

Full Length Research Article
Screening and Optimization of Dextransucrase Production from Lactobacillus acidophilus
Yasameen Hasan Ali Al-anbakey*, Sahira Nsayef Muslim, Israa Hussein Hamzah
Adv. life sci., vol. 11, no. 1, pp. 130-135, February 2024
*- Corresponding Author: Yasameen Hasan Ali Al-anbakey (yasameenhasan75@gmail.com)
Authors' Affiliations
[Date Received: 16/10/2022; Date Revised: 27/12/2023; Date Published: 25/02/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Dextransucrase is an enzyme synthesized by many bacterial genera, which catalyzes the hydrolysis of sucrose to produce dextran, a polysaccharide with a wide range of medical, pharmaceutical, and industrial applications. The study aimed at isolating Lactobacillus species from the human vagina, assessing the production of dextransucrase, and determining the optimal growth conditions.
Methods: A total of 53 bacterial samples were isolated from the reproductive tract of healthy women. Out of these 53 samples, 21 isolates of Lactobacillus spp. were obtained. Dextransucrase production was assessed using both the mucoid and spectrophotometric techniques. The effects of natural carbon, nitrogen sources, concentrations of nitrogen, protein, and sucrose, inoculum size, pH, temperature, and incubation time were evaluated to determine the optimal conditions.
Results: The findings showing that Lactobacillus acidophilus isolates produced more dextransucrase than isolates of Lactobacillus plantarum, with Lactobacillus acidophilus V19 being the strongest producer isolate due to its highest specific activity. The best conditions for producing dextransucrase were to use a dextransucrase production medium that underwent incubation at 37°C for 48 hours at pH 5.5, with a concentration of 6% sucrose, and 4% inoculum size. The most efficient source of nitrogen was 6% beef extract, and the best source of carbon was dates. At this point, the dextransucrase activity was at its highest.
Conclusion: The findings of the study revealed that the optimal conditions for dextransucrase production are a pH of 5.5, 4% inoculum size, 6% dates extract as a carbon source, 6% beef extract as a nitrogen source, and 48 hours of incubation at 37°C.
Keywords: Ecological; Optimization; Dextransucrase; Lactobacillus acidophilus
Introduction![]()
Lactobacillus species were identified as the Gram-positive rod-shaped bacteria that make up the vaginal microbiome. The predominant bacteria in the vaginal microbiota are called lactobacilli, and they are rod-shaped, facultative, anaerobic lactic acid bacteria that are catalase-negative and do not generate spores. As probiotics, several Lactobacillus strains are employed. Nonetheless, by stopping the colonization of dangerous microbes, Lactobacillus plays a vital role in preserving the health of the female reproductive system. When Lactobacillus species are present in high fluid levels in healthy women, they organize the vaginal microbiota, which significantly affects the microflora of the ecosystem [1]. Lactobacilli have vital functions in nutrition, metabolism, immunology, and pathogen defence and are frequently associated with the intestinal microbiota of newborns [2]. Extracellular enzyme called bacterial dextransucrase hydrolyzes sucrose to fructose and glucose monomers, producing an intermediate called glycosyl-enzyme containing glucose that then polymerizes to produce dextran [3].
Dextrans are made by transglucosylases called dextransucrases, and their main chain contains more than 50% -(16) links. The branching ratios of dextran molecules might vary depending on the producing enzyme -(12) or – (13). It has been suggested recently that the production of this first dextran occurs through the combined activity of two enzymes, namely dextransucrase and branching-sucrase [4, 5]. Extracellular glucosyltransferase (GTF), or dextransucrase facilitates the transfer of D-glucopyranosyl residues from sucrose to dextran while releasing fructose [6]. The enzyme dextransucrase produces the polysaccharide dextran. Dextran is utilized as a prebiotic, an antithrombotic agent, a lubricant in some eye drops, and to solubilize other ingredients in some intravenous solutions, among many other significant uses in food, medicine, and pharmaceuticals. When administered intravenously, dextran provides an osmotically neutral fluid that, once within the body, is broken down by cells into glucose and free water, prodrug, and conjugated with additional medications that act as a delivery system [7].
The present study was conducted to isolate Lactobacillus spp. from the human vagina, screen for the formation of dextransucrase, and optimize growth conditions to determine the optimal conditions for dextransucrase production.
Methods![]()
Collection of bacterial isolates
A total of 53 clinical samples were collected from various medical facilities in the city of Baghdad, Iraq for isolation of Lactobacillus spp. The samples were collected from healthy pre-menopause women's vagina. All Lactobacillus spp. isolates were identified utilizing the VITEK 2 compact system, microscopy, culture characteristics, and biochemical tests after collection.
Screening dextransucrase synthesis using the mucoid method
The dextransucrase production medium was prepared by adding sucrose (60g/l),yeast extract (10g/l), CaCl2 (0.14g/l), MgSO4(0.04g/l), FeSO4(0.04g/l), MnSO4 (0.02g/l), NaCl (0.01g/l), H3PO4 (5.7g/l), and 15g agar powder to 100ml distilled water. The pH was adjusted to 5.0. The medium was then autoclaved to achieve sterilization. Finally, 24-hour-old culture of Lactobacillus spp. isolates were introduced into the media. At 37o C, the culture was incubated for 24 hours, and the isolates' ropy mucoid morphology was classified as a positive dextran producer [7].
Screening dextransucrase synthesis using the spectrophotometric technique
The spectrophotometric approach was used to assess the selected Lactobacillus spp. isolates that were observed as dextransucrase-generating isolates. After inoculating 2% of Lactobacillus spp. suspension with 9×108cfu/ml (equivalent to 0.5 McFarland standard or absorbance of 0.134 at 600 nm), the dextransucrase production medium without agar powder was incubated for 24 hours at 37 °C. Following incubation, the culture was centrifuged for 10 minutes at 10,000 rpm to remove the biomass. The supernatant was then used to measure the concentrations of dextran, protein, and dextransucrase activity. Dextransucrase activity was determined using a spectrophotometer equipped with an adapted version of Miller's (1959) method, 50mL of pH 5.0, 0.1M phosphate buffer and 0.5 millilitres of crude enzyme extract and 0.5 ml of reaction using sucrose as substrate. After the reaction mixture was incubated for 30 minutes at 30°C, the enzymatic process was terminated by submerging it in a water bath for five minutes. The mixture's final volume was 2 ml after the addition of 3,5dinitrosalicylic acid reagent. To calculate the concentration of reducing sugars, the spectrophotometer was used to determine the absorbance at 540nm.A 1 g of sucrose solution dissolved in 50mL of 0.1M phosphate buffer at pH 5.0 was combined with 0.5mL of distilled water (D.W.) to prepare the control. The dextransucrase activity was determined using the formula below:
![]()
Where the glucose released (μg) could be estimated from the standard curve, V is the volume of the enzyme sample, and T is hydrolysis time and equals 30 min. One unit of dextransucrase activity is defined as the amount of enzyme releasing 1 μmol of glucose per minute under specific conditions [8].
Determination of protein concentration
The Bradford (1976) method was used to assess the protein concentration using bovine serum albumin (BSA) [9].
Determination of the optimal conditions for dextransucrase production
Natural carbon sources
The selected bacterial isolate was subjected to a dextransucrase production medium containing different plant extracts including date, beetroot, peach, pear, tangerine, apple, honeydew, watermelon, sweet potato, pineapple, and orange at a concentration of 6% each, as carbon sources for the production of dextransucrase. Following the incubation period (48 hours at 37°C), the dextransucrase activity and protein content were determined for each carbon source.
Sucrose concentrations
The selected bacterial isolate was added to a dextransucrase production medium containing varying amounts of sucrose, namely 3, 4, 5, 6, 7, 8, and 9mg/100 mL. Following a 48-hour incubation period at 37°C, the dextransucrase activity and protein content were assessed for each carbon source concentration.
Nitrogen sources
The dextransucrase production medium was supplemented with a variety of inorganic and organic nitrogen sources, including peptone, tryptone, beef extract, yeast extract, ammonium chloride, ammonium sulphate, and ammonium persulfate, separately. After being inoculated with the selected bacterial isolate, the medium was then incubated at 37°C for 48 hours to determine the optimal nitrogen source that induces the best dextransucrase production. Dextransucrase activity and protein content were measured for each nitrogen supply concentration following incubation.
Nitrogen source concentrations
The selected bacterial isolate was added to a dextransucrase production medium containing different concentrations of the optimal nitrogen source, namely 1, 2, 3, 4, 5, 6, 7, 8, 9, and 10g/100mL. The dextransucrase activity and protein concentration were measured for each nitrogen source concentration after 48 hours of incubation at 37°C.
Inoculum size
The selected isolates were cultured at different inoculum sizes ranging from 1, 2, 3, 4, 5, 6, 7, 8, 9, and 10% after being inoculated into a dextransucrase production medium. After the time of incubation, the dextransucrase activity, as well as protein concentration was calculated for each inoculum size.
pH
After being infused into the dextransucrase production medium, the selected bacterial isolate was incubated at a variety of pH values (5, 5.5, 6, 6.5, 7, 7.5, 8, 8.5, 9, and 9.5). Following the incubation period, the activity and protein content were calculated for each of the test pH values.
Temperature
The selected strain was incubated at different temperatures (4, 15, 25, 30, 37, and 40°C) after being inoculated into the dextransucrase production medium. The levels of protein concentration and dextransucrase activity were assessed following each incubation time.
Incubation time
The chosen isolates underwent culturing for varied lengths of time after being inoculated into the dextransucrase production medium (24, 48, 72, and 96 hours). The levels of protein concentration and dextransucrase activity were assessed following each incubation time.
Results![]()
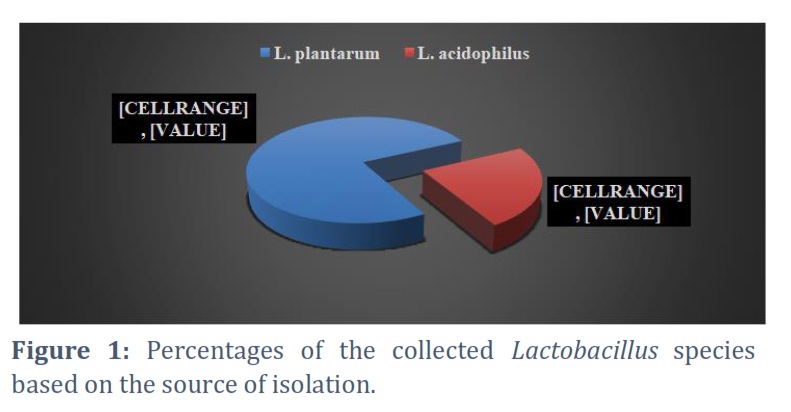
The results revealed that 21 (or 39%) of the 53 clinical samples that were collected were Lactobacillus spp. isolates that were recovered from vaginal swabs as depicted in Figure 1.
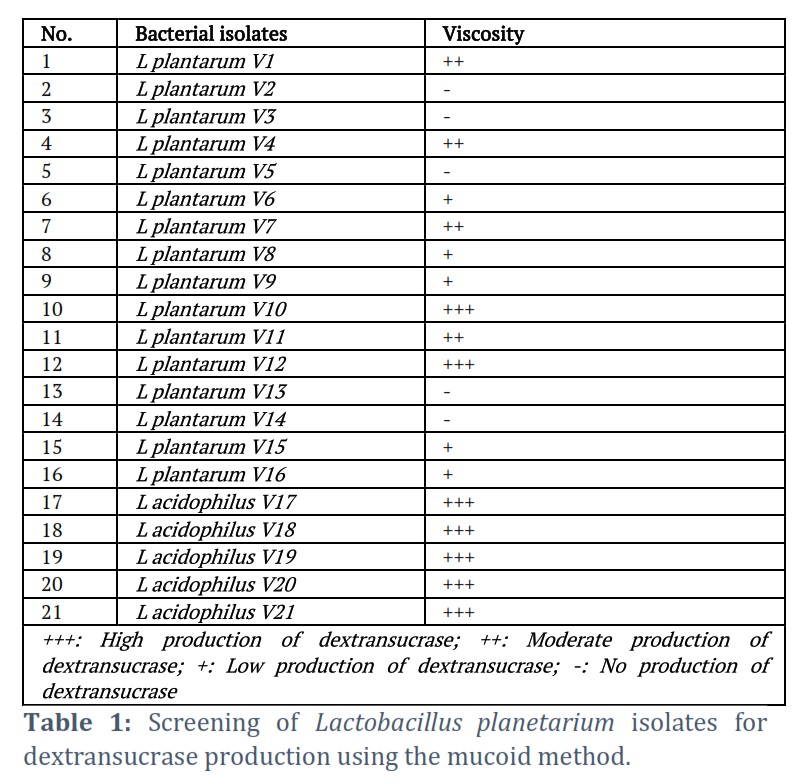
Of the 21 identified Lactobacillus spp., 16 (76.19%) were Lactobacillus plantarum isolates, and only 5 (23.8%) were Lactobacillus acidophilus isolates. The ability of all 21 isolates of Lactobacillus species to produce dextransucrase was examined. The detection and screening of dextransucrase production were recorded using slimy mucoid colonies on the surface of the dextransucrase screening medium. Only 16 isolates produced slimy mucoid colonies, according to the results displayed in Table 1.
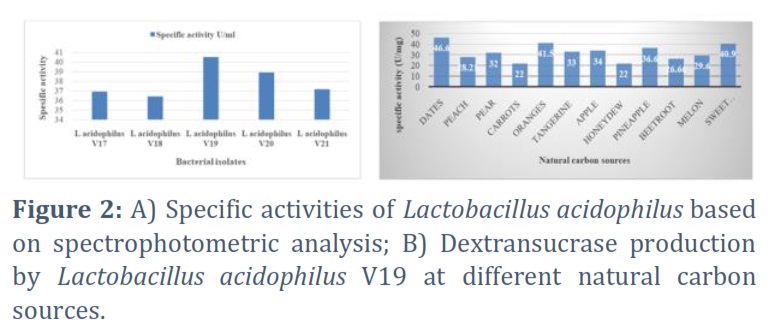
Four of the twenty-one isolates produced somewhat slimy mucoid colonies, five produced weak mucoid colonies, and seven of the isolates produced strong mucoid colonies. Furthermore, compared to Lactobacillus plantarum, Lactobacillus acidophilus had a higher dextransucrase. The results indicated that Lactobacillus acidophilus V19, with maximal dextransucrase activity equivalent to 7.29 and 40.4 U/mg of specific activity, was obtained by selecting the isolates of Lactobacillus acidophilus that produced high mucoid. The dextransucrase activity of these seven isolates varied, as illustrated in Figure 2A, with specific activities ranging from 34 to 40.5 U/mg. To determine the ideal carbon source for the synthesis of Lactobacillus acidophilus V19 dextransucrase, eleven distinct natural carbon sources were examined. The outcomes are shown in Figure 2B.
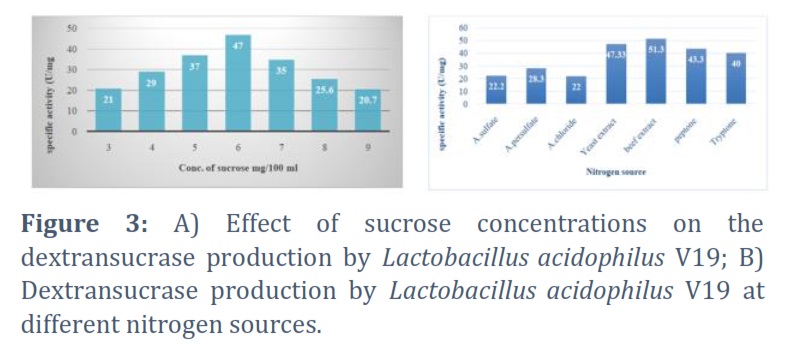
The ideal sucrose concentration for Lactobacillus acidophilus V19 to synthesize dextransucrase was found using several percentages of 1, 2, 3, 4, 5, 6, 7, 8, and 9%. The outcome demonstrated that the optimal sucrose concentration for the synthesis of dextransucrase was 6%, with a specific activity of 47 U/mg for dextransucrase. Conversely, 9% decreased specific activity to 20.7 U/mg, as illustrated in Figure 3A. Using a dextransucrase production medium, the effects of different nitrogen sources on dextransucrase synthesis were investigated. The outcomes seemed to be as illustrated in Figure 3B.
Figures & Tables
The simplest method for recognizing exopolysaccharied (EPS) formation is by looking at the phenotypic characteristics of the colonies, such as their ropy or slimy morphologies. Slimy colonies are characterized by mucilaginous colonies. On the other hand, when an inoculation loop is removed from the colony surface or cell pellet, ropy colonies produce long filaments. To evaluate the microbial capacity to create EPS, screening techniques based on cultivating the lab on a medium enriched with various sugars (lactose, lactose, galactose, or fructose) were employed [10]. The findings in the present study provide evidence that lab synthesizes dextran in a specific way when exposed to an environment that contains sucrose as a carbon source. Through the phosphotransferase system (PTS), sucrose can enter the cell directly and be metabolized by the body to produce either dextran or D-lactate [11].
Extracellular bacteria have enzymes called bacterial dextransucrases that hydrolyze sucrose in its fructose and polymerization to produce dextran. The fructose that results enters the bacterium through PTS to meet its metabolic requirement for glucose monomers, where it forms an intermediate with glucose [12].
The present study revealed that out of twelve different natural carbon sources, dates were the best, followed by sweet potatoes and oranges. The specific activity of dextransucrase was 46.6U/mg after incubation with dates. Conversely, the formation of dextransucrase was hindered by all other natural carbon sources, except carrots and honeydew, each having a specific activity of 22U/mg. Dates are considered the best natural carbon source since sucrose is a disaccharide, indicating that both glucose and fructose monomers are their building blocks. Dates are made of 70% sugars mainly glucose and fructose and small amounts of sucrose and maltose [11, 13].With a specific activity of 51.3U/mg, beef extract was found to be the most effective nitrogen source for the production of dextransucrase, followed by yeast extract, peptone, and tryptone.
On the other hand, the production of dextransucrase was significantly reduced by ammonium chloride, ammonium sulphate, and ammonium persulfate. These observations suggest that organic nitrogen sources are better dextransucrase inducers than inorganic sources, of which beef extract is the best. Sources of nitrogen are second in significance to carbon in terms of their role in the proliferation of microbial cells. In addition to being an essential component of proteins, nucleotides, and enzymes, nitrogen is also a cofactor that is essential to cellular metabolism [14].
However, dextran is an EPS that requires the enzyme dextransucrase for synthesis. Variable nitrogen sources can contain different amounts of amino acids, which could affect the synthesis of proteins (enzyme dextransucrase) and the production of dextran [14]. Meat extract dried into powder form is called beef extract powder. A variety of culture media are made from a nutrient-rich extract to aid in the growth of various microorganisms. It provides nitrogen, vitamins, amino acids, and carbon to the microbiological growth media [15].
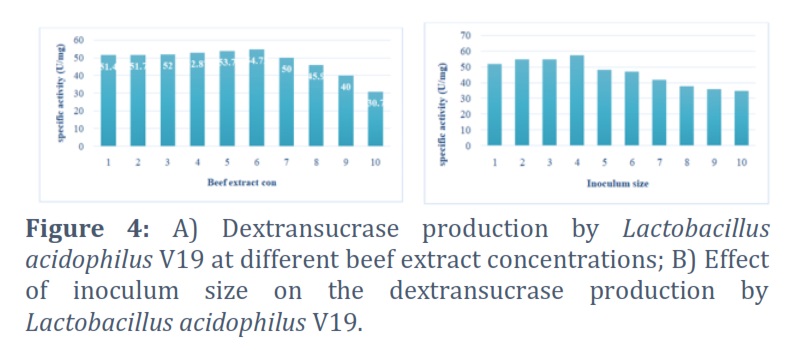
A range of concentrations of the beef extract, from 1 to 10%, was used to establish the ideal concentration of the selected nitrogen source for the synthesis of dextransucrase. The findings indicate that at a concentration of 6% beef extract, the productivity peaked, and the specific activity of dextransucrase was 54.72U/mg. Dextransucrase-specific activity and dextran concentration were both decreased by increasing the content of the beef extract. Figure 4 A and 4 B.
The impact of inoculum size on the generation of dextran was investigated. A range of inoculum sizes (1%–10%) was used to incubate Lactobacillus acidophilus V19. The findings showed that 4% was the ideal inoculum size for dextransucrase production, with 57.4 U/mg protein for dextransucrase-specific activity. However, dextransucrase-specific activity decreased to 34.8 U/ml as the inoculum size was raised. As inoculum size increases, the interaction between substrate, nutrients, and bacteria will intensify, enhancing bacterial metabolism, protein synthesis, and enzyme activity.
On the other hand, the high inoculum size results in competition for nutrients and substrates, as well as a decrease in enzyme activity. Consequently, early nutrient depletion from the medium and competition with high concentrations of elevated growing populations may be the cause of the activity drop with increasing inoculum size. Furthermore, the quick culture development was the cause of oxygen loss. The synthesis of dextransucrase varied at different pH values. It was found that to create dextransucrase, which has a specific activity of 60 U/mg, Lactobacillus acidophilus V19 required a pH of 5.5. The findings indicate that further pH readings showed a decrease in the activity of dextransucrase.
The enzyme's ability to catalyze a process can be affected by or entirely prevented by changes in the pH or acidity of the surrounding environment. When the pH reaches a threshold where the substrate molecule no longer fits, the chemical change will be impeded or not occurring at all because of the effects on the polar and non-polar intramolecular attraction and repulsive interactions, the enzyme configuration, and the active site [16, 17].
To find the ideal incubation temperature for Lactobacillus acidophilus V19 to produce dextransucrase, a range of temperatures including 4, 15, 25, 30, 37, and 40 ºC were investigated. At 37ºC, the maximum amount of dextransucrase was produced. At this temperature, the specific activity of dextransucrase was 60.54 U/mg proteins. However, the findings revealed that both higher and lower temperatures resulted in a decrease in dextransucrase activity. Also, after 48 hours of incubation, Lactobacillus acidophilus V19 produced dextransucrase-specific activity of 60.97 U/mg.
As the incubation period increased, the concentration dropped. After 96 hours, dextransucrase-specific activity reached 19.9U/mg. The bacterial population grew exponentially over the incubation period, depleting the surrounding nutrients. Wastes and harmful by products also began to build up, which inhibited the biological processes inside the cell. That could account for the enzyme's decline in enzymatic activity. It is also important to note that the built-up by products may cause disruptions between the enzyme and its substrate [18].
Compared to Lactobacillus plantarum isolates, Lactobacillus acidophilus isolates produced more of the enzyme dextransucrase. Moreover, the optimal conditions for producing dextransucrase were as follows: pH adjusted to 5.5 with 4% inoculum size, 6% dates extract as a carbon source, and 6% beef extract as a nitrogen source, incubated at 37°C for 48 hours.
Acknowledgment
The authors express their gratitude to Al-Mustansiriyah University (https://www.uomustansiriyah.edu.iq/) for providing the support and assistance needed to complete this research.
Conflict of Interest
The authors declare that there is no conflict of interest.
Yasameen Hasan Ali Al-Anbakey was responsible for collecting the data, performing the analysis, and writing the draft version of the manuscript while Sahira Nsayef Muslim and Israa Hussein Hamzah participated in the data analysis and produced the final version of the manuscript.
![]() References
References
- Mahmood NN, Hameed AA. Probiotic activity of Lactobacillus spp. from vaginal specimens against bacterial pathogens. Journal of University of Babylon for Pure and Applied Sciences, (2018); 26(5): 335-346.
- Ozen M, Piloquet H, Schaubeck M. Limosilactobacillusfermentum CECT5716: Clinical Potential of a Probiotic Strain Isolated from Human Milk. Nutrients, (2023): 15(9), 2207.
- Smith PJ, Ortiz-Soto ME, Roth C, Barnes WJ, Seibel J, Urbanowicz BR, Pfrengle F. ACS Sustainable ChemistryanD Engineering, (2020): 8(32): 11853–11871.
- Mu D, Zhou Y, Wu X, Montalban-Lopez M, Wang L, Li X, Zheng, Z. Secretion of Bacillus amyloliquefacienslevansucrase from Bacillus subtilis and its application in the enzymatic synthesis of l evan. ACS Food Science and Technology, (2021); 1(2): 249-259.
- Vuillemin M, Grimaud F, Claverie M, Rolland-Sabaté A, Garnier C, Lucas P, Monsan P, Dols-Lafargue M, Remaud-Siméon M, Moulis C.. A dextran with unique rheological properties produced by the dextransucrase from Oenococcuskitaharae DSM 17330. Carbohydrate Polymers, (2018); 179: 10-18.
- Besrour-Aouam N, Fhoula I, Hernández-Alcántara AM, Mohedano ML, Najjari A, Prieto A, Ruas-Madiedo P, López P, OuzariHI.. The role of dextran production in the metabolic context of Leuconostoc and Weissella Tunisian strains. Carbohydrate Polymers, (2021); 253: 117254.
- Guzman GYF, Hurtado GB, Ospina SA. New dextransucrase purification process of the enzyme produced by Leuconostocmesenteroides IBUN 91.2. 98 based on binding product and dextranase hydrolysis. Journal of biotechnology, (2018); 265: 8-14.
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry, (1976); 72(1-2): 248-254.
- Prechtl RM, Janßen D, Behr J, Ludwig C, Küster B, Vogel RF, Jakob F. Sucrose-induced proteomic response and carbohydrate utilization of Lactobacillus sakei TMW 1.411 during dextran formation. Frontiers in Microbiology, (2018): 9: 2796.
- Guérin M, Silva CRD, Garcia C, Remize F. Lactic acid bacterial production of exopolysaccharides from fruit and vegetables and associated benefits. Fermentation, (2020); 6(4): 115.
- Hosaka H, Kawamura M, Hirano T, Hakamata W, Nishio T. Utilization of sucrose and analog disaccharides by human intestinal bifid bacteria and lactobacilli: search of the bifid bacteria enzymes involved in the degradation of these disaccharides. Microbiological Research, (2020); 240: 126558.
- YasawyMI. The unexpected truth about dates and hypoglycemia. Journal of Family snd Community Medicine, (2016); 23(2): 115–118.
- Suribabu K, Hemalatha KPJ. Thermal Optimization of α-amylase Production in Brevibacillus sp. International Journal of Current Microbiology and Applied Sciences, (2016); 5(3): 909-916.
- Srinivas B, Padma PN. Screening of diverse organic, inorganic and natural nitrogen sources for dextran production by WeissellaSpp using Plackett-Burman design. International Journal of Scince and Technology Reseatch, (2014); 3(4): 319-328.
- Koskar J, Meremäe K, Püssa T, Anton D, Elias T, Rätsep R, Mäesaar M, Kapp K, Roasto M. Microbial Growth Dynamics in Minced Meat Enriched with Plant Powders. Applied Sciences, (2022); 12(21): 11292.
- Thi Nguyen HY, Tran GB. Optimization of Fermentation Conditions and Media for Production of Glucose Isomerase from Bacillus megaterium Using Response Surface Methodology. Scientifica, (2018); 2018: 6842843.
- Robinson PK. Enzymes: principles and biotechnological applications. Essays in Biochemistry, (2015);59:1-41.
- Krizsan SJ, Huhtanen P. Effect of diet composition and incubation time on feed indigestible neutral detergent fiber concentration in dairy cows. Journal of Dairy Science, (2013); 96(3): 1715-1726.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


