

Full Length Research Article
Effect of quinoa biomass and biocontrol fungi on expression of IPER gene in mung bean in Macrophomina phaseolina contaminated soil
Farman Ahmad Chaudhury1, Iqra Haider Khan2, Arshad Javaid*2
Adv. life sci., vol. 10, no. 2, pp. 193-199, June 2023
*– Corresponding Author: Arshad Javaid (arshad.iags@pu.edu.pk)
Authors' Affiliations
2. Department of Plant Pathology, Faculty of Agricultural Sciences, University of the Punjab, Quaid-e-Azam Campus, Lahore 54590 – Pakistan
[Date Received: 20/09/2022; Date Revised: 20/12/2022; Date Published Online: 30/06/2023; Date Updated:11/09/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Mung bean is a pulse crop of economic significance, grown in Pakistan for its edible seeds. Its production is severely affected by Macrophomina phaseolina, a necrotrophic pathogen. This study was carried out to investigate the effect of different biocontrol fungi on expression of IPER gene in mung bean plants inoculated with M. phaseolina.
Methods: Pot trials were carried out by adding different concentration of dry biomass of quinoa (DBQ) and six antagonistic fungi viz. Aspergillus flavipes, Aspergillus versicolor, Penicillium digitatum, Penicillium italicum, Trichoderma pseudokoningii and Trichoderma viride in M. phaseolina infected soils. After four weeks of germination of mung bean seeds, RNA was extracted from roots and leaves by using TRIzol method and cDNA was prepared by using SuperScriptTM IV First-Strand Synthesis Kit. IPER gene expression was studied on qRT-PCR and ACT was used as a housekeeping gene.
Results: The expression of IPER gene was higher in positive control (only inoculated with M. phaseolina) than in negative control (no amendment). Moreover, 1 and 2% doses of DBQ showed slightly higher Ct values than in 3% dose where it was dropped down indicating the allelopathic stress of DBQ treated soils. The treatments either with sole application of antagonistic fungi or together with DBQ showed much higher Ct values indicating no stress at all.
Conclusion: Plants only inoculated with M. phaseolina showed the highest disease incidence as compared to the negative control. The soils amended with DBQ and antagonistic fungi significantly alleviated the effect of M. phaseolina. The treatments with increased pathogen stress showed reduced Ct values and vice versa.
Keywords: Biocontrol; IPER gene; Gene expression; Macrophomina phaseolina; Mung bean
Introduction![]()
Mung bean is an economically significant pulse crop in Asian agricultural system including Pakistan [1]. It is a warm season leguminous crop that is cultivated for its edible seeds and sprouts in the form of vegetable salad. It is not only a source of protein but also contains iron and folate in higher amounts than in most of other legumes [2]. Although a considerable improvement in its yield has been made yet its production is low in Asian farming systems due to several fungal diseases [3]. Among various pathogenic fungi, M. phaseolina has an important position as a pathogen as it inflicts damage to the mung bean plant at different growth stages [4]. It is globally known as a devastating pathogen that infects more than 500 plant species. It is mainly a soil- and seed-borne pathogen whose growth is favored under low moisture condition coupled with high temperature (30-35°C) [5]. It generally causes charcoal rot disease in various plant species including mung bean [6]. At the start, the fungal hyphae invade cortical tissues of mung beans followed by the development of spindle shaped lesions with minute microsclerotia and pycnidia [7]. The production of sclerotia in infected areas exhibits charcoal rot disease symptoms. Under unfavorable conditions, microsclerotia are responsible for survival of the fungus for prolonged time period in soil and plant debris making its management a challenging task [8].
To date, different disease management approaches such as physical, cultural, regulatory and use of chemicals have been implemented to eradicate M. phaseolina populations in the soil but each has some limitations [9]. Moreover, the conventional use of fungicides has drastic effects as it disrupts the balance of beneficial microbes in the soil and these are economically not feasible to low-income farmers [10]. Apart from health hazards, the regular use of fungicides may give rise to the appearance of resistant pathogenic strains [11]. To reduce the dependence on chemicals, application of biological agents such as Aspergillus spp. [12], Penicillium spp. [13] and Trichoderma spp. [14] offers good alternatives to the growers for control of plant diseases. They have some interesting properties for instance, they are cost effective, do not pollute the environment and are target specific [15]. The mechanism through which antagonists retard the pathogen growth is not always clear. However, they might show a direct parasitism, release of toxins by means of antibiosis, and competition for available resources [16,17].
In recent past, the use of natural plant products has also gained importance. These products have broad host range, safe, easily biodegradable, cheap, non-phytotoxic, environment friendly and exhibit a structural diversity. Several plant families such as Asteraceae [18], Meliaceae [19], Chenopodiaceae [20], Magnoliaceae, Acanthaceae [21] and Amaranthaceae [22] are known for their antifungal properties. Recent studies have demonstrated that purified compounds isolated from plant extracts [23,24], and plant dry biomass can efficiently be used for the control of plant diseases [25,26]. C. quinoa is native to Andean regions since 3000 BC. Recently, it has been introduced in Africa, North America, Europe, Asia and South East Asia due to its tolerance to frost, drought and salinity [27]. Moreover, it contains saponins, triterpenoids, glycosides, phenolic acids, kaempferol, quercetin, tannins and flavonoids that contribute to diverse biological properties [28]. Hence, considering the adverse impact of fungicides the use of beneficial antagonistic fungi and natural plant products may play a key role in eco-friendly sustainable agriculture system.
In response to pathogenic fungal attacks, plants defend themselves with an arsenal of defensive mechanisms. These involve the formation of structural barriers by the release of antimicrobial proteins, which prevent the pathogen colonization into the host plants [29]. Among the proteins induced during plant defense, peroxidase precursor (IPER) belongs to class Ⅲ is well known [30]. It plays a critical role during the pathogen colonization either by development of stiff plant structures or by creating a more oxygenated environment for the pathogens [31]. It is generally involved in many physiological processes such as wound healing, degradation pathways, cell wall metabolism, removal of H2O2, auxin catabolism, host defense mechanism, oxidation of toxic reductants, and cell growth [32]. In addition, IPER can generate highly reactive oxygen species (ROS) that can possess an intrinsic activity during diverse hypersensitive responses. It also acts as a part of signal transduction pathway during biotic and abiotic stress responses, cell division and programmed cell death [33]. In pathogenic attacks, peroxidase gene expression is triggered to repair the damaged tissues and to provide a protective mechanism [34]. Therefore, the present study was undertaken to examine the effect of biocontrol agents namely Trichoderma spp., Aspergillus spp. and Penicillium spp. and dry biomass of quinoa plants on expression of IPER gene in mung bean plants in M. phaseolina contaminated soil.
Methods![]()
Pot trials
A pot study was carried out during March 2017. For this, sandy loam soil was fumigated and filled in 30 cm diameter earthen pots followed by the application of pearl seed-based M. phaseolina inoculum (10 g kg-1 of soil) prepared on pear millet seeds[12], except in pots of negative control where only same amount of boiled pearl millet seeds were added. After one week, inoculum of the antagonistic fungi (10 g kg-1 of soil) viz. A. flavipes, A. versicolor, P. digitatum, P. italicum, T. pseudokoningii and T. viride were added alone and in combination with C. quinoa dry plant biomasses at different concentrations of 1%, 2% and 3% (w/w). The experimental pots were left for one week and watered when required for the establishment of antagonists as well as the release of allelochemicals by C. quinoa added biomass. Ten seeds of mung bean were sown in all the pots, having three replicates of each. After four weeks of germination, sampling was done from each pot. Harvested root and leaf samples were immediately crushed in liquid nitrogen and stored at −80 °C for further analysis.
Isolation of total RNA and synthesis of cDNA
Total RNA from mung bean roots and leaf samples was extracted by using TRIzol method. For this, 2 mL of TRIzol was added into each falcon (15 mL) containing homogenized plant samples and left for 20 min. After that, 3 mL of chloroform: isoamyl alcohol (24:1) were added and the mixture was centrifuged at 13000 rpm for 20 min. Supernatant was taken in a new falcon, added the same mixture and centrifuged for another 20 min. Again, supernatant was separated followed by the addition of isopropanol and 3 M sodium acetate, and kept overnight at −20 °C. After that, RNA pellet was obtained by centrifuge at 13000 rpm for 10 min, washed with 70% ethanol and mixed in double distilled water. The quality and quantity of RNA were measured through Nanodrop spectrophotometer and processed for cDNA by using SuperScriptTM IV First-Strand Synthesis Kit following the manufacturer protocol.
Gene confirmation with specific primers
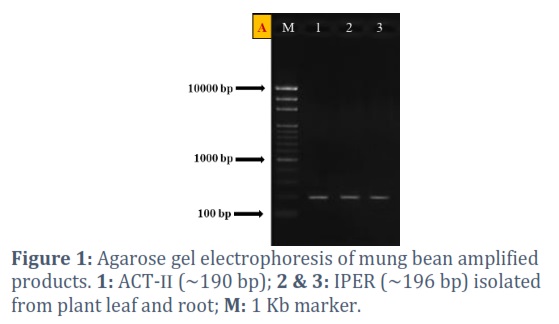
The ORFs of mung bean were amplified by using a set of primers (Table 1) already used for the amplification of ACT (a house keeping gene) and IPER (peroxidase precursor gene). The obtained PCR products were then run on 1% agarose gel for genes confirmation (Fig. 1).
Quantitative real-time PCR assay
The qRT-PCR experiment was done on the StepOnePlusTM RealTime PCR System (Thermo Fisher Scientific) in a 48 well microtiter plates using SYBR® Green PCR Master Mix. A total of 25 µL reaction mixture was prepared by adding 1 µL cDNA template, 1 µL of each forward and reverse primers (10 µM) and 12.5 µL of master mix followed by the addition of 9.5 µL nuclease free water. Three independent biological and technical replicates of each treatment were used to carry out the gene expression (Fig. 2 and 3).
Statistical analysis
All the data were analyzed by ANOVA followed by application of LSD test at P = 0.05.
Results![]()
Disease incidence (DI)
The highest incidence of disease (39%) was recorded in the positive control while no disease was noted in the negative control. In other treatments, where the soil was treated with DBQ and the six species of biocontrol fungi, a significantly (P≤0.05) decrease in DI was recorded. In these treatments, DI ranged from 4 to 8%.
IPER gene expression in response to M. phaseolina
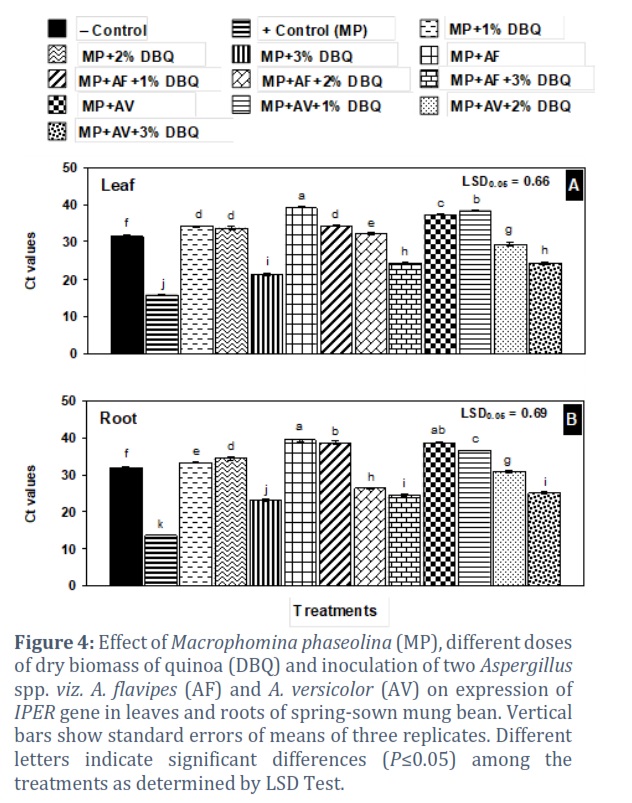
The Ct values were 31.54 and 31.84 in leaves and roots of negative control, respectively. M. phaseolina application in positive control a lower Ct value indicates a higher gene expressionto 15.65 and 13.71 in leaves and roots, respectively, showing the significant increase in IPER gene expression in the positive control as compared to negative control treatment (Fig. 4 A & B).
IPER gene expression in response to quinoa biomass
In leaf, Ct values in 1% and 2% DBQ were a little higher than those in the negative control. Conversely, the Ct value was slump to 21.29 in 3% DBQ showing allelopathic effect of the added plant biomass that resulted in 48% higher gene expression than in the negative control. In root, the effect of DBQ on Ct values was generally similar to that recorded in leaf (Fig. 4 A & B).
IPER gene expression in response to Aspergillus spp.
In leaves of mung bean, generally the Ct values in A. flavipes (AF) or A. versicolor (AV) inoculated treatments, separately or combined with 1% or 2% DBQ, were a little higher or near to that in the negative control (31.54). By contrast, AV + 3% DBQ and AF + 3% DBQ treatments exhibited Ct values of 24.24 and 24.38, showing light stress on the mung bean plants due to allelopathic effects of added plant material causing a slight expression of IPER gene in these treatments over the negative control (Fig. 4 A). A similar trend in Ct values in roots was recorded due to application of Aspergillus species, and DBQ (4 B).
IPER gene expression in response to Penicillium spp.
In treatments where the two Penicillium species were applied singly or in combination with 1% DBQ, the Ct values were much higher than in negative control and were in the range of 35.71–39.19 in leaf and 35.31–39.41 in root (Fig. 5 A and B).
IPER gene expression in response to Trichoderma spp.
Treatments with either of the two Trichoderma spp. alone or together with 1% DBQ showed significantly higher Ct values than the negative control showing no gene expression. The IPER gene was expressed in those treatments where 3% DBQ was added alone or together with T. pseudokoningii (TP) and T. viride (TV) with Ct values in the range of 20.97–23.45 in leaf and 21.62–23.28 in root (Fig. 6 A and B).
Figures & Tables
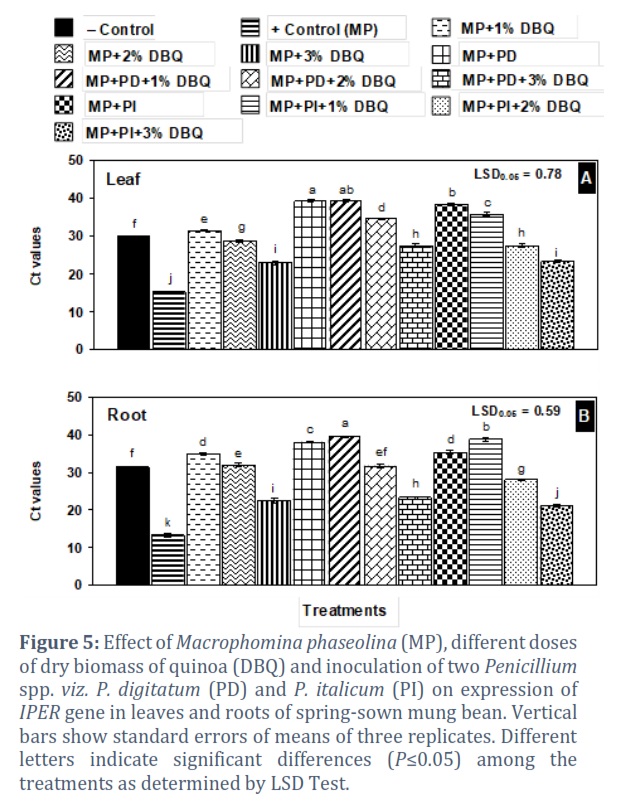
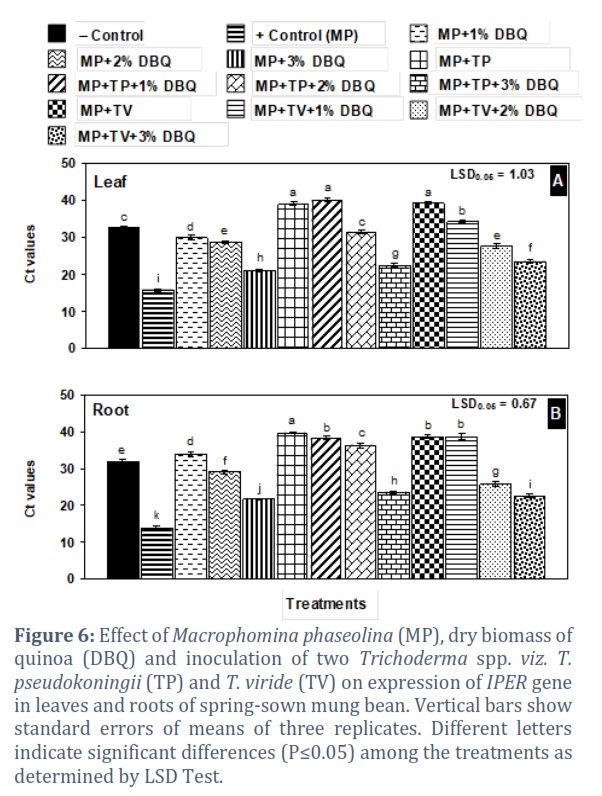

qRT-PCR analysis was used to reveal the different patterns of IPER gene expression and its role in resistance mechanism after M. phaseolina infection. The soil amendments with antagonistic fungi and quinoa dry plant biomass generally increased the resistance in mung bean plants against the charcoal rot pathogen in comparison to positive control. Moreover, different concentrations of quinoa resulted in the distinctive regulation of peroxidase precursor gene in mung bean plants. The present findings showed that the expression of IPER gene was interestingly modulated according to the treatments and increased strongly in positive control root and leaf samples. In 3% amendment of quinoa, it was found to be overexpressed, which was might be due to the release of excessive allelochemicals from the decomposing material. The increased expression of IPER gene after the infection has also been confirmed by previous studies. Bela et al. [35] reported that the attack of fungal pathogens triggered the expression of peroxidase gene and it was proved in Arabidopsis thaliana. Peroxidase precursors perform different biochemical activities and functions under pathogenic stresses by modulating the reactive oxygen species [36]. Peroxidase precursors play a critical role in apple plants defense mechanism infected with Valsa mali [37]. It is a defense related enzyme and an exponentially higher expression was also observed in tomato plants infected with Meloidogyne javanica in comparison to Trichoderma harzianum treated plants [38]. Singh et al. [39] also revealed a positive correlation between IPER activity and resistance to Brassica napus to Verticillium longisporum attack. Previously, it has been reported that IPER is induced exponentially in response to fungal and bacterial infections [30]. Sharma et al. [40] found that the expression pattern of IPER is correlated with phenols oxidation that is dependent on H2O2 availability and is increased exceptionally under tissue damage. To regulate the gene expression, IPER is acquired to perform activities in order to facilitate the opposing plant reactions that are initiated in response to pathogens. Gayoso et al. [41] also described a comparable increase in IPER activity that was parallel to H2O2 production in diseased tomatoes. This might be the result of functions performed by a number of enzymes in susceptible plants. Furthermore, Lanubile et al. [42] reported a rapid induction of IPER gene in maize plants infected with Fusarium verticillioides. In response to Botrytis cinerea, an enhanced expression of IPER has been observed in transgenic tomatoes [43]. Jogaiah et al. [44] also found that the expression of IPER gene was significantly higher in tomato plants when infected with Ralstonia solanacearum whereas in Trichoderma harzianum and Penicillium chrysogenum treated plants, the expression was found to be very low. This change might be due to the presence of antagonistic fungi which arrested the growth of bacterial wilt pathogen. Recently, a finding concluded that soybean plants inoculated with Fusarium virguliforme exhibited an increased expression of IPER gene but in T. harzianum treated plants, a reduced potential of pathogen infection along with defense related gene was observed [45].
Mung bean plants grown only inoculated with M. phaseolina showed the highest expression of IPER gene. Soil amendments with different concentration of DBQ, Aspergillus spp., Penicillium spp. and Trichoderma spp., singly or combined, had very positive effective in lowering the stress of M. phaseolina. The higher Ct values indicated that the pathogen’s stress was very low in these treatments as compared to positive control treatment.
Conflict of Interest
The authors declare that there is no conflict of interest.
![]()
References
- Rani S, Schreinemachers P, Kuziyev B. Mungbean as a catch crop for dryland systems in Pakistan and Uzbekistan: A situational analysis. Cogent Food & Agriculture, (2018); 4(1): Article 1499241.
- Wu X, Islam ASM, Limpot N, Mackasmiel L, Mierzwa J, Cortés AJ, Blair MW. Genome-wide Snp identification and association mapping for seed mineral concentration in mung bean (Vigna radiata L.). Frontiers in Genetics, (2020); 11: Article 656.
- Nair RM, Pandey AK, War AR, Hanumantharao B, Shwe T, Alam AKM, Schafleitner R. Biotic and abiotic constraints in mungbean production – Progress in genetic improvement. Frontiers in Plant Sciences, (2019); 10: Article 1340.
- Sharma CK, Vishnoi VK, Dubey RC, Maheshwari DK. A twin rhizospheric bacterial consortium induces systemic resistance to a phytopathogen Macrophomina phaseolina in mung bean. Rhizosphere, (2018); 5: 71-75.
- Pandey AK, Yee M, Win MM, Lwin HMM, Adapala G, Rathore A, Nair RM. Identification of new sources of resistance to dry root rot caused by Macrophomina phaseolina isolates from India and Myanmar in a mungbean mini-core collection. Crop Protection, (2021); 143: Article 105569.
- Khan AN, Shair F, Malik K, Hayat Z, Khan MA, Hafeez FY, Hassan MN. Molecular identification and genetic characterization of Macrophomina phaseolina strains causing pathogenicity on sunflower and chickpea. Frontiers in Microbiology, (2017); 8: Article 1309.
- Pandey AK, Burlakoti RR, Rathore A, Nair RM. Morphological and molecular characterization of Macrophomina phaseolina isolated from three legume crops and evaluation of mungbean genotypes for resistance to dry root rot. Crop Protection, (2020); 127: Article 104962.
- Khan MR, Haque Z, Rasool F, Salati K, Khan U, Mohiddin FA, Zuhaib M. Management of root-rot disease complex of mungbean caused by Macrophomina phaseolina and Rhizoctonia solani through soil application of Trichoderma spp. Crop Protection, (2019); 119: 24-29.
- Lodha S, Mawar R. Population dynamics of Macrophomina phaseolina in relation to disease management: A review. Journal of Phytopathology, (2020); 168(9): 1-17.
- Kumari R, Shekhawat KS, Gupta R, Khokhar MK. Integrated management against root-rot of mungbean [Vigna radiata (L.) Wilczek] incited by Macrophomina phaseolina. Journal of Plant Pathology and Microbiology, (2012), 3(5): 136.
- Baibakova EV, Nefedjeva EE, Suska-Malawska M, Wilk M, Sevriukova GA, Zheltobriukhov VF. Modern fungicides: mechanisms of action, fungal resistance and phytotoxic effects. Annual Research and Review in Biology, (2019); 32(3): 1-16.
- Khan IH, Javaid A. Antagonistic activity of Aspergillus versicolor against Macrophomina phaseolina. Brazilian Journal of Microbiology, (2022); 53(3): 1613-1621.
- Khan IH, Javaid A. DNA cleavage of the fungal pathogen and production of antifungal compounds are the possible mechanisms of action of biocontrol agent Penicillium italicum against Macrophomina phaseolina. Mycologia, (2022); 114(1): 24-34.
- Khan IH, Javaid A, Ahmed D. Trichoderma viride controls Macrophomina phaseolina through its DNA disintegration and production of antifungal compounds. International Journal of Agriculture and Biology, (2021); 25(4): 888-894.
- Iqbal U, Mukhtar T. Evaluation of biocontrol potential of seven indigenous Trichoderma species against charcoal rot causing fungus, Macrophomina phaseolina. Gesunde Pflanze, (2020); 72(2): 195-202.
- Ghorbanpour M, Omidvari M, Abbaszadeh-Dahaji P, Omidvar R, Kariman K. Mechanisms underlying the protective effects of beneficial fungi against plant diseases. Biol Control, (2018); 117: 147-157.
- Khan IH, Javaid A. In vitro biocontrol potential of Trichoderma pseudokoningii against Macrophomina phaseolina. International Journal of Agriculture and Biology, (2020); 24(4): 730-736.
- Banaras S, Javaid A, Khan IH. Bioassays guided fractionation of Ageratum conyzoides extract for the identification of natural antifungal compounds against Macrophomina phaseolina. International Journal of Agriculture and Biology, (2021); 25(4): 761-767.
- Khan IH, Javaid A, Al-Taie AH, Ahmed D. Use of neem leaves as soil amendment for the control of collar rot disease of chickpea. Egyptian Journal of Biological Pest Control, (2020); 30: Article 98.
- Ali A, Javaid A, Shoaib A, Khan IH. Effect of soil amendment with Chenopodium album dry biomass and two Trichoderma species on growth of chickpea var. Noor 2009 in Sclerotium rolfsii contaminated soil. Egyptian Journal of Biological Pest Control, (2020); 30: Article 102.
- Jara C, Leyton M, Osorio M, Silva V, Fleming F, Paz M, Mellado M. Antioxidant, phenolic and antifungal profiles of Acanthus mollis (Acanthaceae). Natural Product Research, (2017); 31(19): 2325-2328.
- Yasir M, Sultana B, Amicucci M. Biological activities of phenolic compounds extracted from Amaranthaceae plants and their LC/ESI-MS/MS profiling. Journal of Functional Food, (2016); 26: 645-656.
- Javed S, Mahmood Z, Khan KM, Sarker SD, Javaid A, Khan IH, Shoaib A. Lupeol acetate as a potent antifungal compound against opportunistic human and phytopathogenic mold Macrophomina phaseolina. Scientific Reports, (2021); 11: Article 8417.
- Khan IH, Javaid A. Comparative antifungal potential of stem extracts of four quinoa varieties against Macrophomina phaseolina. International Journal of Agriculture and Biology, (2020); 24(3): 441-446.
- Javaid A, Munir R, Khan IH, Shoaib A. Control of the chickpea blight, Ascochyta rabiei, with the weed plant, Withania somnifera. Egyptian Journal of Biological Pest Control, (2020); 30: Article 114.
- Sharf W, Javaid A, Shoaib A, Khan IH. Induction of resistance in chili against Sclerotium rolfsii by plant-growth-promoting rhizobacteria and Anagallis arvensis. Egyptian Journal of Biological Pest Control, (2021); 31: Article 16.
- Angeli V, Miguel SP, Crispim MD, Waleed KM, Hamar A, Khajehei F, Piatti C. Quinoa (Chenopodium quinoa Willd.): An overview of the potentials of the “golden grain” and socio-economic and environmental aspects of its cultivation and marketization. Foods, (2020); 9: Article 216.
- Tang Y, Li X, Zhang B, Chen PX, Liu R, Tsao R. Characterization of phenolics, betanins and antioxidant activities in seeds of three Chenopodium quinoa Willd. genotypes. Food Chemistry, (2015); 166: 380-388.
- Dang L, Van Damme EJ. Toxic proteins in plants. Phytochemistry, (2015); 117: 51-64.
- Pandey VP, Awasthi M, Singh S, Tiwari S, Dwivedi UN. A comprehensive review on function and application of plant peroxidases. Biochemistry & Analytical Biochemistry, (2017); 6(1): 2161-1009.
- Liu H, Dong S, Li M, Gu F, Yang G, Guo T, Wang J. The class III peroxidase gene OsPrx30, transcriptionally modulated by the AT‐hook protein OsATH1, mediates rice bacterial blight‐induced ROS accumulation. Journal of Integrative Plant Biology, (2021); 63(2): 393-408.
- Passardi F, Cosio C, Penel C, Dunand C. Peroxidases have more functions than a Swiss army knife. Plant Cell Report, (2005); 24(5): 255-265.
- Kaman‐Toth E, Danko T, Gullner G, Bozso Z, Palkovics L, Pogany M. Contribution of cell wall peroxidase‐and NADPH oxidase‐derived reactive oxygen species to Alternaria brassicicola‐induced oxidative burst in Arabidopsis. Molecular Plant Pathology, (2019); 20(4): 485-499.
- Brunings AM, Datnoff LE, Ma JF, Mitani N, Nagamura Y, Rathinasabapathi B, Kirst M. Differential gene expression of rice in response to silicon and rice blast fungus Magnaporthe oryzae. Annal of Applied Biology, (2009); 155(2): 161-170.
- Bela K, Horváth E, Gallé Á, Szabados L, Tari I, Csiszár J. Plant glutathione peroxidases: emerging role of the antioxidant enzymes in plant development and stress responses. Journal of Plant Physiology, (2015); 176: 192-201.
- Shigeto J, Tsutsumi Y. Diverse functions and reactions of class III peroxidases. New Phytologist, (2016); 209(4): 1395-1402.
- Feng H, Xu M, Gao Y, Liang J, Guo F, Guo Y, Huang L. Vm‐milR37 contributes to pathogenicity by regulating glutathione peroxidase gene VmGP in Valsa mali. Molecular Plant Pathology, (2021); 22(2): 243.254.
- Naserinasab F, Sahebani N, Etebarian HR. Biological control of Meloidogyne javanica by Trichoderma harzianum BI and salicylic acid on tomato. African Journal of Food Science, (2011); 5(3): 276-280.
- Singh S, Braus-Stromeyer SA, Timpner C, Valerius O, von Tiedemann A, Karlovsky P, Braus GH. The plant host Brassica napus induces in the pathogen Verticillium longisporum the expression of functional catalase peroxidase which is required for the late phase of disease. Molecular Plant Microbe Interactions, (2012); 25(4): 569-581.
- Sharma P, Jha AB, Dubey RS, Pessarakli M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. Journal of Botany, (2012); 2012: Article 217037.
- Gayoso C, Pomar F, Novo-Uzal E, Merino F, de Ilárduya ÓM. The Ve-mediated resistance response of the tomato to Verticillium dahliae involves H2O2, peroxidase and lignins and drives PAL gene expression. BMC Plant Biology, (2010); 10: 1-19
- Lanubile A, Bernardi J, Marocco A, Logrieco A, Paciolla C. Differential activation of defense genes and enzymes in maize genotypes with contrasting levels of resistance to Fusarium verticillioides. Environmental and Experimental Botany, (2012); 78: 39-46.
- Chen S, Vaghchhipawala Z, Li W, Asard H, Dickman MB. Tomato phospholipid hydroperoxide glutathione peroxidase inhibits cell death induced by Bax and oxidative stresses in yeast and plants. Plant Physiology, (2004); 135(3): 1630-1641.
- Jogaiah S, Abdelrahman M, Tran LSP, Shin-ichi I. Characterization of rhizosphere fungi that mediate resistance in tomato against bacterial wilt disease. Journal of Experimental Botany, (2013); 64(12): 3829-3842.
- Pimentel MF, Arnao E, Warner AJ, Subedi A, Rocha LF, Srour A, Fakhoury AM. Trichoderma isolates inhibit Fusarium virguliforme growth, reduce root rot, and induce defense-related genes on soybean seedlings. Plant Disease, (2020); 104(7): 1949-1959
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0