

Review Article
CRISPR/Cas9 application in tomato breeding improvement: a review
Anh Phu Nam Bui
Adv. life sci., vol. 8, no. 4, pp. 315-319, December 2021
*- Corresponding Authors: Anh Phu Nam Bui (Email: buiphunamanh@tdmu.edu.vn)
Authors' Affiliations
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
Tomato (Solanum lycopersicum) is an essential plant because of its social and economic importance. Therefore, research have been focusing on improving tomato production. The introduction of clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated proteins (CRISPR/Cas9) system provides unique opportunities to better understand the gene functions and to rapidly generate new tomato cultivars harboring desired traits such as disease resistance, better harvest quality and abiotic tolerance. This review aims to provide latest information about the application of CRISPR/Cas9 system on tomato breeding.
Keywords: CRISPR; Tomato breeding; Applied research
Introduction![]()
The tomato is a major vegetable crop that has achieved tremendous popularity over the last century. It is practically grown in every country in the world [1,2]. The tomato plant is very versatile, and the crop can be divided into two categories; fresh market tomatoes, which we are concerned with, and processing tomatoes, which are grown only outdoors for the canning industry and mechanically harvested. In both cases, world production and consumption has grown quite rapidly over the past 25 years [3].
Tomatoes, aside from being tasty, are very healthy as they are a good source of vitamins A and C. Vitamin A is important for bone growth, cell division and differentiation, for helping in the regulation of immune system and maintaining surface linings of eyes, respiratory, urinary, and intestinal tracts [4]. Vitamin C is important in forming collagen, a protein that gives structures to bones, cartilage, muscle, and blood vessels. It also helps maintain capillaries, bones and teeth and aids in the absorption of iron [5].
Currently the tomato has a higher consumption rate in more developed countries and is often referred to as a luxury crop. In Israel, for example, the tomato is such an important part of the diet that it is a major part of the food basket, which is used when calculating the consumer price index. In other words, a scarcity of tomatoes can cause the Consumer Price Index to rise and influence the inflation rate [6,7]. In developing countries, the tomato is becoming a more important part of the food basket, but the goal of the farmer is to produce quantity not quality so people can eat (what do you mean?). As varieties improve and new cultivars with better resistance to various diseases are developed, it will become easier to grow the crops in more marginal conditions and the tomato will become a more important part of the diet in poorer countries as well [8,9].
Methods![]()
Literature Search and Selection Criteria
Google Web, Google scholar, NCBI Databases and OMIC Tools were used to obtain data for this review paper. Different key words were used to retrieve the required research articles and bioinformatics-based information, such as “CRISPR/Cas9’’ and “CRISPR/Cas9 in tomato’’. Research papers consulted for this review were those published over last 10 to 15 years and information regarding CRISPR/Cas9 application in tomato was considered for current review.
Discussion![]()
Genome editing techniques and its principles
In the last few decades, progresses in breeding approaches, especially forward genetic approaches, have played vital roles in elucidating the molecular mechanism that control agriculturally important traits in tomato. The advantage of conventional plant breeding consists of increasing the availability of genetic resources for crop improvement through introgression of the desired traits [7,10,11]. However, some plants are at risk of becoming susceptible to environmental stress and losing genetic diversity. Thus, traditional cultivation methods are not sufficient to resolve global food security issues [12].
The newly developed technologies in genome-editing have overcome the limitations of traditional breeding methods in elaborating functional genomics and crop improvement in tomato. These genetic innovations provide more accurate, timesaving, efficient targeted genomic modifications, including whole-gene insertion or deletion, stacking, or pyramiding of genes, in a transgene-free manner [13-15].
Gene editing is a molecular biology technique that intentionally targets user-defined DNA sites within the genome for the purpose of elucidating functions of unknown genes. Since modified genetic information in the parental lines is passed to next generations, gene editing can be employed to purposely alter traits of agricultural importance to develop new cultivars or breeding lines [1,16]. Various gene editing techniques have been established including zinc finger nuclease (ZFN), transcription activator-like effector nuclease (TALEN) and cluster regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 (Cas9) (CRISPR/Cas9) [8]. All these tools rely on the specificity of the endonucleases that recognize and cleave DNA at desired sites to facilitate mutations induced by cellular repair mechanism. In this review, we aim to provide the latest updates of CRISPR/Cas9 application on swine breeding (what do you mean?), although TALEN and ZFN can obtain the same outcomes [3,17].
ZFN and TALEN are two early gene editing techniques that employs similar conceptual nuclease structure to introduce genetic mutation. Both systems depend on the specificity of the DNA-binding domain of zinc finger protein (ZFP) in the ZFN system and transcription activator-like effector (TALE) in the TALEN system. Since each zinc finger in the ZFP recognizes every triplet on single-strand DNA, designing 3-6 zinc finger components in combination will therefore attach to 9-18 base pairs on aimed regions to achieve specificity [18]. On the other hand, the improved targeting property of TALE relies on the programmable tandem repeat modules, of which each module specifically binds to a single base pair. The order of the tandem repeat modules can be rearranged to obtain better directing at chosen DNA sequence. After the binding to DNA region, both ZFP and TALE will orchestrate the dimerized endonuclease Fok1 to break the double strand DNA at predetermined regions [7,19].
The introduction of DSB generated by ZFN and TALEN will trigger the DNA repair mechanisms including non-homologous end-joining (NHEJ) or homologous recombination (HR) [20]. In the error prone NHEJ pathway, the two ends of the cleaved DNA are joined and ligated, resulting in the generation of insertion or deletion at the site of DSB, thus producing knock-out mutation [17]. In the HE pathway, a site-directed nuclease and an exogenous DNA template harboring homologous sequence to the DSB regions are required to facilitate the insertion of single or multiple transgenes, thereby gaining knock-in mutation. Accumulation of reports has demonstrated the successful application of ZFN [4,21].
The latest CRISPR/Cas9 is extensively employed in genome editing research thanks to its reliability, efficiency, and simplicity [22]. Basically, CRISPR/Cas9 is a RNA- mediated adaptive immune system that can be found in bacteria, and archaea [23]. This immune protection provides resistance against genetic attacks and later stores infection histories in a form of spacer sequences for future safety. These spacers function in concert with Cas9 endonuclease proteins to monitor, recognize and degrade exogenous DNA.
This process can be divided in three stages: spacer acquisition, biogenesis, and immunity. In the spacer acquisition stage, the foreign DNA is identified, captured, and embedded into the CRISPR locus in a form of spacer. Subsequently, the expression of the CRISPR/Cas9 system will be initiated in the biogenesis stage, in which the primary CRISPR-RNAs (crRNAs) is synthesized from the CRISPR locus and subsequently undergone many processes to become crRNAs. Finally, in the immunity stage, the crRNAs, together with the trans-activating RNAs (tracrRNAs), will associate with Cas9 endonuclease, forming a ribonucleotide complex. This complex will initiate interference and consequent degradation of the targeted foreign DNA by base pairing recognition mechanism and endonucleases, respectively.
It was not until the work of Jinek et al., the significant contribution of CRISPR/Cas9 technology to genome editing begins to emerge by the establishment of the programmable version of CRISPR/Cas9 [24]. This modified version of CRISPR/Cas9 is made up of the customizable single strand RNA (sgRNA), which is the fusion product of crRNA and tracrRNA, the recombinant Cas9 protein and. This combination will result in Cas9/sgRNA complex that targets and initiates DSB at specific DNA sequences. Once DSBs are introduced, NHEJ or HDR strategy is activated to repair the DNA damages, leading to gene knockout, or gene knock-in, respectively [12,25]. CRISPR/Cas9 system has been widely employed in various research model research, including Prokaryotes (Escherichia coli) [26] and Eukaryotes (Saccharomyces cerevisiae, Drosophila melanogaster, Caenorhabditis elegans, Arabidopsis thaliana, etc.) [27-30].
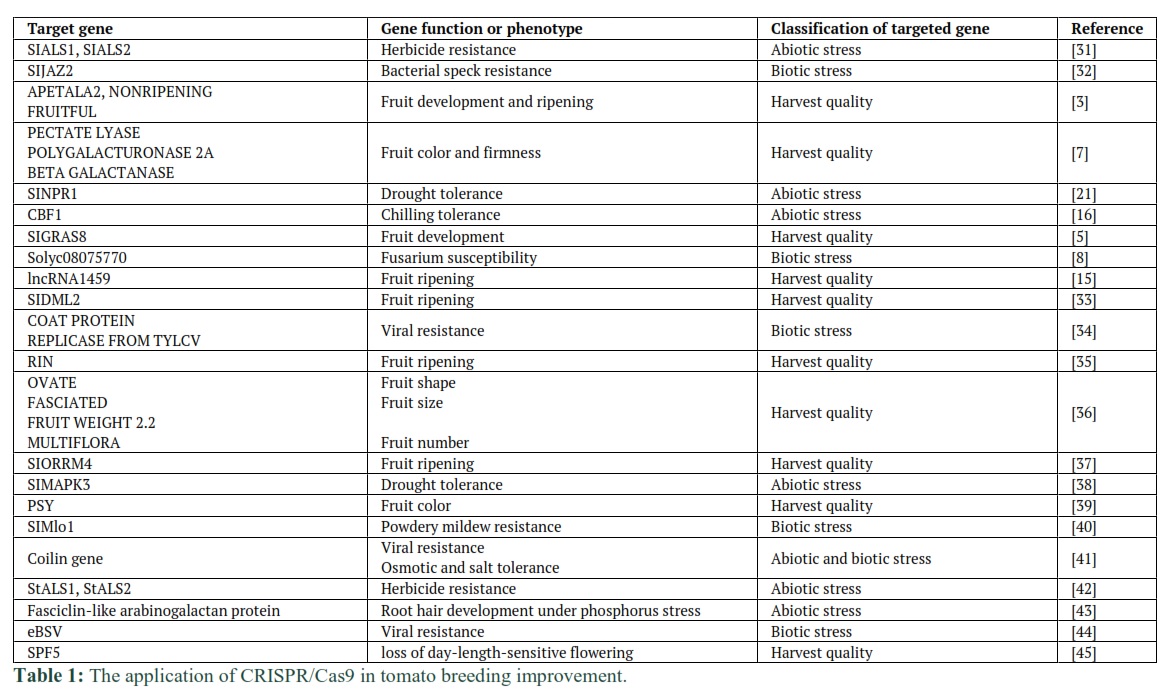
Tomato is an important source for the increasing demand for better quality and quantity for human daily consumption. As a result, tomato production is required to enhance its productivity and reduce environmental impacts. So far, a great amount of achievements have been obtained in many research. With the emergence of CRISPR/Cas9 system, tomato breeders and researchers are offered a novel tool to rapidly understand traits of great economic significance. It is hoped that CRISPR/Cas9 system will accelerate the research progress in tomato industry in the next coming decades.
Conception, revision, and final approval were done by APNB.
The author would like to thank Institute of Applied Science, Thu Dau Mot University, Vietnam for providing their help.
The author declared that present study was performed in absence of any conflict of interest.
References ![]()
- Boase M, Tiffin H, Zhang H, Hunter D, Erridge Z, et al. Gene editing of tomato via Agrobacterium-mediated transformation with CRISPR/Cas9 constructs targeting cell wall genes. 2018; 32-34.
- Chen L, Yang D, Zhang Y, Wu L, Zhang Y, et al. Evidence for a specific and critical role of mitogen-activated protein kinase 20 in uni-to-binucleate transition of microgametogenesis in tomato. New Phytologist, (2018); 219(1): 176-194.
- Wang R, Tavano ECdR, Lammers M, Martinelli AP, Angenent GC, et al. Re-evaluation of transcription factor function in tomato fruit development and ripening with CRISPR/Cas9-mutagenesis. Scientific Reports, (2019); 9(1): 1696.
- Li X, Wang Y, Chen S, Tian H, Fu D, et al. Lycopene Is Enriched in Tomato Fruit by CRISPR/Cas9-Mediated Multiplex Genome Editing. Frontiers in Plant Science, (2018); 9(559): 1-12.
- Hu N, Xian Z, Li N, Liu Y, Huang W, et al. Rapid and user-friendly open-source CRISPR/Cas9 system for single- or multi-site editing of tomato genome. Horticulture Research, (2019); 6(1): 7.
- Ding F, Wang M, Zhang S. Sedoheptulose-1,7-Bisphosphatase is Involved in Methyl Jasmonate- and Dark-Induced Leaf Senescence in Tomato Plants. International journal of molecular sciences, (2018); 19(11): 3673.
- Wang D, Samsulrizal NH, Yan C, Allcock NS, Craigon J, et al. Characterization of CRISPR Mutants Targeting Genes Modulating Pectin Degradation in Ripening Tomato. Plant Physiology, (2019); 179(2): 544-557.
- Prihatna C, Barbetti MJ, Barker SJ. A Novel Tomato Fusarium Wilt Tolerance Gene. Frontiers in Microbiology, (2018); 9(1226).
- D’Ambrosio C, Stigliani AL, Giorio G. CRISPR/Cas9 editing of carotenoid genes in tomato. Transgenic Research, (2018); 27(4): 367-378.
- Wei F-J, Droc G, Guiderdoni E, Hsing Y-IC. International Consortium of Rice Mutagenesis: resources and beyond. Rice (New York, NY), (2013); 6(1): 39-39.
- Li G, Chern M, Jain R, Martin Joel A, Schackwitz Wendy S, et al. Genome-Wide Sequencing of 41 Rice (Oryza sativa L.) Mutated Lines Reveals Diverse Mutations Induced by Fast-Neutron Irradiation. Molecular Plant, (2016); 9(7): 1078-1081.
- Yang N, Wang R, Zhao Y. Revolutionize Genetic Studies and Crop Improvement with High-Throughput and Genome-Scale CRISPR/Cas9 Gene Editing Technology. Molecular plant, (2017); 10(9): 1141-1143.
- Romero FM, Gatica-Arias A. CRISPR/Cas9: Development and Application in Rice Breeding. Rice Science, (2019); 26(5): 265-281.
- Ran FA, Hsu PD, Wright J, Agarwala V, Scott DA, et al. Genome engineering using the CRISPR-Cas9 system. Nature Protocols, (2013); 8(11): 2281-2308.
- Li R, Fu D, Zhu B, Luo Y, Zhu H. CRISPR/Cas9-mediated mutagenesis of lncRNA1459 alters tomato fruit ripening. The Plant Journal, (2018); 94(3): 513-524.
- Li R, Zhang L, Wang L, Chen L, Zhao R, et al. Reduction of Tomato-Plant Chilling Tolerance by CRISPR–Cas9-Mediated SlCBF1 Mutagenesis. Journal of Agricultural and Food Chemistry, (2018); 66(34): 9042-9051.
- Dahan-Meir T, Filler-Hayut S, Melamed-Bessudo C, Bocobza S, Czosnek H, et al. Efficient in planta gene targeting in tomato using geminiviral replicons and the CRISPR/Cas9 system. The Plant Journal, (2018); 95(1): 5-16.
- Forsyth A, Weeks T, Richael C, Duan H. Transcription Activator-Like Effector Nucleases (TALEN)-Mediated Targeted DNA Insertion in Potato Plants. Frontiers in Plant Science, (2016); 7(1572).
- Tomlinson L, Yang Y, Emenecker R, Smoker M, Taylor J, et al. Using CRISPR/Cas9 genome editing in tomato to create a gibberellin-responsive dominant dwarf DELLA allele. Plant biotechnology journal, (2019); 17(1): 132-140.
- Ito Y, Nishizawa-Yokoi A, Endo M, Mikami M, Toki S. CRISPR/Cas9-mediated mutagenesis of the RIN locus that regulates tomato fruit ripening. Biochemical and Biophysical Research Communications, (2015); 467(1): 76-82.
- Li R, Liu C, Zhao R, Wang L, Chen L, et al. CRISPR/Cas9-Mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance. BMC plant biology, (2019); 19(1): 38-38.
- Mishra R, Joshi RK, Zhao K. Genome Editing in Rice: Recent Advances, Challenges, and Future Implications. Frontiers in Plant Science, (2018); 9(1361).
- Xie K, Yang Y. RNA-Guided Genome Editing in Plants Using a CRISPR–Cas System. Molecular Plant, (2013); 6(6): 1975-1983.
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science (New York, NY), (2012); 337(6096): 816-821.
- Xu R, Wei P, Yang J (2017) Use of CRISPR/Cas Genome Editing Technology for Targeted Mutagenesis in Rice. In: Reeves A, editor. In Vitro Mutagenesis: Methods and Protocols. New York, NY: Springer New York. pp. 33-40.
- Jiang Y, Chen B, Duan C, Sun B, Yang J, et al. Multigene Editing in the Escherichia coli Genome via the CRISPR-Cas9 System. Applied and Environmental Microbiology, (2015); 81(7): 2506-2514.
- Laughery MF, Wyrick JJ. Simple CRISPR-Cas9 Genome Editing in Saccharomyces cerevisiae. Current Protocols in Molecular Biology, (2019); 129(1): e110.
- Gratz SJ, Rubinstein CD, Harrison MM, Wildonger J, O'Connor-Giles KM. CRISPR-Cas9 Genome Editing in Drosophila. Current protocols in molecular biology, (2015); 11131.32.31-31.32.20.
- Dickinson DJ, Goldstein B. CRISPR-Based Methods for Caenorhabditis elegans Genome Engineering. Genetics, (2016); 202(3): 885-901.
- Miki D, Zhang W, Zeng W, Feng Z, Zhu J-K. CRISPR/Cas9-mediated gene targeting in Arabidopsis using sequential transformation. Nature Communications, (2018); 9(1): 1967.
- Danilo B, Perrot L, Mara K, Botton E, Nogué F, et al. Efficient and transgene-free gene targeting using Agrobacterium-mediated delivery of the CRISPR/Cas9 system in tomato. Plant Cell Reports, (2019); 38(4): 459-462.
- Ortigosa A, Gimenez-Ibanez S, Leonhardt N, Solano R. Design of a bacterial speck resistant tomato by CRISPR/Cas9-mediated editing of SlJAZ2. Plant biotechnology journal, (2019); 17(3): 665-673.
- Lang Z, Wang Y, Tang K, Tang D, Datsenka T, et al. Critical roles of DNA demethylation in the activation of ripening-induced genes and inhibition of ripening-repressed genes in tomato fruit. Proceedings of the National Academy of Sciences, (2017); 114(22): E4511-E4519.
- Tashkandi M, Ali Z, Aljedaani F, Shami A, Mahfouz MM. Engineering resistance against Tomato yellow leaf curl virus via the CRISPR/Cas9 system in tomato. Plant Signaling & Behavior, (2018); 13(10): e1525996.
- Jung YJ, Lee GJ, Bae S, Kang KK. Reduced Ethylene Production in Tomato Fruits upon CRSPR/Cas9-mediated LeMADS-RIN Mutagenesis. Horticultural Science and Technology, (2018); 36396-405.
- Zsögön A, Čermák T, Naves ER, Notini MM, Edel KH, et al. De novo domestication of wild tomato using genome editing. Nature Biotechnology, (2018); 36(12): 1211-1216.
- Yang Y, Zhu G, Li R, Yan S, Fu D, et al. The RNA Editing Factor SlORRM4 Is Required for Normal Fruit Ripening in Tomato. Plant Physiology, (2017); 175(4): 1690-1702.
- Wang L, Chen L, Li R, Zhao R, Yang M, et al. Reduced Drought Tolerance by CRISPR/Cas9-Mediated SlMAPK3 Mutagenesis in Tomato Plants. Journal of Agricultural and Food Chemistry, (2017); 65(39): 8674-8682.
- Filler Hayut S, Melamed Bessudo C, Levy AA. Targeted recombination between homologous chromosomes for precise breeding in tomato. Nature Communications, (2017); 8(1): 15605.
- Nekrasov V, Wang C, Win J, Lanz C, Weigel D, et al. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Scientific Reports, (2017); 7(1): 482.
- Makhotenko AV, Khromov AV, Snigir EA, Makarova SS, Makarov VV, et al. Functional Analysis of Coilin in Virus Resistance and Stress Tolerance of Potato Solanum tuberosum using CRISPR-Cas9 Editing. Doklady Biochemistry and Biophysics, (2019); 484(1): 88-91.
- Veillet F, Perrot L, Chauvin L, Kermarrec M-P, Guyon-Debast A, et al. Transgene-Free Genome Editing in Tomato and Potato Plants Using Agrobacterium-Mediated Delivery of a CRISPR/Cas9 Cytidine Base Editor. International Journal of Molecular Sciences, (2019); 20(2): 402.
- Kirchner TW, Niehaus M, Rössig KL, Lauterbach T, Herde M, et al. Molecular Background of Pi Deficiency-Induced Root Hair Growth in Brassica carinata – A Fasciclin-Like Arabinogalactan Protein Is Involved. Frontiers in Plant Science, (2018); 9(1372): 1-17.
- Tripathi JN, Ntui VO, Ron M, Muiruri SK, Britt A, et al. CRISPR/Cas9 editing of endogenous banana streak virus in the B genome of Musa spp. overcomes a major challenge in banana breeding. Communications Biology, (2019); 2(1): 46.
- Soyk S, Müller NA, Park SJ, Schmalenbach I, Jiang K, et al. Variation in the flowering gene SELF PRUNING 5G promotes day-neutrality and early yield in tomato. Nature Genetics, (2017); 49(1): 162-168.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


