawakil M, Morsi M, Ahmed MS. Genetic & Immunological Effects of High Altitude Illnesses on Human Health: Review. Journal of High Altitude Medicine (2023); 12: 156-168.
- Irarrázaval S, Allard C, Campodónico J, Pérez D, Strobel P, Vásquez L, Urquiaga I, Echeverría G, Leighton F. Oxidative stress in acute hypobaric hypoxia. High Altitude Medical Biology (2017); 18(2): 128-134.
- Liu Z, Chen H, Xu T, Wang X, Yao C. HSPA1A gene polymorphism rs1008438 is associated with susceptibility to acute mountain sickness in Han Chinese individuals. Molecular Genetics & Genomic Medicine (2020); 8(8): e1322.
- Giaccia AJ, Simon MC, Johnson R. The biology of hypoxia: the role of oxygen sensing in development, normal function, and disease. Genes Development (2004); 18(18): 2183-2194.
- Mehta SR, Chawla A, Kashyap AS. Acute Mountain Sickness, High Altitude Cerebral Oedema, High Altitude Pulmonary Oedema: The Current Concepts. Medical Journal of Armed Forces India (2008); 64(2): 149-153.
- Bishop T, Ratcliffe PJ. Signaling hypoxia by hypoxia-inducible factor protein hydroxylases: a historical overview and future perspectives. Hypoxia (Auckland, N.Z.) (2014); 5(2): 197-213.
- Hopfl G, Ogunshola O, Gassmann M. Hypoxia and high altitude. The molecular response. Advanced Exposure Medical Biology (2003); 543: 89-115.
- Gurung PD, Upadhyay AK, Bhardwaj PK, Sowdhamini R, Ramakrishnan U. Transcriptome analysis reveals plasticity in gene regulation due to environmental cues in Primula sikkimensis, a high altitude plant species. BMC Genomics (2019); 20(1): 989.
- Ananthakrishnan. Environmental triggers in IBD: a review of progress and evidence. Nature Reviews Gastroenterology & Hepatology (2018); 7: 345-357.
- Das B, Ghosh TS, Kedia S, Rampal R, Saxena S, Bag S, Mitra R, Dayal M, Mehta O, Surendranath A, Travis SPL, Tripathi P, Nair GB, Ahuja V. Analysis of the gut microbiome of rural and urban healthy Indians living in sea level and high altitude areas. Scientific Reports (2018); 8(1): 10104.
- Eissler Y, Galvez MJ, Dorador C, Hengst M, Molina V. Active microbiome structure and its association with environmental factors and viruses at different aquatic sites of a high-altitude wetland. Microbiology Open (2019); 8(3): e00667.
- Fan Q, Wanapat M, Yan T, Hou F. Altitude influences microbial diversity and herbage fermentation in the rumen of yaks. BMC Microbiology (2020); 20(1): 370.
- Hackett PH, Roach RC. High-altitude illness. New England Journal of Medicine (2001); 345(2): 107-114.
- Guo L, Zhang J, Jin J, Gao X, Yu J, Geng Q, Li H, Huang L. Genetic variants of endothelial PAS domain protein 1 are associated with susceptibility to acute mountain sickness in individuals unaccustomed to high altitude: A nested case‑control study. Spandidos Publications (2015); 10(3): 907-914.
- Jablonski NG. Genes for the high life: New genetic variants point to positive selection for high altitude hypoxia in Tibetans. Zoological Research (2017); 38(3): 117.
- Simonson TS, Yang Y, Huff CD, Yun H, Qin G, Witherspoon DJ, Bai Z, Lorenzo FR, Xing J, Jorde LB, Prchal JT, Ge R. Genetic evidence for high-altitude adaptation in Tibet. Europe PMC (2010); 329(5987): 72-75.
- Hanaoka M, Droma Y, Basnyat B, Ito M, Kobayashi N, Katsuyama Y, Kubo K, Ota M. Genetic variants in EPAS1 contribute to adaptation to high-altitude hypoxia in Sherpas. PLoS One (2012); 7(12): e50566.
- Heinrich EC, Wu L, Lawrence ES, Cole AM, Anza-Ramirez C, Villafuerte FC, Simonson TS. Genetic variants at the EGLN1 locus associated with high-altitude adaptation in Tibetans are absent or found at low frequency in highland Andeans. Annals of Human Genetics (2019); 83(3): 171-176.
- Shang C, Wuren T, Ga Q, Bai Z, Guo L, Eustes AS, McComas KN, Rondina MT, Ge R. The human platelet transcriptome and proteome is altered and pro-thrombotic functional responses are increased during prolonged hypoxia exposure at high altitude. Platelets (2020); 31(1): 33-42.
- Djarova T, Bardarev D, Boyanov D, Kaneva R, Atanasov P. Performance enhancing genetic variants, oxygen uptake, heart rate, blood pressure and body mass index of elite high altitude mountaineers. Acta Physiologica Hungarica (2013); 100(3): 289-301.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0