

Review Article
Appraisal of various approaches to produce biohydrogen and biodiesel from microalgae biomass
Liaqat Zeb*1,4, Muhammad Shafiq1, Munir Ahmad2, Asaf Khan3, Dawood Ahmad4, Arshad Mehmood5
Adv. life sci., vol. 9, no. 1, pp. 01-12, May 2022
*- Corresponding Author: Liaqat Zeb (Email: liaqatzeb135@gmail.com)
Authors' Affiliations
2. School of Environmental Engineering, Dalian University of Technology, No.2 Linggong Road, Dalian 116024, P.R. China
3. School of Life Sciences, Sun Yat-sen University, Guangzhou, China
4. Institute of Biotechnology and Genetic Engineering, The University of Agricultural Peshawar, Peshawar – Pakistan
5. The Second Hospital of Hebei Medical University, Hebei Medical University, Hebei, China
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
The need for fossils fuels, including oil, coal and natural gases, has increased dramatically due to a vast expansion in human population and economic growth. The emission of greenhouse gases from fossil fuels leads to disastrous alterations in the earth's climate. In a few decades, fossil fuels will run out because of their non-renewability. Many researchers are motivated to develop new renewable energy sources to replace fossil fuels. Recent research has identified microalgae as a promising resource for the production of biohydrogen and biodiesel. Biohydrogen production affords an environmentally friendly and sustainable approach to generate clear, clean and reliable energy from renewable resources. Green microalgae use sunlight to convert water molecules into oxygen and molecular hydrogen under special conditions. Microalgae have also been reported as a promising feedstock for biodiesel production, as biodiesel is considered the best alternative to petroleum-derived diesel. In the present review, the production of biohydrogen and biodiesel from microalgae through different methods, including indirect and direct biophotolysis, transesterification and lipid synthesis, enhancement approach, is briefly discussed. This article is critical for exploring ideas for future research that can be applied in the commercialization of biohydrogen and biodiesel from microalgae biomass.
Keywords: Biohydrogen; Biodiesel; Biophotolysis microalgae; Transesterification
Introduction![]()
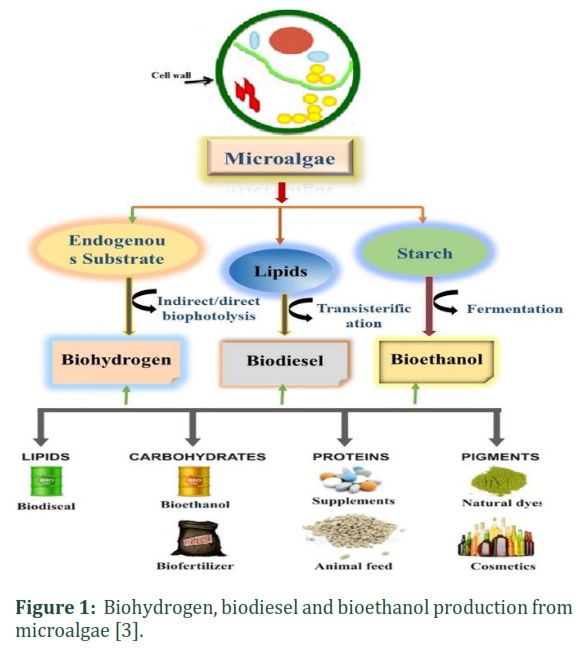
Scarcities in oil, coal and other fossil energy have occurred in the modern era in addition to environmental harm caused by the use of such fossil energy sources. For a country's economic and social development, energy is not only a basic national resource but also a vital material foundation. Both of these problems have completely hindered worldwide fossil fuel development. Development of innovative energy is both a primary method of solving the energy crisis and also an important development strategy for several countries [1]. The accumulation of greenhouse gasses in the environment and the use of fossil fuels as an energy source is unsustainable. The major contributors of carbon dioxide (CO2) release have been identified as the combustion of coal, natural gas, and petrol, which are the main causes of global warming. Rapid industrialization and an increase in population growth compound the problem further, so that fossil fuels are being depleted more rapidly [2]. It is estimated that in almost 40 to 64 years, natural gas and oil will be depleted. The world will need almost 60% more energy in 2030 than what is needed today [2]. During the last century, an increased release of greenhouse gasses and waning water quality were caused by human activities. Each year, approximately 590-880 million tons of methane are released by microbial activities into the atmosphere, and 90% come from biogenic sources [2]. To solve these environmental problems, new renewable technologies need to be developed to generate energy that will reduce CO2 emissions to almost zero in 2100 [1]. Microalgae are mostly photosynthetic eukaryotic organisms that can be altered to live in different environments because of their simple multicellular or unicellular structure. Photoautotrophically, microalgae use CO2 for growth and produce almost half of the atmospheric oxygen. The four divisions of microalgae are included in terms of their abundance: Bacillariophyta (diatoms), Chlorophyta (green algae), Cyanophyta (blue-green algae) and Chrysophyta (golden algae). Microalgae can also be cultured heterotrophically and further subdivided into chemoheterotrophic and photoheterotrophic [3]. The former can oxidize organic compounds to produce energy, and the latter mostly uses solar light as a source of energy. Autotrophic microalgae need inorganic compounds to grow, such as light, salts (phosphate, nitrate ion) and CO2. This autotrophic group can also be further subdivided into chemoautotrophs and photoautotrophs. Microalgae exist as more than 100,000 species around the world, but no more than 30,000 have been classified and investigated according to their pigments, size, color, and cell wall constituents or metabolism, which are important for human requirements [4]. Biofuel production from microalgae is attractive, as algae produce 10-100 times the amount of oil from waste when compared to other well-known oil and food crops [3]. When compared to land-based plants, the algal growth rate is 100 times faster, and the doubling time can be as low as 3-4 h under promising growth conditions [3]. Due to the simple cell composition and vast ratio of surface to volume in algae, algae can uptake a large quantity of nutrients from water to promote growth rate. Algae can transform CO2 and light during photosynthesis into a biomass rich in mineral components. They can grow in fresh, waste and marine water, which shows their robustness and versatility, and the photosynthetic yields of algae increase with year-round production. An algae biomass rich in minerals can be harvested for various kinds of biofuels, including bioethanol from carbohydrate fermentation, biomethane from anaerobic digestion, biohydrogen production, and biodiesel from microalgae lipid and protein via thermochemical processes or biochemical processes (Fig. 1) [4,5]. The starch-based cytoplasm and high-carbohydrate content of their holocellulose-based cell walls have made the algal biomass a suitable feedstock for bioethanol production. These carbohydrate polymers are broken into simple sugars during microbial hydrolysis, followed by fermentation to yield bioethanol. To produce bioethanol via fermentation, starch-rich microalgae have been extensively studied, and different pretreatment methods have been appraised to release the fermentable sugars from the algal biomass to boost bioethanol production. Significant research efforts have been carried out to use the algal biomass as a feedstock for bioethanol production. Although the overall process can vary, the production of bioethanol from biomass usually contains the following steps: (i) pretreatment of feedstock; hydrolysis; and (iii) fermentation of bioethanol [5].
Methods![]()
Literature survey and selection criteria
The Google scholar, Xue Baidu, and Google Web databases were used to obtain data for this review manuscript. Through various keywords, research articles were retrieved such as biodiesel, biohydrogen, microalgae, and biofuels. The literature achieved for this manuscript were further screen by exclusion and inclusion criteria. More than 40 articles were selected which were published in the last 10 to 15 years..
Biohydrogen Production from Microalgae
Currently, there is an abundant need for hydrogen energy in the world; the exhaustion of hydrogen increased up to 900 billion m3 in 2011. Currently, 96% of all hydrogen is produced from fossil fuels: 18% from coal, 48% from natural gas, and 30% from different hydrocarbon-containing fossils [6]. Hydrogen will be used in the future as a substitute source for fossil fuels, and this could be of key significant interest because the reserve of fossil fuels is being drained [4]. Hydrogen production using microorganisms is an ecologically friendly way of obtaining renewable energy. There are various kinds of microbes utilized for this purpose, including archaea, bacteria, and algae. Investigating in this article, the main focus will be on the use of microalgae for the production of hydrogen. The key significance of the generation of photosynthetic hydrogen is to reduce the emission of greenhouse gases, hence, this strategy is promising and attractive [5].
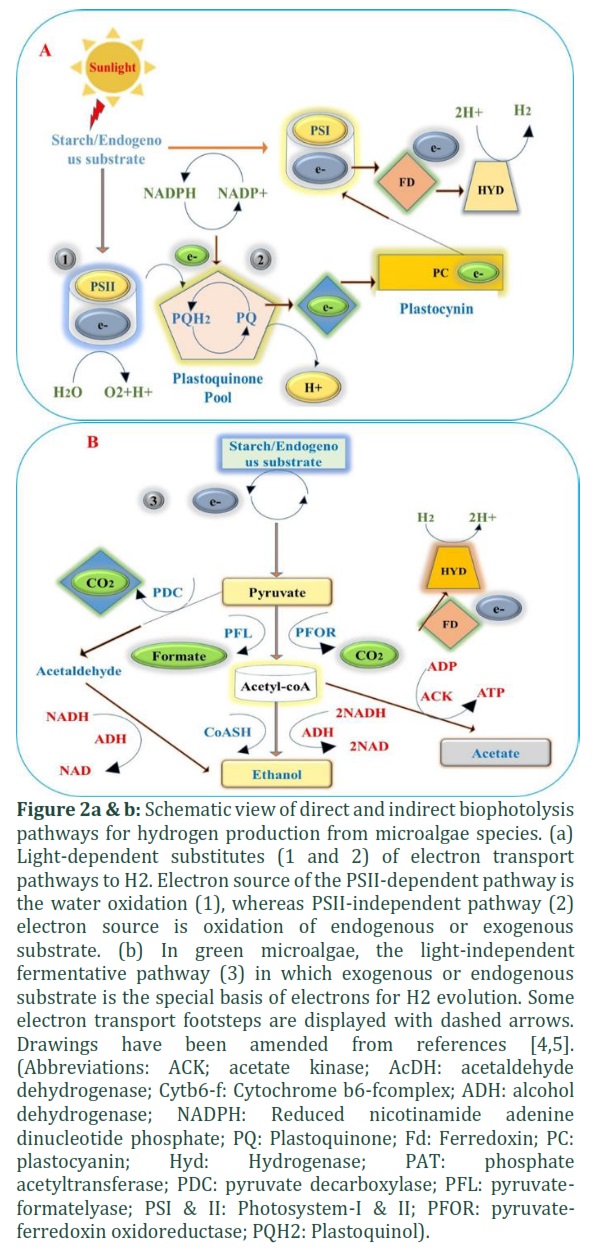
Several kinds of photosynthetic organisms are capable of producing hydrogen. In the late 1990s, green algae Chlamydomonas reinhardtii was found to switch from oxygen production to hydrogen production during photosynthesis in sulfur scarcity medium [6]. Due to the depletion of sulfur the internal oxygen flow was interrupted, resulting in hydrogen production catalyzed by the enzyme hydrogenase [7]. Cyanobacteria and green microalgae have a pigment molecule which are capable to capture sunlight energy and transform it into biochemical energy splitting water to molecular protons (H+) and oxygen (O2). In the thylakoid membranes of chloroplast and cytoplasm of green algae and cyanobacteria occurring a complex redox reaction called photosynthesis. This complex photosynthetic redox reaction transfer electrons and establish two different reactions, namely indirect biophotolysis (dark) and direct biophotolysis (light) reactions. The detailed procedures for biohydrogen generation through both routes are discussed below [5].
Direct biophotolysis route
Hydrogen production under light irradiation is referred to as direct biophotolysis. Photosystem II (PSII) or photosystem I (PSI) absorb light energy in their pigments, P680 and P700, in which the oxidation of water occurs to increase the energy level of electrons [4]. The electrons are shifted from PSII through various electron transport phases (e.g., the plastoquinone (PQ) pool, cytochrome b6-f complex and plastocyanin (PC)) to PSI and then to ferredoxin (Fd), driving hydrogenase to catalyze hydrogen formation from protons (Fig. 2a) [8]. Photosynthetic oxygenic organisms (e.g., algae, cyanobacteria and plants) consume H2O as the resource of together protons and electrons and release oxygen as a by-product [9,10]. In the thylakoid membranes under anaerobic conditions, unicellular green algae can transport their inner photosynthetic electron flow toward H2 generation. While the final reaction of this process is catalyzed by [FeFe]-hydrogenase enzyme. [8]. The enzyme catalyzes two bidirectional reactions according to the following chemical equations:
![]()
In the PSII-independent pathway oxidation of cellular endogenous substrates occurs which generates electrons for hydrogen production. Study the analysis of endogenous substrates, electrons are converted through the plastoquinone pool to PSI with the help of the enzyme [FeFe]-hydrogenase (Fig. 2a) [4].
Three kinds of enzymes are responsible for the reaction of hydrogen formation. [FeFe]-hydrogenase is generally found in algae, [NiFe]-hydrogenase is generally found in cyanobacteria, and [Fe]-hydrogenase is found in archaea. Several genes of different [FeFe]-hydrogenase and related enzymes have been genetically modified from various microalgae sources, comprising Chlorococcum littorale, Chlamydomonas reinhardtii, Chlorella fusca, Scenedesmus obliquus and Platymonas subcordiformis. hydA1 and hydA2, two genes of [FeFe]-hydrogenases from the model green microalgae C. reinhardtii, are only expressed under anaerobic conditions. hydA1 is an important gene for hydrogen generation, while hydA2 plays a secondary role in this process. The accessory genes hydEF and hydG encode the proteins necessary for stimulation of the enzyme [FeFe]-hydrogenase [5].
Various methods have had limited success at overcoming the inhibiting effect of oxygen on [FeFe]-hydrogenase. The growing medium for microalgal biomass culturing has some inert gasses in gas purging, which help to eliminate the amount of O2 from the suspension. The cells’ own mitochondria consume oxygen when relieved over innovative manipulation of the equivalence within respiration and photosynthesis due to the use of sulfur-deprivation metabolic mechanisms. To use alternative tools for microalgae H2-production, approaches include engineering an enzyme [FeFe]-hydrogenase O2-tolerant, changing the tertiary structure of proteins to shrink, opening the channel of [FeFe]- hydrogenase to allow molecular H2 to disperse via the H-cluster [7], or altering the configuration of the active site of [FeFe]-hydrogenase to block oxidative inhibition. Photoheterotrophic and facultative green microalgae, Chlamydomonas reinhardtii and Scenedesmus obliques, are representative of biohydrogen research [4]. The unicellular microalgae Scenedesmus obliquus could either release H2 under anaerobic conditions in the light or dark or during CO2-fixation using H2 as an electron donor. Many other algal species, such as Platymonas subcordiformis and Chlorococcum littorale, have also been reported to produce hydrogen. Some of microalgae species has been reported in Table. 1, which produce biohydrogen through direct biophotolysis way [5].

Indirect biophotolysis
In the case of light-independent fermentation, glycogen and starch content are used as a substrate to produce hydrogen, and this pathway is called indirect biophotolysis [13]. This method proceeds in two stages. During the first step, under light, the synthesis of carbohydrates occurs. During the second step, hydrogen is produced from carbohydrates and different tiny organic molecules, like ethanol, acetate, and format are generated [4,13].
During oxygenic photosynthesis, the accumulated microalgae cellular starch reserves are degenerated biochemically by the cells into pyruvate, and then the enzyme pyruvate-ferredoxin oxidoreductase (PFOR) oxidizes pyruvate to produce acetyl-CoA and CO2. A modification of Fd also occurs, which carries electrons to the enzyme hydrogenase for the catalytic production of H2. It is reported in a species Chlamydomonas reinhardtii that the enzyme pyruvate-formate-lyase (PFL) oxidizes pyruvate to releases acetyl-CoA and formate [10, 14]. Though metabolic ways, such as the alcohol dehydrogenase [14]/acetaldehyde dehydrogenase (AcDH) pathway, exist, ethanol is generated by these pathways [10]. Acetate kinase (ACK) together with phosphate acetyltransferase (PAT) cooperate to produce acetate from acetyl-CoA.
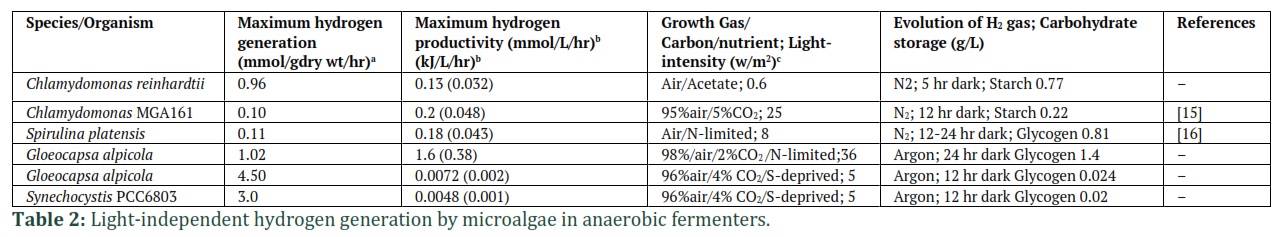
It is investigated that microalgal cells producing less H2 through a light-independent fermentative pathway as compare to light-dependent pathways [17]. The electrons are derived from endogenous substrates, and water oxidation catabolism does not precisely describe their percentages and contribution. In the algal chloroplasts, the key significance of physiological electron transport leading to H2-generation is to enable ATP synthesis. Following prolonged and unfavorable anaerobic circumstances, ATP is required for the survival and function of cells. Under anaerobic requirements, survival depends on the stored starch amount overlaps in the cells with the disruption of starch duration, which generates hydrogen (Table. 2) [17].
It is very important to analyze PSII inhibitor treatment. Such as treatment like 3-3,4-dichlorophenyl-1,1- dimethyl urea (DCMU), which deactivates PSII in sulfur-deprived and H2-generating cells right afterwards carrying cells to a sulfur-free media, results in neither starch accumulation nor H2 generation [4]. In another case, earlier to DCMU treatment, cells were induced to accumulate starch and were capable of generating H2, displaying the importance of both starch accumulation and PSII in the H2 generation procedure (Fig. 2b). Indirect biophotolysis has benefits because of the capability to utilize various carbon sources to afford electrons and protons for hydrogen production which is prominently found in cyanobacteria [18].
Enhancing the yield of biohydrogen production
Hydrogen generation rates depend on various parameters, including approaches to elevate hydrogen production yields and efficiencies. These are the following recent evaluated and major parameters discussed below: (i) metabolic/genetic engineering; and [19] photobioreactor and its geometry. Currently, numerous metabolic engineering methods have been recognized for biological hydrogen generation utilizing microalgae biomasses. Contrast to some wild kind species, a huge substantial outcome on H2 evolution has been perceived ensuing metabolic modification of few microalgal species [20]. Various key concerns for maintainable and necessary H2 generation are the struggle for electrons through assimilatory pathways and the sensitivity of oxygen enzymes. Therefore, the enhancement utilizing microalgal cells for high hydrogen production yield is a challenging concern. On the laboratory scale, there is merely hydrogen production with low product yield for commercial applications. To address low product yield, it’s a key issue to improve hydrogen production for economical purposes by noticing the key scientific background research on microalgae-based biofuels [4].
There are certain possible routes to enhance the hydrogen generation utilizing genetic engineering tools: i) eradication of ways competing for electrons; ii) uptake hydrogenase inactivation; iii) elimination of the sensitivity of oxygen for hydrogen enzymes. Generally, a combination is needed between primary electron donors such as NAD(P)H and decreased ferredoxin with protons to generate hydrogen by the role of Hox-hydrogenase bidirectional and nitrogenase. In a light-dependent race through using targeted ferredoxin-NADP+-oxidoreductase (FNR) variants and ferredoxin, electrons can be converted to the enzyme hydrogenase generating a five-fold boosted hydrogen progression [4]. One of the studied species Synechocystis PCC 6803 by deleting the big subunit NdhB of NADPH-dehydrogenase (NDH-1) and deactivation of the respiratory complex I concluded in a reduced level of O2 below light situation. Besides, another possible pathway is nitrate assimilation pathway which may overcome the flow of electrons to hydrogen metabolism. Genetic engineering pathways by removing electrons in competitive ways can be an encouraging and attractive strategy to enhance hydrogen evolution [5].
Secondly, uptake hydrogenase inactivation for enhanced hydrogen production is a key obstacle. Enhancing the hydrogen evolution in nitrogen-fixing cyanobacteria, the main hurdle is the uptake of hup-hydrogenase. Consequently, a prominent approach to improve the hydrogen generation from different microalgae species required to overcome the hydrogenase uptake. Aforementioned literature studies demonstrated that hydrogen uptake-deficient mutants of Nostoc punctiforme NHM5 (∆hupL), Anabaena variabilis AVM13 (∆hupSL) and Anabaena siamensis TISTR 8012 (∆hupS) , these all species have the capability to generate hydrogen with greater significant rate compared to wild kinds [4].
Another key concern for hydrogen production consuming various systems is the utmost oxygen sensitivity of hydrogen-producing enzymes. The splitting of water in PSII system may prohibit the hydrogenase activities which generate oxygen because the cyanobacteria and green algae are oxygenic photosynthetic [4]. More importantly, rapid deactivation of hydrogenases by oxygen is supposed to be a main aspect for maintained and proficient H2 generation. One of the mutant strain C. reinhardtii D1 has been investigated that imported a double amino acid substitution, isoleucine substituted by leucine residue L159, and tyrosine (L159I-N230Y) by asparagine residue N230 exhibited a greater rate of H2 generation under sulfur deprivation [11]. Besides, suitable hydrogenase enzyme heterologous overexpression into the cells is also of great interest. Furthermore, the heterologous expression in cyanobacteria of O2-tolerant hydrogenases is entirely challenging. Because of a wide number of particular maturation enzymes, the maturation of [NiFe]-hydrogenases is required [6]. In different literature studies reported that nitrogenase enzyme in cyanobacteria frequently produced H2. Currently, it has been identified that light-induced O2 uptake performed protected nitrogenase, because of the bounded heterocyst-specific flavodilol protein (Flv3B). Such changes maintain microoxic conditions inside of the heterocyst Anabaena sp. PCC 7120 of filamentous cyanobacterium [21].
For large scale, economic biohydrogen production needs improvement in the photobioreactor. In common, many large open/raceway ponds are used for microalgae cultivation however many closed photobioreactors have been designed for economic scale offering maximum biohydrogen production. Designing an optimized photobioreactor for large industrial-scale hydrogen production is still a limiting factor. In analysis of the study, various key measures should be focused for a particular photobioreactor designing: i) enlarge the surface area of incident light which improves hydrogen production; ii) with efficiency and convenience the reactor should allow sterilization; iii) the photobioreactor must be designed like enclosed system in which H2 stored without any loss. There is still required extensive research towards the implementation of photobioreactors for large industrial scale low-cost H2 production. Besides, the following parameters must also be considered like: (i) material and size construction of reactor; [19] sources of electrons; (iii) photobioreactor geometry; (iv) metabolic/genetic engineering tools (v) illumination settings; and (vi) mixing and cultivation conditions [4].
Biodiesel production
In recent years, biodiesel fuel has gained significant attention, as it is renewable, non-toxic and biodegradable. It releases less gaseous toxins than normal diesel and contributes no net sulfur or carbon dioxide to the atmosphere. Different four primary methods are reported which make biodiesel, microemulsions, thermal cracking (pyrolysis), direct use and blending and transesterification [22]. The utmost important and common way is transesterification as the biodiesel from transesterification amalgams with diesel fuel in diesel engine or can be used directly. Since 1988, in many European countries biodiesel rapeseed methyl ester has been in commercial use as an alternative fuel. Since 1988, in many European countries biodiesel rapeseed methyl ester has been in commercial use as an alternative fuel. The commercialization and economic aspect of biodiesel production hinders its large-scale use and development, primarily due to its high feed cost. For biodiesel fuel production, microalgae have been advocated as very worthy candidates because of their advantages of faster growth, higher biomass production and higher photosynthetic efficiency compared to other resources [5].
Biodiesel production from microalgae oil
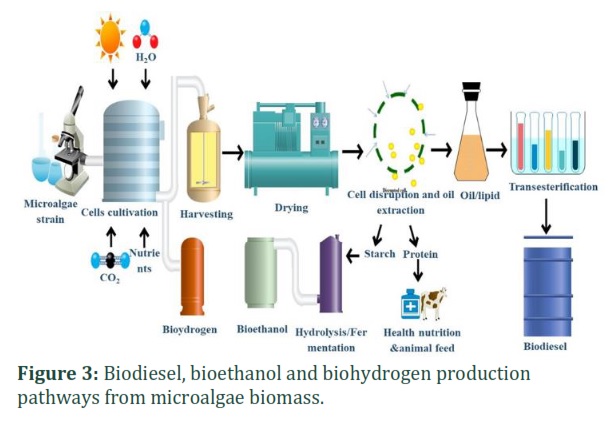
Common crude material, such as biomass enriched with triglycerides, is used for biofuel production (Fig. 3). Biodiesel production is a two-step process: extraction of lipids followed by a triglyceride transesterification reaction. In a transesterification reaction, alcohol and oil utilize different kinds of catalysts to generate glycerine as a by-product and methyl esters. Triglyceride (TAG) converts to diglycerides and then monoglycerides, followed by esters along with glycerol as a by-product. This method overcomes the viscosity of oil to natural, such as the oil used as diesel. Commonly, alcohol and oil are processed in a different (1–9) ratio, which produces the product biodiesel and glycerol in 9–10: 1 ratio. The product, fatty acid methyl esters (FAME), is the main constituent of biodiesel. The given reversible reaction equation shows the biodiesel production process [5].
Chemically, mineral diesel is near to biodiesel, but it has less volumetric heat capability. Also, biodiesel can be a substitute for petroleum diesel fuels. It is probable to remove mineral diesel using biodiesel, and biodiesel viscosity are close to that of mineral diesel. There is various kind of advantages for using biodiesel as an alternative source, because it is easily renewable, cheap to produce, has reduced toxicity, reduced the risk of global warming, and decreased CO2 emission. CO2 is generated in the atmosphere when biodiesel is burned, while crops use this CO2 for growth. Compared to petroleum diesel fuel, biodiesel can minimize the emission of CO2 by up to 78.5%. The most promising source for the production of biodiesel is algae. Various data reported that some species of algae are composed of 60% lipid, which can be easily transformed to biodiesel. Compared to some plants, from one kilogram of algae biomass can be easily produced biodiesel [19]. Photobioreactors or high-rate algal ponds can be used for algal growth to control specific conditions When the algae biomass produced under special conditions, then this biomass is pre-treated, led by a methyl transesterification process of alcohols with lipid fractions. Some catalysts are necessary at this step. After the reaction, the obtained substances are then refined and improved into biodiesel. It is reported that compared to tradition crops, microalgae can produce biodiesel 200 times more efficiently [23].
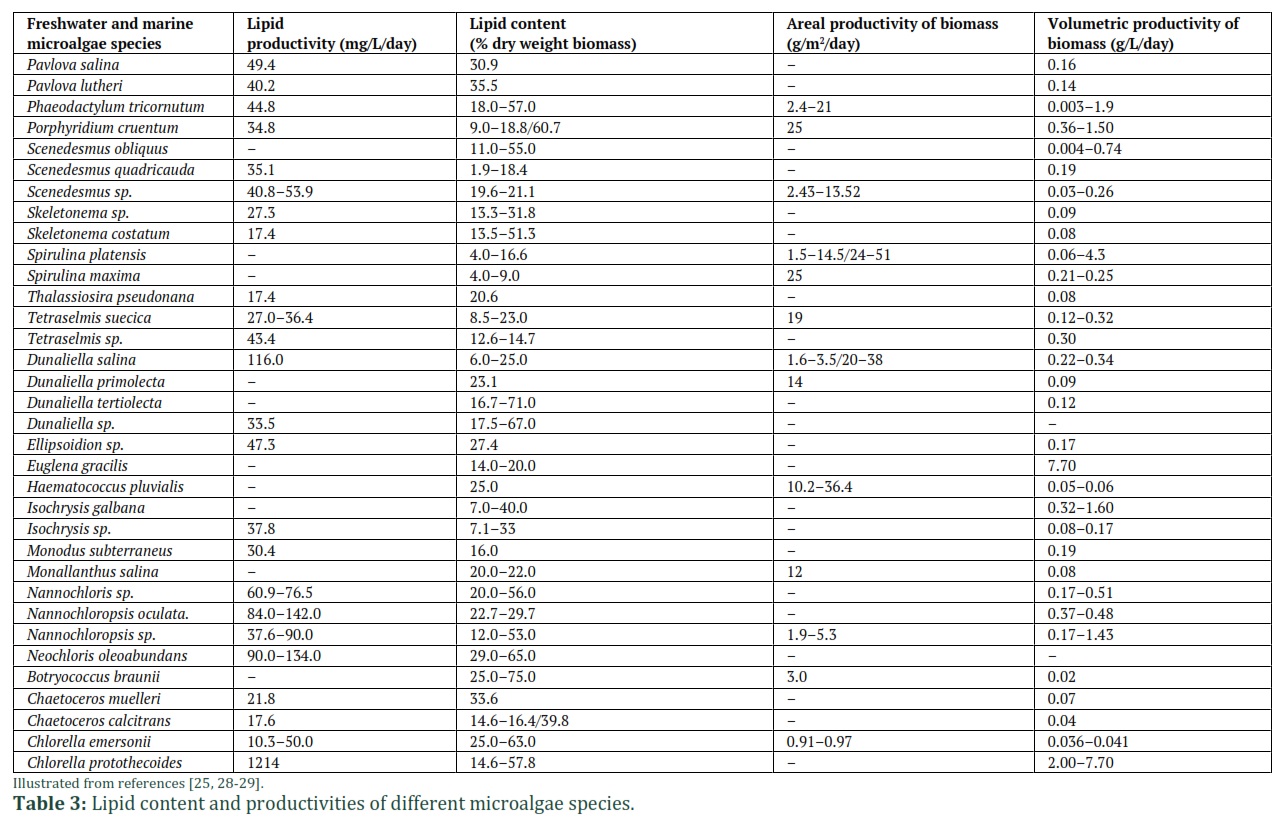
Obtaining a large amount of biofuel from microalgae is faster and easier than in crops, and large amounts of pesticides and herbicides are not necessary. Some important features of microalgae make it a valuable source for biodiesel production. For example, Chlorella minutissima produces higher quantity of lipids at 25 °C in basic medium, and the microalgae C. protothecoides contains 55% lipid when grown under nitrogen limiting conditions [24]. Additionally, it has been reported that B. braunii765 can generate biodiesel under 25 °C. Additionally, it has been revealed that crops oil and petroleum diesel have similar physical and chemical characteristics to biodiesel from microalgae. In microalgal biomass, the total lipid content varies from 1% to 75%, which depends upon cultivation conditions and strains, and the values are mostly bigger than 40% below stress nutrient conditions (Table. 3). The highest total lipid content was reported in microalgae compared to cyanobacteria [25]. Compared to microalgae, macroalgae have insufficient oil content (below 5%w/w), which considered imperfect for generation of biodiesel [12].
Various data reported that some species of algae are composed of 60% lipid, which can be easily transformed to biodiesel. Compared to some plants, from one kilogram of algae biomass can be easily produced biodiesel [19]. Photobioreactors or high-rate algal ponds can be used for algal growth to control specific conditions When the algae biomass produced under special conditions, then this biomass is pre-treated, led by a methyl transesterification process of alcohols with lipid fractions. Some catalysts are necessary at this step. After the reaction, the obtained substances are then refined and improved into biodiesel. It is reported that compared to tradition crops, microalgae can produce biodiesel 200 times more efficiently [23].
Obtaining a large amount of biofuel from microalgae is faster and easier than in crops, and large amounts of pesticides and herbicides are not necessary. Some important features of microalgae make it a valuable source for biodiesel production. For example, Chlorella minutissima produces higher quantity of lipids at 25 °C in basic medium, and the microalgae C. protothecoides contains 55% lipid when grown under nitrogen limiting conditions [24]. Additionally, it has been reported that B. braunii765 can generate biodiesel under 25 °C. Additionally, it has been revealed that crops oil and petroleum diesel have similar physical and chemical characteristics to biodiesel from microalgae.
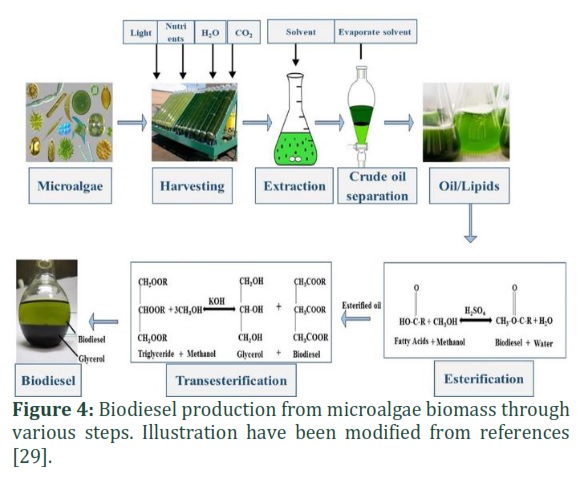
In microalgal biomass, the total lipid content varies from 1% to 75%, which depends upon cultivation conditions and strains, and the values are mostly bigger than 40% below stress nutrient conditions (Table. 3). The highest total lipid content was reported in microalgae compared to cyanobacteria [25]. Compared to microalgae, macroalgae have insufficient oil content (below 5%w/w), which considered imperfect for generation of biodiesel [12]. The single-stage process replaced the conventional various two-step method of methyl esterification because it was expensive. The direct transesterification of wet biomass can significantly minimize two-step of oil extraction and methyl esterification into one step by overcoming downstream processing (Fig. 4) [26]. Due to the presence of free fatty acid, foam is generally in the bottleneck during the transesterification process. Cleaning of the biodiesel is needed to make it valuable while removing foam other raw organic acids. Utilizing the hydrogenation process can improve the short life of biodiesel. Using 100 kg of oil in a transesterification process produced almost 100 kg biodiesel. For example, 100 kg of biodiesel is equivalent to 117.6 L defining a density equal to 850 kg m3 at 277–373 K [27]. Reported in the literature that about 200 kg of microalgae biomass comprises up to 50% (w/w) lipid, which produces 117.6 L of biodiesel [28]. To produce commercially inexpensive biodiesel, the economic cost of algal biomass should be kept up to 0.5 $/kg, based on the oil price in the market. Moreover, the current price of algal biomass requires to reduce by 6–7 fold to secure the practices commercially valuable. It is estimated that the cost of biodiesel derived from microalgae may be comparable to the cost of diesel in 2050. Some of the optimistic views of biodiesel cost reports estimated a range of US$ 0.42–0.97 L1. Solazyme calculated cost of approximately US$17 L1 to generate biodiesel exhausting heterotrophic cultivation methods [30]. Mostly, the cost depends on the growing system, climatic conditions, availability of labor, the algal species, etc. Comparison of various important characteristics of diesel, biodiesel and microalgae biodiesel are elaborated in Table. 4.

Distribution and importance of total lipid in algal cells
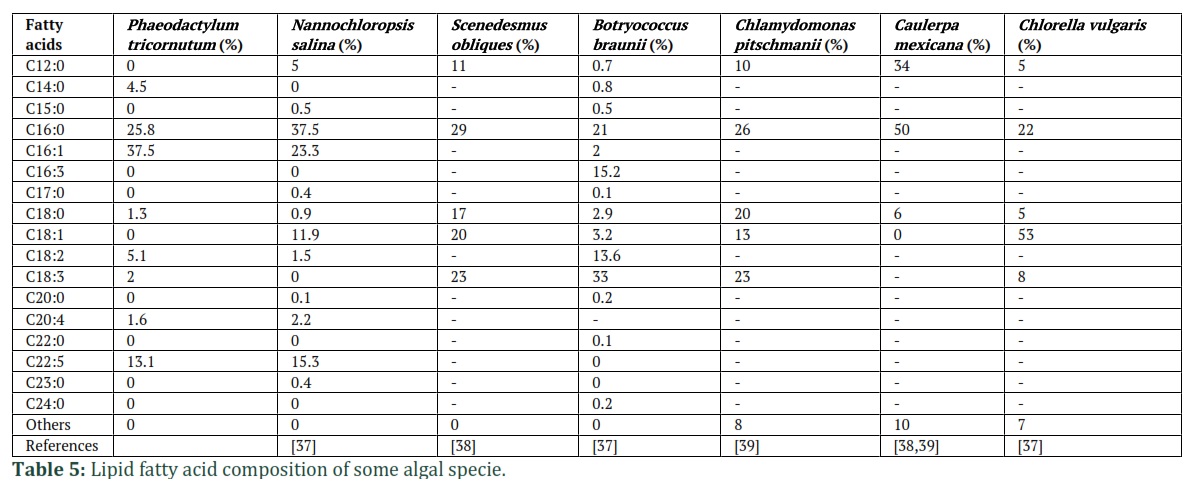
Waxes, sterols, free fatty acids, different glycerides, carotenoids, isoprenoids, etc., are distributed in the cytoplasm of algal cells. Glycolipids and phospholipids are polar lipids that compose the layers of cell membranes. Phosphate moieties, polar hydrophilic sugars combine with saturation degree of fatty acyl chains manages membrane fluidity. Fatty acid and glycerol esters are glycolipids which bind to sugars by one hydroxyl group such as galactose. The most common monounsaturated and saturated fatty acids are found in algal samples with oleic acid (C18:1) or palmitic acid (C16:0) as the major fatty acids (Table. 5). Further, long-chain polyunsaturated fatty acids (VLC-PUFAs) are synthesized in significant amounts, which enables to adjustment of various modifications including pH, UV irradiation, temperature conditions and the environment [5]. Triacylglycerols (TAGs) are profoundly stored in the cytoplasm. TAGs are a highly decreased form carbon comprising of three fatty acids attached to a glycerol backbone. Under normal conditions, glycerol-based membrane lipid in algae largely synthesizes to maintain increased growth. Furthermore, in the cells TAGs are generated when energy depletion of the cells increases by input energy. Under favorable environmental conditions, the stored TAGs can be readily utilized for energy [36]. Cell lipotoxicity can be caused by various generated free fatty acids (FFAs) surviving in the cell's cytoplasm, but TAGs protect the cell from lipotoxicity by serving as a sink for FFAs. Furthermore, during stress conditions, TAGs eradicating excess energy and electrons preserving oxidation and save the cell.
Various factors affecting TAG synthesis
Different parameters producing stress to microalgae, such as nutrient starvation, various temperatures, salinity, pH and culturing time, control fatty acid composition and improve TAG synthesis [14]. For example, the lower temperature has been reported for the enhancement of unsaturated fatty acids production to maintain cellular processes by maintaining membrane fluidity [40]. The yields of polyunsaturated fatty acids (PUFAs) in P. tricornutum are significantly increased when the temperature decreases. The streamflow of stabilized carbons towards the production of sugar and lipid rather of protein was gained in the situation of nitrogen starvation [41]. During nitrogen deprivation, more than a four-fold increase in lipid synthesis and growth of significant amounts of C18:1 was examined in Chlorella vulgaris. But more importantly the amount of DHA, EPA (omega-3 fatty acids), C18:3, C18:2, and C16:2 was decreased. In the formation of PUFAs, small light intensity was determined to maintain PUFAs, while maximum light intensity was found to enhance mono-unsaturated and saturated fatty acid synthesis [42]. Increasing the number of TAGs will enhance the efficiency and yield of biodiesel, although the total amount of oils can be transformed into biodiesel. The quality of biodiesel depends on the existence of different unsaturated and saturated fatty acid in the TAGs. Oxidative stability, quality, cold storage, and ignition quality characteristics are essential parameters of biodiesel. Hydrolytic degradation of raised cetane numbers and water prevents biodiesel from oxidative stability, which ensures quicker ignition and helps in lasting usage. The quality of cold flow enhances due to the unsaturated to saturated fatty acid composition, which is a characteristic of biodiesel that inhibits it from freezing at lower temperatures. To modify the transesterification and purification of FAME, a smaller acyl chain length (C8–C14) was utilized.
Brief mechanisms of lipid synthesis
Utilizing two different pathways can synthesize TAGs in the cell, the acyl CoA dependent (de novo fatty acid biosynthesis) and acyl CoA independent pathway. For de novo synthesis of fatty acid, the chloroplast of algal cells is the synthesis site. This pathway begins with a carboxylation reaction, which is followed by the production of three carbon complexes, such as malonyl CoA. Moreover, with the help of ATP and acetyl-CoA carboxylase (ACCs) enzyme, acetyl-CoA irreversibly merges with CO2 in this stage. It is reported that in different green algae kinds, acetyl CoA is formed in both the chloroplast and cytosol [43]. In the diatom Cyclotella cryptica, ACCs were over-expressed by two- to threefold with no improvement in the lipid content [44]. In addition, malonyl-CoA: acyl carrier protein (ACP) connected with the help of ACP transferase (MAT) which transfer malonyl moiety, producing malonyl-ACP. Malonyl-ACP and acetyl-CoA combine and undergo condensation, reduction, and dehydration reactions. They then undergo a process called reduction reaction with the help of enzymes dehydrase, [18], reductase (KAR), enoyl-ACP reductase (ENR) and 3-ketoacyl-ACP synthase, respectively [43]. Repeating the cycle results in the four steps of progressive elongation of 2 carbons per series, following the formation of the precursor acyl-ACP moiety chain. At the end of each cycle, saturated fatty acid 16-18 carbon chains are formed.
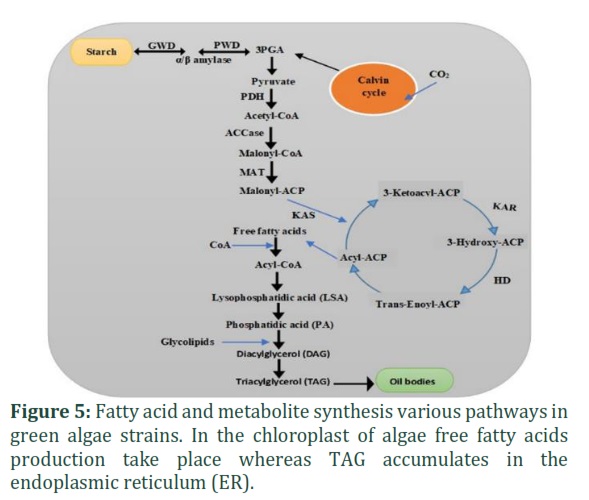
An enzyme stearoyl-ACP desaturase double bonds were proposed in the saturated fatty acid chain. In the Kennedy pathway, direct transfer of the acyl carrier protein (ACP) to the glycerol-3-phosphate (G3P) backbone by acyltransferases. While the elimination of the acyl group from ACP by the action of the acyl-ACP thioesterases (FATs) enzyme is also a possible way, where the elongation process eliminates. The Kennedy pathway, a stepwise reaction adding various hydroxyl groups of the glycerol, effects with G3P. An estimated about 40% improvement was found in the lipid content of algae due to the over-expression of G3P [45]. First, the FFA chain was transformed to position one of the G3P. Furthermore, glycerol-3-phosphate acyltransferase catalyzed to form lyso-phosphatidic acid (LPA). Using lysophosphatidic acid acyltransferase (LPAAT), when FFA transports LPA, phosphatidic acid (PA) is formed. The enzyme phosphatidic acid phosphatase (PAP) exploits dephosphorylation of PA to form DAG. Finally, the TAG makes when locus three of DAG is controlled by extra FFA with the help of diacylglycerol acyltransferase (DGAT) (Fig. 5) [45]. Currently, genetic engineering approaches have been summarized to improve the yield of oil quantity and modify the production of fatty acids. Such approaches like the removal of enzymes capable for β-oxidation of fatty acid, enhancing the stock of overcoming sugar, optimization of fatty acid chain length by including thioesterase, decrease feedback inhibition, overexpression of thioesterase, and increased in acyl-ACP concentration. It is very important to recognize the vital limiting factors which control the lipid synthesis pathway. Various trials to overexpress genes required in fatty acid production have not worked yet [44].
Future Perspectives
Biodiesel and biohydrogen both have been proposed as the clean and promising energy carriers of the future. Microalgae utilize solar energy and generate biodiesels and biohydrogen which play a key role to reduce greenhouse gases emissions. The most key and important challenges with regard to biodiesel and biohydrogen economy are its lack of storage approaches, production cost and optimization methods, which are essential to be solved. Recent study progress is concentrated on genetic engineering improving strains capability and investigate appropriate circumstances to enhance the levels of biodiesel and biohydrogen evolution. Utilizing genetic engineering tools, it may be possible to do insertions and knockouts-based study using previous modelling research efforts. For investigation of large amount data analysis of physiology and cellular metabolism, various genome-wide tools are used. Such metabolic engineering systems biology tools are including, proteomics, computational analyses, cover genomic, metabolomics, transcriptomic and analytical techniques. Research efforts achieved from such investigations can be performed in a combined way throughout genetic engineering of strain modification.
The economic utility of biodiesel and biohydrogen production may depend on proficient production approaches with significant yield, efficient storage and transport systems assuring a prominent amount of supply. Furthermore, biodiesel and biohydrogen generation may depend on the development of well-organized bioreactors, research advances, and modification in strains efficiency utilizing metabolic engineering and optimization approaches. Besides, it is also very important to focus on commercial feasibility as cost compares to conventional sources, worth of such energy in our society and social awareness and acceptance. Importantly, biodiesel and biohydrogen have been trialed in few countries to power a fleet of vehicles. Biodiesel and biohydrogen will be accepted for various industrial purposes if an economical renewable approaches of its evolution is obtained. It is a very key concern to notice that research efforts are needed to be integrated from both engineers and scientists and completely apply bioenergy as the clean and alternative energy for the future.
Conclusion
The production of various kinds of biofuels (gaseous, liquid, and solid) through different methods is potentially impacted by microalgae. Low-cost biomaterial generation has been found in microalgae, including different types of bioenergy sources, such as biohydrogen, biodiesel and bio-alcohol. Microalgae appear to be a good solution for many environmental problems. Biohydrogen production needs to be developed to enhance bioenergy strategies and commercialize production, which is a long-term goal. Photo-biological production of hydrogen yields the advantage of inevitable product extraction and separation, and it is a positive feature of microalgal biomass that will substantially develop the economic process. Biophotolysis seems interesting for the generation of hydrogen from microalgae, as hydrogen can be directly obtained by transforming solar energy. However, the production rate is too low, and the procedure has not yet been fully scaled up, even in the laboratory. Microalgae have been shown to be a potential source of biodiesel production, but still, the cost is quite high compared to petroleum-derived diesel. Policy makers and researchers have focused attention on biodiesel. It is possible to directly displace biodiesel instead of fossil diesel without any alteration of the engine. Different factors must be measured if microalgae are used to generate biofuels. The correct selection of the strain is crucial due to the lipid or carbohydrate amount, the improvement of harvesting methods, optimization of chemical and thermal methods, and the increase in the growth rate and genetic engineering. Due to genetic engineering approaches, microalgae strains can now produce lipids and increase production rates. Various species can be utilized to generate certain types of biofuels, but the production processes still need to be optimized.
Author Contributions
Liaqat Zeb: Conceptualization, Investigation, Formal analysis, Writing-original draft, Visualization. Muhammad Shafiq: Conceptualization, Writing-review & editing. Munir Ahmed: Writing-review & editing. Asaf Khan: Review & writing. Dawood Ahmad: Writing-review & editing.
The authors declare no conflict of interest.
References
- Omer AM. Energy, environment and sustainable development. Renewable and sustainable energy reviews, (2008); 12(9): 2265-2300.
- Liming H. Financing rural renewable energy: a comparison between China and India. Renewable and Sustainable Energy Reviews, (2009); 13(5): 1096-1103.
- Tredici MR. Photobiology of microalgae mass cultures: understanding the tools for the next green revolution. Biofuels, (2010); 1(1): 143-162.
- Wang K, Khoo KS, Chew KW, Selvarajoo A, Chen W-H, et al. Microalgae: The Future Supply House of Biohydrogen and Biogas. Frontiers in Energy Research, (2021); 9158.
- Srivastava RK, Shetti NP, Reddy KR, Aminabhavi TM. Biofuels, biodiesel and biohydrogen production using bioprocesses. A review. Environmental Chemistry Letters, (2020); 18(4): 1049-1072.
- Melis A. Photosynthetic H2 metabolism in Chlamydomonas reinhardtii (unicellular green algae). Planta, (2007); 226(5): 1075-1086.
- Allahverdiyeva Y, Aro E, Kosourov S (2014) Recent developments on cyanobacteria and green algae for biohydrogen photoproduction and its importance in CO 2 reduction. Bioenergy research: advances and applications: Elsevier. pp. 367-387.
- Esquível MG, Amaro HM, Pinto TS, Fevereiro PS, Malcata FX. Efficient H2 production via Chlamydomonas reinhardtii. Trends in biotechnology, (2011); 29(12): 595-600.
- Melis A, Zhang L, Forestier M, Ghirardi ML, Seibert M. Sustained photobiological hydrogen gas production upon reversible inactivation of oxygen evolution in the green AlgaChlamydomonas reinhardtii. Plant physiology, (2000); 122(1): 127-136.
- Antal TK, Krendeleva TE, Rubin AB. Acclimation of green algae to sulfur deficiency: underlying mechanisms and application for hydrogen production. Applied microbiology and biotechnology, (2011); 89(1): 3-15.
- Torzillo G, Scoma A, Faraloni C, Giannelli L Advances in the biotechnology of hydrogen production with the microalga Chlamydomonas reinhardtii. 2014; 35.
- Jayaprabakar J, Karthikeyan A, Saikiran K, N B, Joy N Comparative study of performance and emissions of a CI engine using biodiesel of microalgae, macroalgae and rice bran. 2017; 197.
- Guan Y, Deng M, Yu X, Zhang W. Two-stage photo-biological production of hydrogen by marine green alga Platymonas subcordiformis. Biochemical Engineering Journal, (2004); 19(1): 69-73.
- Minhas AK, Hodgson P, Barrow CJ, Adholeya A. A Review on the Assessment of Stress Conditions for Simultaneous Production of Microalgal Lipids and Carotenoids. Frontiers in Microbiology, (2016); 7546.
- Ohta S, Miyamoto K, Miura Y Hydrogen evolution as a consumption mode of reducing equivalents in green algal fermentation. [Chlamydomonas reinhardii; Chlorella pyrenoidosa; Chlorococcum minutum]. 2018.
- Ainas M, Hasnaoui S, Bouarab R, Abdi N, Drouiche N, et al. Hydrogen production with the cyanobacterium Spirulina platensis. Chapter: Book Name. 2017; 42.
- Noth J, Krawietz D, Hemschemeier A, Happe T Pyruvate:Ferredoxin Oxidoreductase Is Coupled to Light-independent Hydrogen Production in Chlamydomonas reinhardtii. 2012.
- Yang W, Catalanotti C, Dadamo S, Wittkopp T, Ingram-Smith C, et al. Alternative Acetate Production Pathways in Chlamydomonas reinhardtii during Dark Anoxia and the Dominant Role of Chloroplasts in Fermentative Acetate Production. 2014; 26.
- Voloshin R, Kreslavskii V, Zharmukhamedov S, S. Bedbenov V, Ramakrishna S, et al. Photoelectrochemical cells based on photosynthetic systems: a review. 2015; 2.
- Eroglu E, Melis A. Microalgal hydrogen production research. International Journal of Hydrogen Energy, (2016); 41(30): 12772-12798.
- Burlacot A, Sawyer A, Cuiné S, Auroy-Tarrago P, Blangy S, et al. Flavodiiron-Mediated O<sub>2</sub> Photoreduction Links H<sub>2</sub> Production with CO<sub>2</sub> Fixation during the Anaerobic Induction of Photosynthesis. Plant Physiology, (2018); 177(4): 1639-1649.
- Kumar Mishra V, Goswami R A review of production, properties and advantages of biodiesel. 2017.
- Rodionova M, Poudyal R, I T, Voloshin R, Zharmukhamedov S, et al. Biofuel production: Challenges and opportunities. 2017.
- Sonkar S, Mallick N An alternative strategy for enhancing lipid accumulation in chlorophycean microalgae for biodiesel production. 2018.
- Wei D, Yuan XY, Xiang W-z Comparison of methods for rapid determination of total lipid content in microalgae. 2014; 30.
- Suh W, Mishra S, Kim T-H, Farooq W, Moon M, et al. Direct transesterification of wet microalgal biomass for preparation of biodiesel. 2015; 12.
- Sun C, Xia A, Liao Q, Kumar G, Murphy J Biomass and Bioenergy: Current State. 2018.
- Driver T, Bajhaiya DA, Pittman J Potential of Bioenergy Production from Microalgae. 2014; 1.
- Faried M, Samer M, Abdelsalam E, Yousef R, Attia Y, et al. Biodiesel production from microalgae: Processes, technologies and recent advancements. 2017; 79.
- Shimako A, Tiruta-Barna L, Pigné Y, Benetto E, Navarrete Gutierrez T, et al. Environmental assessment of bioenergy production from microalgae based systems. 2016; 139.
- Palander T, Hietanen A MANAGEMENT PLANNING METHOD FOR SUSTAINABLE ENERGY PRODUCTION FROM FOREST BIOMASS: DEVELOPMENT OF AN OPTIMIZATION SYSTEM AND CASE STUDY FOR A FINNISH ENERGY PLANT. 2018; 17.
- Miao X, Wu Q Biodiesel Production From Heterotrophic Microalgal Oil. 2006; 97.
- Tyson KS Biodiesel handling and use guidelines, report of national renewable energy laboratory. 2018.
- Demirbaş A Relationship derived from physical properties of vegetable oil and biodiesel fuels. 2008; 87.
- Knothe G, Krahl J, Van Gerpen J The Biodiesel Handbook: Second Edition. 2010.
- Tan KWM, Lee YK. The dilemma for lipid productivity in green microalgae: importance of substrate provision in improving oil yield without sacrificing growth. Biotechnology for Biofuels, (2016); 9255.
- Abou-shanab R, Matter I, Kim S-N, Oh Y-K, Choi J, et al. Characterization and identification of lipid-producing microalgae species isolated from a freshwater lake. 2011; 35.
- Zhila N, S. Kalacheva G, G. Volova T Influence of nitrogen deficiency on biochemical composition of the green alga Botryococcus. 2005; 17.
- Schlagermann P, Göttlicher G, Dillschneider R, Sastre R, Posten C Composition of Algal Oil and Its Potential as Biofuel. 2012.
- Zhu LD, Li ZH, Hiltunen E. Strategies for Lipid Production Improvement in Microalgae as a Biodiesel Feedstock. BioMed Research International, (2016); 20168792548.
- Simionato D, Block MA, La Rocca N, Jouhet J, Maréchal E, et al. The Response of Nannochloropsis gaditana to Nitrogen Starvation Includes De Novo Biosynthesis of Triacylglycerols, a Decrease of Chloroplast Galactolipids, and Reorganization of the Photosynthetic Apparatus. Eukaryotic Cell, (2013); 12(5): 665-676.
- Siri-Tarino PW, Chiu S, Bergeron N, Krauss RM. Saturated Fats Versus Polyunsaturated Fats Versus Carbohydrates for Cardiovascular Disease Prevention and Treatment. Annual review of nutrition, (2015); 35517-543.
- Goncalves EC, Wilkie AC, Kirst M, Rathinasabapathi B. Metabolic regulation of triacylglycerol accumulation in the green algae: identification of potential targets for engineering to improve oil yield. Plant Biotechnology Journal, (2016); 14(8): 1649-1660.
- Guarnieri M, Nag A, Smolinski S, Darzins A, Seibert M, et al. Examination of Triacylglycerol Biosynthetic Pathways via De Novo Transcriptomic and Proteomic Analyses in an Unsequenced Microalga. 2011; 6.
- Turchetto Zolet AC, Maraschin F, Loss G, Cagliari A, Andrade C, et al. Evolutionary view of acyl-CoA diacylglycerol acyltransferase (DGAT), a key enzyme in neutral lipid biosynthesis. 2011; 11.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


