

Full Length Research Article
Capability of Trichoderma viride to Produce Cellulolytic and Pectolytic Enzymes
Abdel Moneim Elhadi Sulieman1*, Siham M. Hakim2, Wasima Alshammari1, Nawaf I. Alshammari1, Zakaria A. Salih3
Adv. life sci., vol. 10, no. 3, pp. 491-496, September 2023
*- Corresponding Author: Abdel Moneim Elhadi Sulieman (am.sulieman@uoh.edu.sa)
Authors' Affiliations
2. Center of Biosciences and Biotechnology, Faculty of Engineering and Technology, University of Gezira, Wad-Medani – Sudan
3. Department of Research and Training, Research and Training Station, King Faisal University, Al-Ahsa – Saudi Arabia
[Date Received: 29/06/2023; Date Revised: 21/08/2023; Date Published: 30/09/2023]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Species of the genus Trichoderma have been used in the food and textile industries to produce cellulases and other enzymes that degrade complex polysaccharide structures. Trichoderma species have been utilized to make cellulases and other enzymes that break down intricate polysaccharide structures in the food and textile industries. The study investigated the nutritional requirements and the production of enzymes by the fungus Trichoderma viride fungus' ability to produce enzymes).
Methods: We used a medium supplemented with cellulolytic and pectic substances for enzyme production.
Results: The results of the study proved that. Methionine, Glutamic acid, and leucine effectively enhanced mycelial growth. Findings regarding impact of pH level on the development of the fungus T. viride indicated that the maximum growth was at pH 5.0. However, growth decreased dramatically with increasing pH values. We examined the power of the fungus T. viride to produce cellulolytic and pectolytic enzymes in various substrates in the current study.
Conclusion: The findings demonstrated that Pectin was the best substrate for pectolytic enzyme synthesis, whereas Carboxymethyl cellulose (CMC) was the best substrate for the cellulolytic enzyme.
Keywords: Carboxymethyl cellulose; Rhizosphere; Digestibility; Hemicellulases
Introduction![]()
Trichoderma filamentous fungi of the rhizosphere constitute a significant component of diverse soil ecosystem microbiomes and are distinguished by their capacity to penetrate plant roots. Trichoderma's employment in agriculture can be effectively ensured by thoroughly understanding its characteristics, including metabolic activity and the nature of interactions with plants and other microorganisms [1]. T. harzianum and T. viride are the two most significant species in the genus. The genus species are adaptable fungi used in various commercial and industrial applications. In addition to many bioactive metabolites against NTCD, fungi also produce colors, antioxidants, polysaccharides, and enzymes that may be investigated as natural alternatives to synthetic food additives. Through better fermentation conditions, metabolic engineering techniques, and fungi genetic manipulation, research in this area has enhanced the yields of metabolites for industrial purposes [2].
Trichoderma is frequently found as an indoor pollutant and has been shown to make up a sizeable portion of the fungal biomass in soil [3, 4, 5]. Trichoderma is frequently found in soil and woody materials. Due to its potent disintegrating function as spoilage organisms and for use in various plans for degrading cellulose waste, it has recently attracted significant economic interest [6].
Fungi produce cellulases and other enzymes that break down complex polysaccharide structures, and they have been exploited in the food and textile industries. Many extracellular enzymes are produced by them quite effectively. They are employed commercially in synthesizing enzymes like cellulases that can digest tough polysaccharides. These applications are typical in the businesses related to both food and textiles. The enzymes are also used in poultry feed to increase the digestion of hemicelluloses from barley or other crops [7].
According to Wang et al. [8, During their aggressive action against plant pathogens, Trichoderma species exhibit five mechanisms: antibiosis via secondary metabolite production; myco-parasitism via hydrolytic enzyme secretion; competition for space and nutrients; promotion of plant growth; and induction of systemic resistance in the host plant.
It is possible to add additional enzymatic capabilities and expand the applications of a selective consortium of fungal strains to deconstruct biomass into sugar [9, 10]. Co-cultivating diverse species of fungi enabled the production of enzymatic extracts abundant in different kinds of enzymes [11, 12]. In their investigation, Romero-Peláez et al. [13] shown that co-cultures of the ascomycete T. reesei RUT-C30 and the basidiomycete P. lecome resulted in enzymatic extracts high in cellulases, ligninases, and hemicellulases, allowing for an increase in the hydrolysis yield of pretreated sugarcane bagasse.
Pectin depolymerization is frequently related to the ripening process of fruit. A particular group of pectic enzymes breaks down pectic enzymes produced by many plant pathogens. Due to their importance, these enzymes affect how fruits and vegetables are stored after harvest [14]. Numerous studies have been conducted on these enzymes.
Although many fungi can live on cellulose or create enzymes that break down amorphous cellulose, it should be stressed that only a few fungi can fully develop extracellular cellulase systems that can break down crystalline cellulose [15]. The most well-known species are Trichoderma and Phanerochaete [16].
Cellulosic enzyme production has been attributed to a variety of microorganisms. Trichoderma viride, Verticillium albo-atrum, Fusarium oxysporum, F. solani, and Stemphylium botryosum are among them [17]. Trichoderma spp. were thought to be essential contributors to the lyses of Rhizoctonia solani cells by producing cellulase and chitinase enzymes [18].
Cellulose, in its native form, is insoluble and partly crystalline. Cellulose conversion to glucose needs a complex of enzymes, initially designated as; C1, Cx, and cellobiase [19]. The Cx (endo B- 1,4 glucanase) or the carboxymethyl cellulase (CMCase) splits soluble cellulose (carboxy methyl cellulose) to produce glucose from cellobiose.
The pectolytic enzymes include three main groups of enzymes; the esterases (Pectinmethylesterases PME), the polygalacturonases (P.G.), and the lyases (pectin methyl trans-eliminates PTE). The pectin methyl esterases (PME) from bacteria (Clostridium multi-fermentans), and fungi (Botrytis cinerea, Corticium rolfsii, Fusarium oxysporum, and Phytophthora infestans) as was reported by Forster [20]. The lyases (pectin methyl trans-eliminates PTE) break the a-1,4 linkage through a trans-eliminate cleavage, accompanied by a simultaneous elimination of H from C-5. Microorganisms in natural settings are responsible for regulating enzyme production to ensure the best effective utilization of the carbon supply that is present [21]. The ultimate objective of the industry is to discover and make use of naturally occurring microorganisms that are capable of manufacturing the desired enzymes in large yields and with a high level of productivity. The purpose of this study was to evaluate several elements of the capacity of the fungus T. viride to produce the virus.
Methods![]()
The isolates of the fungus T. viride used in the present study were obtained from the Plant Pathology Laboratory, Agriculture Research Corporation, Wad Medani, Sudan. Five different carbohydrates (glucose, sucrose fructose, galactose, and mannose) and five amino acids (glutamic acid, aniline, Arginine, leucine, and methionine) were obtained from the Food Science and Technology Department Lab. The fungus was grown on PDA slants and stored at -200 C before being.
Media used: Potato Dextrose Agar (PDA)
This study used Oxoid compounds to make PDA media locally. Fungal growth was sustained on this media. After boiling 200 grams of peeled and diced potato in one liter of distilled water for an hour, the extract was filtered and made up to 1 liter. Steaming and guiding dissolved D-glucose and agar. Then 100 ml samples of the medium were dispensed in conical flasks with cotton plugs and aluminum foil and sterilized in the autoclave at 121°C (15 Ib/in2) for 15 minutes. It was refrigerated at 40°C. When needed, the medium was melted in a water bath and put into sanitized Petri dishes. For cellulase and pectic enzyme synthesis, the fungus was cultivated on basal salt medium with the following.
K2HPO4 (1.0 g)
KH2PO4 (0.5 g)
(NH4)2SO4 (1.0 g)
Substrate (10.0 g)
Distilled water 1000ml
Three 5.0 mm fungal discs from an actively growing fungus were injected into the medium after 15 minutes of autoclaving at 121°C, cooling, and incubating on a rotary incubator at room temperature. The following substrates produced the cellulase enzyme; Carboxymethylcellulose (CMC), cotton fibers, filter paper, and glucose. Nevertheless, for the production of the pectic enzyme, the following were used; Pectin, glucose, and orange peels.
Effects of amino acids on fungal radial growth
PDA was utilized for fungal radial growth[22]. Substituting medium D-glucose yielded five amino acids at varied amounts. Sterilized media was placed into Petri dishes and solidified at room temperature (28-30°C). A fungal growth disc from an actively developing T. viride culture on PDA was injected into each cemented Petri plate. Petri dishes were cultured at room temperature for eight days after inoculation. All treatments were tripled. Each disc in a Petri dish was averaged two crossed dimensions to measure growth diameter every 48 hours. Radial growth was calculated as a percentage of Petri dish diameter.
Effects of the amino acids on mycelial dry weight
PDB was made and distributed in 100 ml conical flasks (45 ml each). For each amino acid, 0.0, 25.0, 50.0, 75.0, and 100.0 concentrations were synthesized. Each solution was sterilized in an autoclave at 121°C (15 1b/in2) for 15 minutes and cooled to room temperature. A sterile cork-borer (5.0 mm diameter) formed three discs from an edge of an actively developing T. viride culture on a solidified PDA medium to inoculate each flask. For 12 days, inoculated plates were incubated at 28-30°C. Following incubation, the culture was filtered through Wittman's No. 1 filter papers and dried at 800 C for 24 hours before weighing. All treatments were tripled [23].
Effects of different pH levels on mycelial growth
Abubakar et al. [23] examined how pH affects mycelial growth. PDB medium was made and administered in 250 ml flask. Three 5.0 mm discs from an edge of an actively developing T. viride culture on solidified PDA media were injected into each flask. Samples pH was adjusted to be ranging from 4 – 9 pH (4, 5, and 6 were made by adding a concentrated HCl acid, while the pH values of 7, 8, and 9 were made by using NaOH solution). Three treatments were used for each pH level; the solutions were autoclaved at 121°C and cooled at room temperature (28 – 300°C). The flasks that were inoculated were placed in an incubator set at room temperature for a duration of 8 days. Following the incubation period, the cultures were subjected to filtration using a Whatman filter paper. The resulting mass was measured as the fresh weight (wt), after which it was subjected to a drying process at a temperature of 800°C for a duration of 24 hours. Subsequently, the dried mass was measured again as the dry weight (wt).
Production of cellulase and pectic enzymes
The fungus was grown on a basal salt medium containing the following constituents for cellulase and pectic enzyme production (prepared according to Nwodo-Chinedu et al [24]. The medium was autoclaved at 1210 C for 15 minutes, cooled, and inoculated by three fungal discs (5.0 mm diameter), cut from an actively growing fungus, and incubated on a rotary incubator at room temperature. The following substrates produced the cellulase enzyme; Carboxymethylcellulose (CMC), cotton fibers, filter paper, and glucose. However, Pectin, glucose, and orange peels were used for pectic enzyme production [25].
Enzyme assay: viscometrical method
Both the cellulase (Cx) and pectinase enzymes were assayed by the substrate viscosity reduction method using Viscometers (Cannon Fenke type) [26]. The viscometers were suspended in a water bath at 300 C. The reaction mixtures contained each of the followings:
Substrate (CMC or Pectin) 8.0 ml
Water 1.0 ml
CaCl2 0.01 ml
Enzyme 1.0 ml
Before any viscometer was used, it was checked against a standard of water. A 100% reduction in viscosity was defined as the time it took to blow 10 ml of water.
Results![]()
Effect of amino acids on radial growth
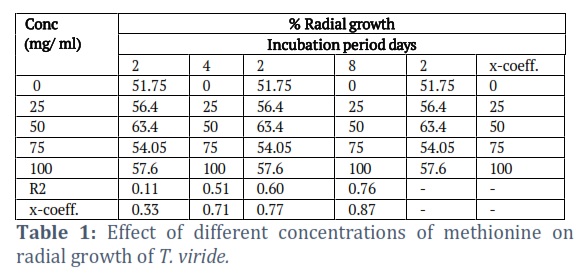
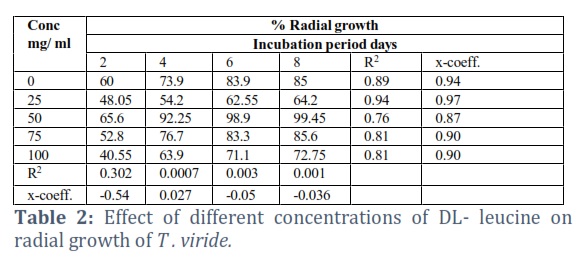
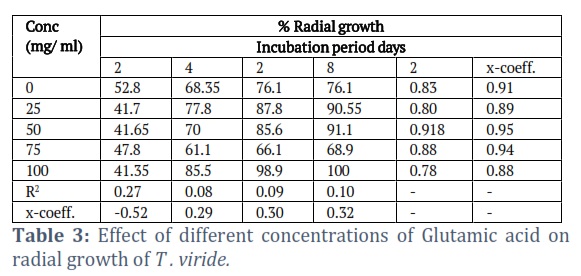
The effects of the different concentrations of the amino acids on the radial growth of the fungus were compared. The results showed that methionine was the most effective and gave significantly better results than all of the amino acids tested (Tables 1-3). It gave 100% radial growth at its higher concentration and the growth was also increased at its lower concentrations. Among the different amino acids tested, only Arginine was less effective in the radial growth of the fungus.
Methionine and leucine were also significantly better, giving 99.4 and 99.45% radial growth at 50 mg/ml concentrations. The glutamic acid has seemlier significant to the leucine. The better growth of glutamic acid at its higher concentration gave 100% radial growth (Table 3). The other amino acids (alanine and Arginine) were ineffective in enhancing the radial growth of the fungus, even at their higher concentrations. Although alanine was slightly better than the control treatment at 50 mg/ml, it is far less than the other amino acids, while Arginine gave less radial growth than the control.
Production of cellulosic enzymes
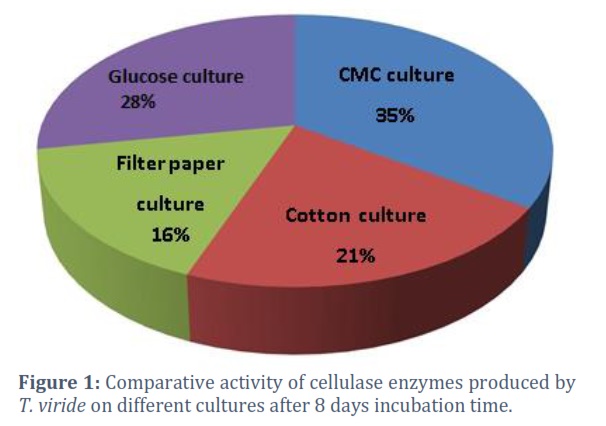
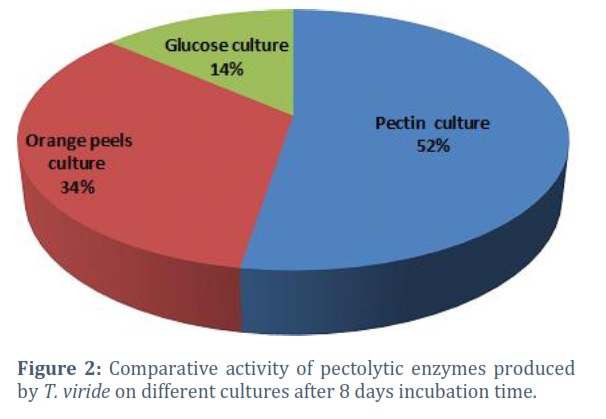
Culture media containing cellulose substrates (Carboxymethyl cellulose-CMC, cotton fibers, filter paper, and L-glucose) were inoculated with the fungus T. viride and incubated at room temperature (28- 30°C) for 24 days. The enzyme activity was measured every eight days using the viscosity-reducing method [27]. The comparative activity of the cellulosic enzyme at different incubation intervals (8,16, and 24 days) is shown in Fig. (1). Activity was found to increase with time. Carboxymethyl cellulose (CMC) was the best substrate for cellulase enzyme production, giving 35% of the comparative Activity. Other materials include L-glucose (dextrose), which gave 28%, then cotton fibers which gave 21%, and the minor comparative Activity was recorded for the filter paper culture T. viride was tested on cultures of three pectic substrates (Pectin, orange rind, and L-glucose). The enzyme activity was also measured by the viscosity-reducing method. The comparative Activity of the pectolytic enzyme at different incubation intervals (8, 16, and 24 days) and in different substrates is shown in Fig. (2). Pectin culture was found as the best one for pectolytic enzyme production, giving 52% comparative Activity. Followed by the orange rind, which gave 34%, while the lowest comparative Activity was recorded in the L-glucose cultures.
Figures & Tables
The degradation and poor soil quality due to intensive agriculture, lousy land management techniques, and resource extraction from the soil are major global concerns. Contamination, erosion, salinization, flooding, and biodiversity loss are risks linked to soil deterioration [28]. Degraded soils are characterized by a lack of available organic matter in the soil as well as a depleted pool of organic carbon, with losses ranging from 25–75 percent of the initial soil organic matter and organic carbon pool [29]. Loss of biological variety and functioning occurs together with soil degradation. Recent research has demonstrated that anthropogenic activities and climate change impair soil’s ability to carry out essential functions and processes such as primary production, nitrogen cycling, litter decomposition, and organic matter mineralization [30]. Most of the research under consideration reported that the rhizosphere had more microbial variety and richness than the control. The reason is that more nutrients and organic C substrates are available for mineralization [31].
According to Qiao et al.[32], the rhizosphere is a critical interface for interactions, signaling, and exchanging resources and energy between plants and soil microbes. This discovery demonstrates how these organisms often thrive in carbon-rich environments, which are prevalent in the rhizosphere, thanks to their high metabolic Activity, rapid development, and proliferative abilities. According to these results, O.A.s increase the supply of fresh organic materials, which widens niches, lessens competition, and fosters positive co-occurrence patterns with keystone saprotrophic species [8]. The rhizosphere also controls the dynamics of the C and N in the soil. The decomposition and transformation of organic compounds that were frequently more intense in the rhizosphere included cellulolysis, xylanolysis, ligninolytic, ureolysis, and chitinolysis, indicating the presence and Activity of more organisms that break down organic compounds [33].
Saprophytic fungi, including Trichoderma species significance in plant residue digestion and plant health, as well as the vast amount of microbial biomass they contribute to the soil, make them critical and typical components of the soil rhizosphere. [34, 35]. T. viride, the subject of the present study, was very important. Trichoderma species, being saprophytic, are adapted to thrive in diverse situations, producing a wide array of enzymes [36, 37]. Microbial pectolytic enzymes, on the other hand, are known to play a vital role in many industrial processes such as; fiber extraction, clarification and pectination of fruit juices, and extraction of vegetable oils [38]. The present study tested the ability of the fungus T. viride to produce both cellulolytic and pectolytic enzymes in different substrates. The results showed that Carboxymethyl cellulose (CMC) was the best substrate for the cellulolytic enzyme, while Pectin was the best for the pectolytic enzyme production [39] in a similar study also reported that CMC was the best substrate for cellulolytic enzyme production. However, many fungi were reported to produce cellulolytic enzymes of high potency and stability; of these, Trichoderma species were found to be the most important [38]. Enzymatic hydrolysis of cellulose to glucose is simple, efficient, and cost-effective, reducing pollution and creating new food sources [39, 40].
The main polymer in plant cell walls, cellulose, is stable and recalcitrant [41]. The macromolecular structure of cellulose is more complicated than expected for a linear glucan chain polymer. Cellulose I is composed of β-(1,4)-d-glucan polymer fibrils. Supramolecular properties like lateral diameters vary by species in cellulose I fibrils. A fibril’s polymers’ order depends on their location, with surface polymers being the least restricted. Depending on the starting material and isolation method, cellulose I fibrils can form supramolecular structures, fibril aggregates, and the cellulose network, such as wood pulp fiber walls [42]. This complete porous network is space-oriented complexly. The surface area exposed to enzyme assault may limit material characteristics during hydrolysis [43]. Besides the complicated spatial orientation, cellulose structures have hydrophilic and hydrophobic faces, which exocellulase (CBHI) seems to prefer in cellulose hydrolysis [44].
The present study confirmed that T. viride needs some organic compounds. like nitrogenous compounds such as amino acids and others for their growth. Glutamic acid, methionine, and leucine were all influential in enhancing mycelial growth, but methionine is highly significant, while leucine and Glutamic acid gave a similar effect. The amino acid leucine gave the best mycelial growth at 50 % concentration. The effect of pH level on the fungus T. viride indicated that the maximum growth was at pH 5.0. However, growth decreased dramatically with increasing pH values.
The fungus T. viride had the ability to produce both cellulolytic and pectolytic enzyme production in some different substrates. Carboxymethylcellulose (CMC) was the best substrate for the cellulolytic enzyme. Furthermore, Pectin was the best for pectolytic enzyme production. For any industrial process or as a biocontrol agent, the growth of the fungus needs the addition of carbon and a nitrogen source to the medium. The best amino acids for culturing fungus are T. viride, leucine, and methionine, at a 50 mg/ ml concentration. Carboxymethylcellulose (CMC) was the best substrate for the cellulolytic enzyme. Pectin was the best for pectolytic enzyme production. The best incubation time for cellulolytic enzyme and pectolytic enzyme production was after 16 days. The present study confirmed that the fungus T. viride could produce cellulolytic and pectolytic enzymes in different substrates. Carboxymethyl cellulose (CMC) was the best substrate for cellulolytic enzymes. In addition, Pectin was the best for pectolytic enzyme production. Moreover, the best incubation time for cellulolytic enzyme and pectolytic enzyme production was after 16 days.
Acknowledgments
The authors would like to thank the Deanship of Scientific Research of The University of Hail for funding this research (BA-2208).
Conflict of Interest
None.
Conceptualization: A.S., W.A and S. H; methodology, A.S., S.H and N.A.; formal analysis, A.S., W.A and S. H; writing-original draft preparation, A.S and S.H.; writing–review and editing, A.S., S.H.; project administration, W.A. All authors have read and agreed to the published version of the final manuscript).![]()
References
- Tyśkiewicz R, Nowak A, Ozimek E, & Jaroszuk-Ściseł J. Trichoderma: The current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. International Journal of Molecular Sciences, (2022); 23(4): 2329.
- Takahashi J, Barbosa BV, Martins BD, Guirlanda C, & Moura M. Use of the versatility of fungal metabolism to meet modern demands for healthy aging, functional foods, and sustainability. Journal of Fungi, (2020); 6(4): 223.
- Nelson E, Occurrence of Trichoderma in a Douglas-fir soil. Mycologia, (1982); 74(2): 280-284.
- Widden P, & Abitbol J. Seasonality of Trichoderma species in a spruce-forest soil. Mycologia, (1982); 72(4): 775-784.
- Thrane C, Tronsmo A, & Jensen DF. Endo-1, 3-β-glucanase and cellulase from Trichoderma harzianum: purification and partial characterization, induction of and biological activity against plant pathogenic Pythium spp. European Journal of Plant Pathology, (1997); 103: 331-344.
- Onions AH, Allsopp D, & Eggis HO. Smith's introduction to industrial mycology. Seven edition Edward Arnold Publisher Ltd., U.K. P.P., (1981): 221-373.
- Kunamneni A, Plou FJ, Alcalde M, & Ballesteros A. Trichoderma enzymes for food industries. In Biotechnology and biology of Trichoderma. (2014); 339-344. Elsevier.
- Wang Y, Zeng L, Wu J, Jiang H, & Mei L. Diversity and effects of competitive Trichoderma species in Ganoderma lucidum–cultivated soils. Frontiers in Microbiology, (2022); 13:1067822.
- Sperandio GB, & Ferreira FE. Fungal co-cultures in the lignocellulosic biorefinery context: a review. International Biodeterioration & Biodegradation, (2019); 142: 109-123.
- Li P, Wang H, Liu G, Li X, & Yao J. The effect of carbon source succession on laccase activity in the co-culture process of Ganoderma lucidum and a yeast. Enzyme and microbial technology, (2011); 48(1): 1-6.
- Sanitá LM, & Coutinho de Lucas R. Co-cultivation, co-culture, mixed culture, and microbial consortium of fungi: an understudied strategy for biomass conversion. Frontiers in Microbiology, (2022); 12: 837685.
- Shu G, Yang H, Chen H, & Yang Z. Research on extraction and characterization of cellulase from commercial enzyme preparation. Advance Journal of Food Science and Technology, (2013); 5(7): 839-842.
- Peláez RD, Wischral D, Mendes TD, Pacheco TF, Urben AF, Helm CV, & de Siqueira FG. Co-culturing of micro-and macro-fungi for producing highly active enzyme cocktail for producing biofuels. Bioresource Technology Reports, (2021); 16: 100833.
- Wong DW. Pectic enzymes. In: Food enzymes- structure and mechanism, (1995): 212-236. Boston, MA: Springer US.
- Mandels M, & Wilke CR. Cellulose as a chemical and energy resource. In Biotechnol. Bioeng. Symp (1975), Vol. 5: 81.
- Ljungdahl LG, & Eriksson XE. In: Advances in Microbial Ecology. Vol. 5, ed. K .C. Marshal , Plenum Press. New York, (1985): 237.
- Vasquez GS, Lead MC and Herrea EA. Analysis of the beta 1,2-Glyconolytic system of the biological agent Trichoderma harzianum. Applied And Environmental Microbiology, (1998); 4:1442-1446.
- Abbas HK, Mirocha CJ, & Gunther R. Production of zearalenone, nivalenol, moniliformin, and wortmannin from toxigenic cultures of Fusarium obtained from pasture soil samples collected in New Zealand. Mycotoxin Research, (1991); 7: 53-60.
- Jian D, Yuanyuan L, Hongman Z, Hongbo Z & He H. Factors to decrease the cellulose conversion of enzymatic hydrolysis of lignocellulose at high solid concentrations. Cellulose, (2014); 21: 2409–2417.
- Förster H. Pectinesterases from Phytophthora infestans. In: Methods in enzymology (1980); 161: 355-361. Academic Press.
- Aro N, Pakula T, & Penttilä M. Transcriptional regulation of plant cell wall degradation by filamentous fungi. FEMS microbiology reviews, (2005); 29(4): 719-739.
- Zheng W, Lehmann A, Ryo M, Vályi KK, & Rillig MC. Growth rate trades off with enzymatic investment in soil filamentous fungi. Scientific Reports, (2020);10(1), 11013.
- Abubakar A, Suberu HA, Bello IM, Abdulkadir R, Daudu OA., & Lateef AA. Effect of pH on mycelial growth and sporulation of Aspergillus parasiticus, Journal of Plant Sciences, (2013); 1(4): 64-67.
- Chinedu SN, Eni AO, Adeniyi AI, & Ayangbemi JA. Assessment of growth and cellulase production of wild-type microfungi isolated from Ota, Nigeria. Asian Journal of Plant Sciences, (2010); 9(3): 118.
- Omojasola PF, & Jilani OP. Cellulase production by Trichoderma longi, Aspergillus niger and Saccharomyces cerevisae cultured on waste materials from orange. Pakistan Journal of Biological Sciences, (2008); 11(20): 2382-2388.
- Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical chemistry, (1959); 31(3): 426-428.
- Nayyar BG, Woodward S, Mur LAJ, Akram A, Arshad M, Saqlan Naqvi SM & Akhund S. The Incidence of Alternaria Species Associated with Infected Sesamum indicum L. Seeds from Fields of the Punjab, Pakistan. Plant Pathology Journal. (2017); 33(6): 543-553.
- Gomez-Sagasti MT, Epelde L, Anza M, Urra, J, Alkorta I, & Garbisu C. The impact of nanoscale zero-valent iron particles on soil microbial communities is soil dependent. Journal of hazardous materials, (2019); 364: 591-599.
- Cerecetto V, Smalla K, Nesme J, Garaycochea S, Fresia, P, Sørensen SJ., … & Leoni C. Reduced tillage, cover crops and organic amendments affect soil microbiota and improve soil health in Uruguayan vegetable farming systems. FEMS Microbiology Ecology, (2021); 97(3), fiab023.
- Delgado-Baquerizo M, Maestre FT, Reich PB, Jeffries TC, Gaitan JJ, Encinar D, … & Singh BK. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nature communications, (2016); 7(1): 10541..
- Esperschütz J, Gattinger A, Mäder P, Schloter M, & Fließbach A Response of soil microbial biomass and community structures to conventional and organic farming systems under identical crop rotations. FEMS Microbiology Ecology, (2007): 61(1): 26-37.
- Qiao C, Penton CR, Xiong W, Liu C, Wang R, Liu Z., … & Shen Q. Reshaping the rhizosphere microbiome by bio-organic amendment to enhance crop yield in a maize-cabbage rotation system. Applied Soil Ecology, (2019); 142, 136-146.
- Ling L, Fu Y, Jeewani PH, Tang C, Pan S, Reid BJ, … & Xu J. Organic matter chemistry and bacterial community structure regulate decomposition processes in post-fire forest soils. Soil Biology and Biochemistry, (2021);160: 108311.
- Domsch KH, Gams W, & Anderson TH. (1980). Compendium of soil fungi. Volume 1. Academic Press (London) Ltd.
- Smith KP, & Goodman RM. Host variation for interactions with beneficial plant-associated microbes. Annual review of phytopathology, (1999); 37(1): 473-491.
- Seyis I, & Aksoz N. Production of Lactase by Trichoderma sp., Food Technol. Biotechnol, (2004); 42 (2): 121–124
- Mandels M, & Weber J. The production of cellulases.In Cellulases and their applications. Advan. Chem. Ser., (1969); 95:391- 414.
- Harman GE. Overview of Mechanisms and Uses of Trichoderma spp. Phytopathology, (2006); 96(2): 190-194.
- Faheem VK. Razdan FA, Mohiddin KA, Bhat A & Saba B. Potential of Trichoderman Species As Biocontrol Agents of Soil Borne Fungal Propagules. Journal of Phytology. (2010); 2(10): 38–41
- Cheetham PSJ. The applications of enzymein industry. Handbook of enzyme biotechnology. (1985): 274-379.
- Wolfenden R, & Snider MJ. The depth of chemical time and the power of enzymes as catalysts. Accounts of chemical research, (2001); 34(12): 938-945.
- Hult EL, Iversen T, & Sugiyama J. Characterization of the supermolecular structure of cellulose in wood pulp fibres. Cellulose, (2003); 10 : 103-110.
- Chunilall V, Bush T, Larsson, Per Tomas I, & Kindness A. "A CP/MAS 13C-NMR study of cellulose fibril aggregation in eucalyptus dissolving pulps during drying & the correlation between aggregate dimensions and chemical reactivity" , (2010), 64 (6): 693-698.
- Liu J, Lu J, & Cui Z. Enzymatic hydrolysis of cellulose in a membrane bioreactor: assessment of operating conditions. Bioprocess and biosystems engineering, (2011); 34: 525-532.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


