

Full Length Research Article
Molecular identification of Epstein-Barr virus in human placental tissue
Amer Nubgan
Adv. life sci., vol. 10, no. 4, pp. 632-638, December 2023
*- Corresponding Author: Amer Nubgan (amer.nubgan@sc.uobaghdad.edu.iq)
Authors' Affiliations
[Date Received: 03/12/2023; Date Revised: 26/12/2023; Date Published: 31/12/2023]
Editorial Note: You are viewing latest version of this manuscript which is different from the originally published article.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: The Epstein-Barr virus (EBV) relates to the torch virus family and is believed to have a substantial impact on mortality and perinatal events, as shown by epidemiological and viral studies. Moreover, there have been documented cases of EBV transmission occurring via the placenta. Nevertheless, the specific location of the EBV infection inside the placenta remains uncertain.
Methods: The genomic sequences connected to the latent EBV gene and the levels of lytic EBV gene expression in placental chorionic villous cells are examined in this work. A total of 86 placentas from patients who had miscarriage and 54 placentas from individuals who had successful births were obtained for analysis.
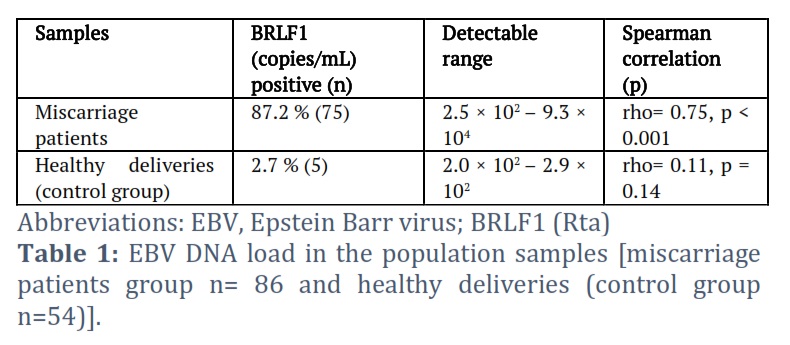
Results: The research employed QPCR to detect the BRLF1 (Rta) EBV lytic gene and quantify DNA burden in miscarriage patients and controls. In miscarriage patients, endpoint PCR and Sanger sequencing validated a particular region of the EBER1 EBV latent gene. BRLF1 gene presence ranged from 2.5 × 102 to 9.3 × 104 copies/mL in 75 miscarriage patients. However, a sample of 5 people with healthy deliveries showed a range of 2.0 × 102 to 2.9 × 102 copies/mL. All miscarriage samples were observed EBER1. The sequencing results indicated full sequence identity to EBV strains.
Conclusion: The detection of EBV gene expression in placental tissues in Iraq is a novel finding. The examination of EBV is of utmost importance in pregnant women who are experiencing severe illness, since it has the potential to lead to mortality in both the mother and the developing fetus.
Keywords: Epstein-Barr virus (EBV); Placental tissues; Prenatal; BRLF1; EBV-encoding RNA-1 (EBER1)
Introduction![]()
Numerous infectious pathogens that could harm the fetus are exposed to pregnant women. When these infectious agents enter an embryo or fetus, they typically result in one of two outcomes: either severe congenital defects or early embryonic and fetal mortality, which can lead to miscarriages; long-term effects can also include developmental issues. They may also cause severe neonatal and/or perinatal illness or interfere with fetal growth, leading to intrauterine growth restriction [1,2]. The herpesvirus family includes the widespread EBV, One of the most widespread viruses identified in humans [3,4]. It is well known that it has a preference for B lymphocytes and particular kinds of epithelial cells [5,6]. Over ninety-five percent of individuals all over the globe have an EBV infection that is dormant in their B cells. Getting EBV on B cells as a child is linked to several diseases in humans, such as infectious mononucleosis, various types of malignant lymphomas, lymphoproliferative disorders, and systemic autoimmune diseases [7–10]. More than 86 open reading frames may be found in the genome of the EBV double-stranded DNA, which is around 170 kilobytes in size. The virus genome has terminal repeats (TR) and four significant internal repeats (IR1 to IR4). Nine latent proteins are encoded by genes located in certain parts of the genome. These are latent membrane protein 1 (LMP1) and LMP2A, -2B, EBV nuclear antigen 1 (EBNA1), EBNA2, -3A, -3B, and -3C, and EBNA-LP [11]. Different lytic proteins with different functions are also encoded by other open reading frames, along with transcription factors and capsid proteins such as (BRLF1, BZLF1, BALF2, BNLF2a, BcLF-1, BPLF1, BILF1, and BNLF2a) [12]. To make more EBV RNAs that don’t code for proteins, the virus also makes microRNAs from BART and BHRF-1. These include EBV-encoded small RNAs 1 and 2 [13].
The life cycle of EBV includes two distinct phases: lytic replication and latency. It uses two different gene classes, each of which performs a very different function during its life cycle [14,15]. Many viruses exploit host cell manipulation-specifically, changes in cell cycle progression-as a means of accelerating their replication. Cells enter the S phase when exposed to little DNA tumor viruses. Herpesviruses might stop cells at any stage in the cell cycle, while small DNA tumor viruses can prompt cells to enter the S phase [16–19]. BRLF1 is also unknown as a replication transcription activator (Rta). It is a factor that controls the transcription that EBV expresses during the initial, early stage of the lytic cycle. Within two hours of the administration of an inducing stimulus, BRLF1 encodes and expresses it. The promoter just upstream of the BRLF1 ORF regulates the expression of the 4.0-kb mRNA that codes for the BRLF1 gene [20,21]. Rta is recognized to perform a wide range of biological tasks. The Rta-responsive element (RRE), a GC-rich motif presents in the viral promoters of various EBV lytic genes, including BMRF1, BMLF1, and BALF2, is where Rta attaches. Rta, however, also stimulates the transcription of viral genes whose promoters do not contain an RRE. For example, Rta indirectly turns them on by supporting the PI3-K and p38 mitogen-activated protein kinase-connected cell-signaling pathways. Rta may potentially use the regulation of E2F1 to activate certain viral genes [22–24]. Rta encourages growth arrest and starts a cellular senescence programme [25,26]. It is likely that Rta’s impacts on the host cell environment increase the effectiveness of lytic virus replication. Numerous viruses use host cell environment manipulation, specifically targeting cell cycle progression, to establish the ideal circumstances for viral reproduction [27,28]. The most prevalent EBV transcripts in EBV-associated tumors are the short RNAs that EBV encodes (EBERs, EBER1 and EBER2) [29,30]. It has long been assumed different EBV isolates have a common set of EBER coding sequences. Therefore, the connection between EBV-associated illnesses and mutations in the EBER genes has received minimal study [31].
Scientific investigation and debate have focused on the link between EBV infection and a number of pathologic diseases that can occur during pregnancy, including depression, pre-eclampsia, abortion, and stillbirth. Although several studies have revealed a possible connection, the proof is not yet conclusive [32–34]. The current study examined the expression of the BRLF1 (Rta) gene to identify the EBV infection in the placenta and connect it with abortion reasons in Baghdad, Iraq. Furthermore, we investigated sequence variation in the EBER gene, the reliable method for identifying latent EBV infection in tissues and contrasted our results with those from earlier studies. This work is the first description of the expression patterns of lytic EBV genes in placental tissue, namely in chorionic villous cells. Our research also demonstrates the connection between EBV infection and EBV-related abortions.
Methods![]()
Study population
In the present cohort research, placental tissues were collected from 86 female miscarriage patients (patients’ group), whose ages ranged from 20 to 41 years, and 54 placental tissues from healthy deliveries (control group). Samples obtained from the teaching laboratories at AL-Alwaiya Maternity Hospital in Baghdad, Iraq, between June – August 2023 were selected for this research. This study was approved by the Human and Animal Research Ethics Committee of the University of Baghdad (reference No. CSEC/0923/0060). All participants received verbal explanations of the study’s methodology and goals before reading the consent form and signing it.
DNA extraction and sample preparation
Placentas were kept chilled at 4°C from the moment of delivery and processed for DNA as soon as they arrived at the laboratory. 25-50 mm3 in volume of placental tissue (chorionic villous) samples were collected and fresh frozen at -80°C for 2 hours before being proceeded for DNA extraction. During the pre-treatment phase, the tissue was homogenised using proteinase K and ATL tissue lysis buffer at 56°C. DNA samples were extracted using the New iGNatal Kit (iGNatal Bioteh, Spain) according to the manufacture’s protocol. A NanoDrop ND-1000 spectrophotometer (Thermo Scientific, USA) was used to assess the DNA quantity and quality.
Assessment of viral loads by qPCR
For PCR amplification, a primer set targeting the highly conserved BRLF1 (Rta) EBV lytic locus, gene: BRLF1-FOR 5`-GCACATCTGCTTCAACAGGA-3` (Forward) and BRLF1-REV 5`-TACAAGAATCGGGTGGCTTC-3` (Reverse) gene bank accession NC_007605.1 (89838-93925), was utilized to increase detection EBV lytic rates and decrease false negative results. In lytic infection, BRLF1 (Rta) is a crucial regulatory gene that cooperates with BZLF1 to start the lytic phase. It contributes to the lytic cycle's driving force by triggering the expression of several lytic genes [35]. As a “house-keeping” gene, the β-actin gene: Actin-FOR 5’- CACCTTCTACAATGAGCTGCGTGTG-3’ (Forward) and Actin-REV 5’-ATAGCACAGCCTGGATAGCAACGTAC-3’ (Reverse) was utilized to check for DNA integrity and control for PCR inhibitors [36]. Prior to the EBV QPCR, beta-actin PCR was performed on all samples. DNase/RNase free water (no template DNA) controls were included to monitor contamination.
Each QPCR reaction mixture was prepared in a 200 μl flat capped tube (PCR-certified colorless) (Thermo Fishier Scientific, USA) to a final 12 μl volume, including 6 μL of 2X iTaq Universal SYBR Green Supermix (BioRad, USA), 4 μL of DNA elute, 0.25 μL (10 µM) of sense and antisense primers (Macrogen, South Korea), and 1.5 μL of DNase/RNase free water. Each primer set underwent a QPCR cycling profile with two steps. After an initial activation/denaturation stage of 13 minutes at 98°C, followed by 40 cycles were performed, each involving of the following: 15 seconds at 94°C, followed by 30 seconds optimised annealing temperature at 60°C using Rotor-Gene Quantitative Real Time Analyzer (QIAGEN, USA). Rotor-Gene software was used to analyse the QPCR readings. Between 55°C and 95°C, melt curves were measured at 0.5°C intervals. The cycle threshold (Ct) was set at 0.03 levels during the exponential amplification phase. DNA samples with no identifiable target DNA had a load of zero and were classified as negative.
The viral load calculations were evaluated based on the number of replicates tested, the final elution volume, and the
EBV identification and Sanger sequencing
Fifteen placental aborted samples derived from population carriers underwent PCR amplification and subsequent Sanger sequencing to confirm the EBV region of the EBV-encoded small RNA 1 (EBER1) (coordinates 6776-7397). The sense primer (EBER1-FOR) and antisense primer (EBER1-REV) used for the endpoint PCR amplification are 5`-GTCTTCGGTCAAGTACCA-3` and 5`-GTAGAGTGGGAGTGCTATC-3`, respectively [11]. 20 ng of gDNA was added to a 50 μL reaction mixture, containing 5 μL each of 10 μM forward and reverse primers and 25 μL of 2X Phusion High-Fidelity DNA Polymerase (Thermo Scientific, USA). Conventional PCR was performed in a T100TM Thermocycler (Bio-Rad, USA). The PCR condition was setup as follows: an initial DNA denaturation step at 98°C for 30 seconds, followed by 35 cycles at the temperatures: 98°C for 10 seconds, 59°C for 15 seconds, 72°C 15 seconds, followed by 72°C for 5 minutes.
All sequences were matched with the B95-8 prototype sequence, which was found in the GenBank database under the accession number V01555. In order to mitigate the potential impact of PCR-induced mismatched bases, the amplification and sequencing of samples containing mutations were repeated. The alignment of all EBER1 sequences was conducted using the programme Geneious 9.0.4.
EBV DNA verification
Direct gel analysis was carried out to find amplified PCR products. The amplification result was run on a 2 % agarose gel using a 10 µL sample (PCR amplicon). PCR amplicon was stained with ethidium bromide (0.5 µg/mL), and then it was examined using a GeneFlash UV transilluminator (Syngene, Cambridge, UK).
Statistical analysis
The GraphPad (Prism) software programme (version 5) was used. The correlations between the EBV DNA load in the miscarriage patients and healthy deliveries were evaluated using Spearman's (rho) correlation coefficient. P values below 0.05 were taken into consideration while analysing the results.
Results![]()
EBV lytic rates are strongest associated with aborted women
In the miscarriage patients’ group and the control group, BRLF1 DNA was detected in 87.2 % (75) and 2.7 % (5) of the samples, respectively (Table 1). In both sample groups, viral loads were detectable in the placental tissues at levels ranging from 2.0 × 102 to 9.3 × 104 copies/mL in both groups. Of the 75 BRLF1 DNA-positive miscarriage patient samples, 85.2% were detectable between 5.5 × 102 to 9.3 × 104 copies/mL. However, of the 5 BRLF1 DNA-positive control group samples, 80% were detectable between 2.0 × 102 to 2.5 × 102 copies/mL.
Endpoint PCR and genomic analysis of EBV isolated from placental tissues
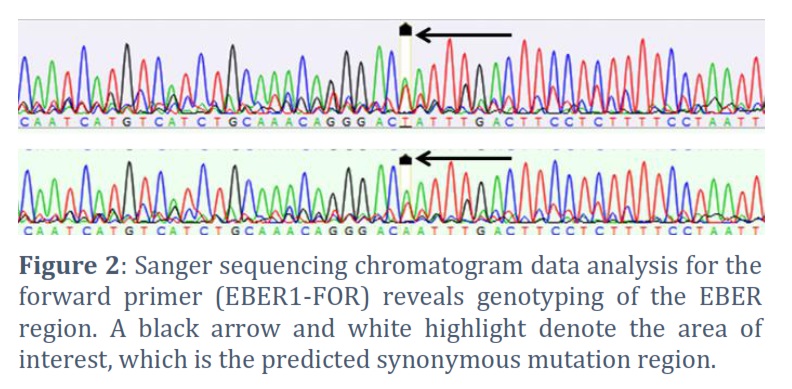
Miscarriage DNA samples were used for endpoint PCR amplification and Sanger sequencing to confirm a region of the EBER haplotype (coordinates 6776-7397). The endpoint PCR findings revealed that all samples tested positive for EBER1 (Figure 1). Positive identification, the electrophoretic sizes of the PCR products were identical to the sizes that were expected based on the annotated sequencing. This confirmed that EBV was present in the placental tissues.
Fifteen of the PCR amplicon samples were exposed to commercial Sanger sequencing to confirm the PCR results. Figure 2 displays a chromatogram of sequencing data that represents various samples. We compared our results with globally published EBV genomes because this work takes into account the first sequencing of EBV genomes isolated from patients in Iraq. It was possible to directly check the PCR products by comparing them to herpesvirus genomic DNA sequences from the National Library of Medicine/National Institutes of Health. The 653-base pair fragments were amplified using endpoint PCR to target EBER1 region, specifically covering coordinates 6776 to 7397. A single-nucleotide change was identified by comparative analysis with the EBV reference sequence B95-8 (No. V01555) and other spontaneous alterations (synonymous mutations). Of fifteen sequenced samples, only two were revealed synonymous mutations from T to A. Interestingly, the viral loads for these two samples (mismatch sequences) were below 2.8 × 102 copies/mL. There was a weak association (not statistically significant) between BRLF1 DNA load and mismatch sequence (Spearman’s rho = 0.11, p = 0.11). No nucleotide deletions, insertions, or premature stop codons were observed in the investigated versions of EBER1, indicating a lack of evidence for such genetic alterations.
Figures & Tables

Genital HSV can cause a serious neonatal condition, particularly in primary infections, which puts the newborn at risk if infected during birth. Even though they don't happen very often, intrauterine infections can cause abortion or stillbirth, skin scars (called cutaneous symptoms), eye problems (like chorioretinitis and microphthalmia), and neurological problems (like brain damage) [1,38,39]. The EBV-encoded BRLF1 (Rta) and EBER1 genes, which trigger the lytic cycle, and the EBER1 gene, which participates in the interaction between the virus and its host cells, respectively, during virus infection, have been verified in this study as an indicator for lytic and latent virus infections [40,41]. The findings of the current investigation reveal a noteworthy incidence of EBV in placental tissues. In this study, a total of 86 placental samples were examined to identify the presence of the BRLF1 gene associated with EBV using the quantitative polymerase chain reaction (QPCR) method. The results revealed that 75 out of the 86 samples, accounting for 87.2 %, tested positive for the presence of the BRLF1 gene (which serves as an indicator for EBV lytic replication). The aforementioned high proportion indicates a significant prevalence of EBV infection in the placental tissues being examined.
EBV has the potential to induce placental infection during pregnancy, leading to subsequent difficulties for the developing fetus. During pregnancy, it is possible for both primary and secondary EBV infections to take place. Nevertheless, initial infections with EBV by transplacental transmission are few, although secondary maternal EBV infections are often seen [1]. Since more than two decades the nucleic acid testing (NAT) techniques have been more widely available, quantifying EBV DNA in blood has shown to be helpful in both monitoring and diagnosing as Burkitt’s lymphoma (BL), chronic active EBV infection, nasopharyngeal carcinoma (NPC), infectious monoclonal (IM), EBV infection in HIV-positive patients, and posttransplant lymphoproliferative disorder (PTLD) [42–49].
According to the study conducted by Avgil and Ornoy, the findings indicate that in 22.5% of instances (9 out of 40), there was observed nuclear localization of EBV-Viral Capsid Antigen (VCA) expression. This finding indicates that a significant proportion of EBV infections occurring in placental tissues during pregnancy exhibited a degree of severity ranging from mild to severe [1,50].
The findings of the current study suggest QPCR method, as a high sensitivity and precision in determining the EBV viral nucleic acids. We employed the QPCR technique here in order to determine the EBV lytic cycle by referring to the BRLF1 gene. However, this is contradiction to the research that was performed by Avgil and Ornoy [1]. Wang and his research group revealed the remarkable conservation noticed in the BHRF-1 gene through various clinical samples connected to the EBV. A study revealed that BHRF1 is considered as one of the highly conserved gene, therefore the direct mutations in this gene may not have a direct correlation with disorders connected to EBV infections [40]. These results are considered crucial for our current hypothesis in related to identifying EBV infection by looking for the BHRF-1 gene. By looking on a gene that is known for its constancy and preservation in EBV linked circumstances, we may expose elusive details in the detecting of EBV process. There is slight known about the relations between EBER1 mutations and the progression of illnesses connected with EBV. Furthermore, there is currently a lack of published data relating to the alterations of the EBER1 gene across numerous terrestrial counties. It’s crucial and time-sensitive to highlight and find solution these issues. By looking to the clinical data gained from Shandong State in North China, it has been detected that EBER1 mutations are not connected to the progression of leukemia and myelodysplastic syndrome (MDS) [31]. The preservation of coding sequences between various EBV variants in EBERs has been a long-lasting certainty. Therefore, the study of mutations happening in the EBER genes and their association with disorders linked with EBV has received little research attention [31,41]. The most predominant transcripts in tumors related with EBV are EBERs [29,30]. Mutations in these genes have the likely to influence its biological roles, carcinogenic belongings, and the immunological reactions of the host. Further examination is desirable to conclude the association between mutations in the BHRF-1 gene and EBV- related disorders across diverse geographical localities.
To assess the probable clinical consequences of EBV infection in this matter and to better comprehend the connection between EBV and the placenta during pregnancy, more research is required. In order to improve our knowledge of the virus's participation in pregnancy and its possible impact on fetal progress, more examination is vital. The results of the current study highlight the probable effect of EBV on the wellbeing of the placenta and warrant the need for more investigation to search the concerns of EBV infection in the course of pregnancy. The knowledge of the rate of EBV in placental tissues improves our understanding of viral dynamics in the context of generative well-being, theoretically impacting the health and well-being of both the mother and the fetus. It is worthwhile to carry out more surveys and in depth study in order to investigate the unique processes and implications of EBV infection during of pregnancy.
Acknowledgment
We express our gratitude to the teaching laboratory staff at AL-Alwaiya Maternity Hospital in Baghdad, Iraq, for their valuable assistance in acquiring samples. Additionally, we extend our appreciation to the staff members at the Department of Biology, College of Science, University of Baghdad, for their generous support in conducting this study.
Funding Statement
The researcher states that no grants or other funds were gained during the conducting of this manuscript.
Ethical Declaration
This investigation was permitted by the HAREC of University of Baghdad (register No. CSEC/0239/0006). All contributors gained oral clarifications of the study's approach and aims upon impression the agreement form and signing it.
The authors declare no conflict of interest.
![]() References
References
- Avgil M, Ornoy A. Herpes simplex virus and Epstein-Barr virus infections in pregnancy: consequences of neonatal or intrauterine infection. Reproductive Toxicology, (2006); 21(4): 436–445.
- W. Ahmed D. Effects of Interleukin-2 (IL-2) and Interleukin-6 (IL-6) in Recurrent Spontaneous Abortion (RSA). Iraqi Journal of Pharmaceutical Sciences, (2017); 17(2): 74–79.
- Chang MS, Lee HS, Kim CW, Kim Y Il, Kim WH. Clinicopathologic Characteristics of Epstein-Barr Virus-Incorporated Gastric Cancers in Korea. Pathology – Research and Practice, (2001); 197(6): 395–400.
- Fleisher G, Henle W, Henle G, Lennette ET, Biggar RJ. Primary Infection with Epstein-Barr Virus in Infants in the United States: Clinical and Serologic Observations. Journal of Infectious Diseases, (1979); 139(5): 553–558.
- Borza CM, Hutt-Fletcher LM. Alternate replication in B cells and epithelial cells switches tropism of Epstein–Barr virus. Nature Medicine, (2002); 8(6): 594–599.
- Lafta FM, AL-Jumaily RMK, Rasoul LM. Global DNA Methylation Levels in Epstein-Barr-Virus-Positive Iraqi Patients with Acute Lymphoblastic Leukaemia. Iraqi Journal of Science, (2023); 64(3) : 1109–1118.
- Draborg AH, Duus K, Houen G. Epstein-Barr Virus in Systemic Autoimmune Diseases. Clinical and Developmental Immunology, (2013); 2013: 535738.
- Mousa Al-Mousawi HT, AL-Hajjar QN. Immunological and Molecular Diagnosis of Cytomegalovirus Infection between Aborted & Pregnant Women in Babylon City. Baghdad Science Journal, (2021); 18(2): 1086.
- Ali SHM, Mohammed KIA, Ali WM, AL-Fakhar SA, Al-Alwany SHM, Mousa JM. In Situ Hybridization for Molecular Detection of Human Papilloma Viral 6 / 11 DNA in Adenoctomized Tissues from A group of Iraqi Pediatric Patients. Baghdad Science Journal, (2022); 19(1): 0026.
- Okano M, Gross TG. Acute or Chronic Life-Threatening Diseases Associated With Epstein-Barr Virus Infection. The American Journal of the Medical Sciences, (2012); 343(6): 483–489.
- Hui KF, Chan TF, Yang W, Shen JJ, Lam KP, et al. High risk Epstein-Barr virus variants characterized by distinct polymorphisms in the EBER locus are strongly associated with nasopharyngeal carcinoma. International Journal of Cancer, (2018); 2(144): 3031–3042.
- Yap LF, Wong AKC, Paterson IC, Young LS. Functional Implications of Epstein-Barr Virus Lytic Genes in Carcinogenesis. Cancers, (2022); 14(23): 5780.
- Swaminathan S. Noncoding RNAs produced by oncogenic human herpesviruses. Journal of Cellular Physiology, (2008); 216(2): 321–326.
- Hwang SP, Huang LC, Wang WH, Lin MH, Kuo CW, et al. Expression of Rta in B Lymphocytes during Epstein–Barr Virus Latency. Journal of Molecular Biology, (2020); 432(19): 5227–5243.
- Klein G. Epstein-Barr virus strategy in normal and neoplastic B cells. Cell, (1994); 77(6): 791–793.
- Turkey EF, Hamad AG, AL-Mansoori AK. The Association of Prothrombin Gene Mutations and Cytomegalovirus Infection with Abortion Among Iraqi Women. Baghdad Science Journal, (2022); 19(4): 0768.
- Song B, Liu JJ, Yeh K-C, Knipe DM. Herpes Simplex Virus Infection Blocks Events in the G1 Phase of the Cell Cycle. Virology, (2000); 267(2): 326–334.
- Ehmann GL, McLean TI, Bachenheimer SL. Herpes Simplex Virus Type 1 Infection Imposes a G1/S Block in Asynchronously Growing Cells and Prevents G1 Entry in Quiescent Cells. Virology, (2000); 267(2): 335–349.
- Tiwari D, Mittal N, Jha HC. Unraveling the links between neurodegeneration and Epstein-Barr virus-mediated cell cycle dysregulation. Current Research in Neurobiology, (2022); 30(3): 100046.
- Packham G, Brimmell M, Cook D, Sinclair AJ, Farrell PJ. Strain Variation in Epstein-Barr Virus Immediate Early Genes. Virology, (1993); 192(2): 541–550.
- Zhang JB, Huang SY, Wang TM, Dong SQ, He YQ, et al. Natural Variations in BRLF1 Promoter Contribute to the Elevated Reactivation Level of Epstein-Barr Virus in Endemic Areas of Nasopharyngeal Carcinoma. EBioMedicine, (2018); 37: 101–109.
- Darr CD, Mauser A, Kenney S. Epstein-Barr Virus Immediate-Early Protein BRLF1 Induces the Lytic Form of Viral Replication through a Mechanism Involving Phosphatidylinositol-3 Kinase Activation. Journal of Virology, (2001); 75(13): 6135–6142.
- Liu C, Sista ND, Pagano JS. Activation of the Epstein-Barr virus DNA polymerase promoter by the BRLF1 immediate-early protein is mediated through USF and E2F. Journal of Virology, (1996); 70(4): 2545–2555.
- Myers JE, Schaal DL, Nkadi EH, Ward BJH, Bienkowska-Haba M, Sapp M, Bodily JM, Scott RS. Retinoblastoma Protein Is Required for Epstein-Barr Virus Replication in Differentiated Epithelia. Banks L, ed. Journal of Virology, (2023); 97(2): e0103222
- Ragoczy T, Heston L, Miller G. The Epstein-Barr Virus Rta Protein Activates Lytic Cycle Genes and Can Disrupt Latency in B Lymphocytes. Journal of Virology, (1998); 72(10): 7978–7984.
- Heilmann AMF, Calderwood MA, Portal D, Lu Y, Johannsen E. Genome-Wide Analysis of Epstein-Barr Virus Rta DNA Binding. Journal of Virology, (2012); 86(9): 5151–5164.
- Ho JWY, Li L, Wong KY, Srivastava G, Tao Q. Comprehensive Profiling of EBV Gene Expression and Promoter Methylation Reveals Latency II Viral Infection and Sporadic Abortive Lytic Activation in Peripheral T-Cell Lymphomas. Viruses, (2023); 15(2): 423.
- Abbas MD, Egbe SS. Seroprevalence of CMV in Women with Bad Obstetric History in Babil/Iraq. Iraqi Journal of Pharmaceutical Sciences, (2021); 30(2): 106–112.
- Chen WG, Chen YY, Bacchi MM, Bacchi CE, Alvarenga M, Weiss LM. Genotyping of Epstein-Barr virus in Brazilian Burkitt’s lymphoma and reactive lymphoid tissue. Type A with a high prevalence of deletions within the latent membrane protein gene. The American journal of pathology, (1996); 148(1): 17–23.
- Yao QY, Rowe M, Martin B, Young LS, Rickinson AB. The Epstein–Barr virus carrier state: dominance of a single growth-transforming isolate in the blood and in the oropharynx of healthy virus carriers. Journal of General Virology, (1991); 72(7): 1579–1590.
- Wang HY, Sun L, Li P, Liu W, Zhang ZG, Luo B. Sequence Variations of Epstein-Barr Virus-Encoded Small Noncoding RNA and Latent Membrane Protein 1 in Hematologic Tumors in Northern China. Intervirology, (2021); 64(2): 69–80.
- Zhu P, Chen YJ, Hao JH, Ge JF, Huang K, et al. Maternal depressive symptoms related to Epstein-Barr virus reactivation in late pregnancy. Scientific Reports, (2013); 3(1): 3096.
- Elliott SE, Parchim NF, Kellems RE, Xia Y, Soffici AR, Daugherty PS. A pre-eclampsia-associated Epstein-Barr virus antibody cross-reacts with placental GPR50. Clinical Immunology, (2016); 168: 64–71.
- Tomai X. Stillbirth following severe symmetric fetal growth restriction due to reactivation of Epstein–Barr virus infection in pregnancy. Journal of Obstetrics and Gynaecology Research, (2011); 37(12): 1877–1882.
- Long X, Yang J, Zhang X, Yang Z, Li Y, Wang F, Li X, Kuang E. BRLF1 suppresses RNA Pol III‐mediated RIG‐I inflammasome activation in the early EBV lytic lifecycle. EMBO reports, (2021); 22(1): e50714.
- Nubgan A. The deubiquitylase-ubiquitin-specific protease 4 absence in HeLa cells leads to a reduction in Semliki Forest virus replication. Human Gene, Elsevier, (2023); 36: 201170.
- Hakim H, Gibson C, Pan J, Srivastava K, Gu Z, Bankowski MJ, Hayden RT. Comparison of Various Blood Compartments and Reporting Units for the Detection and Quantification of Epstein-Barr Virus in Peripheral Blood. Journal of Clinical Microbiology, (2007); 45(7): 2151–2155.
- Saleh DS, Kharibet KE. Detection of Cytomegalovirus, Rubella virus, and IL-2 Levels in a Sample of Recurrently Aborted Iraqi Women. Iraqi Journal of Science, (2015); 56(3A): 1890–1894.
- Zekri ARN, Bahnassy AA, Mohamed WS, El-Kassem FA, El-Khalidi SJ, et al. Epstein-Barr virus and breast cancer: Epidemiological and Molecular study on Egyptian and Iraqi women. Journal of the Egyptian National Cancer Institute, (2012); 24(3): 123–131.
- Wang Y, Rong Y, Yang L, Lu Z. Genetic variability and mutation of Epstein‒Barr virus (EBV)-encoded LMP-1 and BHRF-1 genes in EBV-infected patients: identification of precise targets for development of personalized EBV vaccines. Virus Genes, (2023); 59(4): 541–553.
- Abd Al Kareem RM, Abd WS. Impact of EBV on Multiple in a Sample of Iraqi Females: Immunological and Molecular Study. Iraqi Journal of Science, (2020); 61(5): 1008–1015.
- Rowe DT, Qu L, Reyes J, Jabbour N, Yunis E, Putnam P, Todo S, Green M. Use of quantitative competitive PCR to measure Epstein-Barr virus genome load in the peripheral blood of pediatric transplant patients with lymphoproliferative disorders. Journal of Clinical Microbiology, (1997); 35(6): 1612–1615.
- Van Esser JWJ, Niesters HGM, Thijsen SFT, Meijer E, Osterhaus ADME, et al. Molecular quantification of viral load in plasma allows for fast and accurate prediction of response to therapy of Epstein–Barr virus‐associated lymphoproliferative disease after allogeneic stem cell transplantation. British Journal of Haematology, (2001); 113(3): 814–821.
- Chan KCA, Zhang J, Chan ATC, Lei KIK, Leung SF, et al. Molecular characterization of circulating EBV DNA in the plasma of nasopharyngeal carcinoma and lymphoma patients. Cancer research, (2003); 63(9): 2028–32.
- Lin JC, Wang WY, Chen KY, Wei YH, Liang WM, et al. Quantification of Plasma Epstein–Barr Virus DNA in Patients with Advanced Nasopharyngeal Carcinoma. New England Journal of Medicine, (2004); 350(24): 2461–2470.
- Yamamoto M, Kimura H, Hironaka T, Hirai K, Hasegawa S, et al. Detection and quantification of virus DNA in plasma of patients with Epstein-Barr virus-associated diseases. Journal of Clinical Microbiology, (1995); 33(7): 1765–1768.
- Kimura H, Morita M, Yabuta Y, Kuzushima K, Kato K, et al. Quantitative Analysis of Epstein-Barr Virus Load by Using a Real-Time PCR Assay. Journal of Clinical Microbiology, (1999); 37(1): 132–136.
- Stevens SJC, Blank BSN, Smits PHM, Meenhorst PL, Middeldorp JM. High Epstein–Barr virus (EBV) DNA loads in HIV-infected patients: correlation with antiretroviral therapy and quantitative EBV serology. AIDS, (2002); 16(7): 993–1001.
- Stevens SJC, Pronk I, Middeldorp JM. Toward Standardization of Epstein-Barr Virus DNA Load Monitoring: Unfractionated Whole Blood as Preferred Clinical Specimen. Journal of Clinical Microbiology, (2001); 39(4): 1211–1216.
- Ad’hiah AH, Atiyah NS, Fadhil HY. Qualitative and Quantitative Molecular Analysis of Epstein-Barr Virus in Iraqi Patients with Relapsing-Remitting Multiple Sclerosis. Iraqi Journal of Science, (2023); 64(1): 127–137.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


