

Full Length Research Article
In silico analysis to reveal underlying trans differentiation mechanism of Mesenchymal Stem Cells into Osteocytes
Muhammad Shahid Javaid1, Haiba Kaul2, Numan Fazal1, Faiza Yaqub1, Nadia Naseer1, Muzaffar Hanif1, Noreen Latief1*
Adv. life sci., vol. 8, no. 4, pp. 412-418, December 2021
*– Corresponding Author: Noreen Latief (Email: noreen.latief@cemb.edu.pk)
Authors' Affiliations
2. University of Veterinary and Animal Sciences, Lahore, Punjab – Pakistan
Editorial Expression of Concern
08 July 2025: Following publication of this paper, the internal audit (consequent to concerns on quality raised by Web of Science) notified Advancements in Life Sciences about problems in use of English language. By this Editorial Expression of Concern, we alert the scientific community as we address the errors.
Editorial Note
25 July 2025: You are viewing the latest version of this article having minor corrections related to the use of English language. Expression of concern is hereby revoked.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
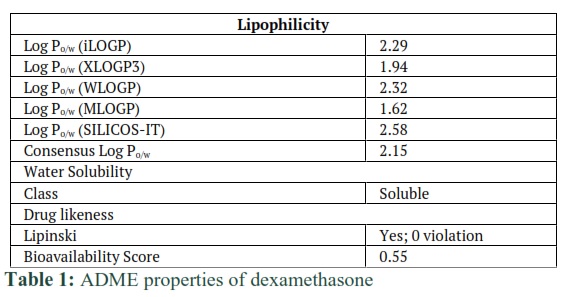
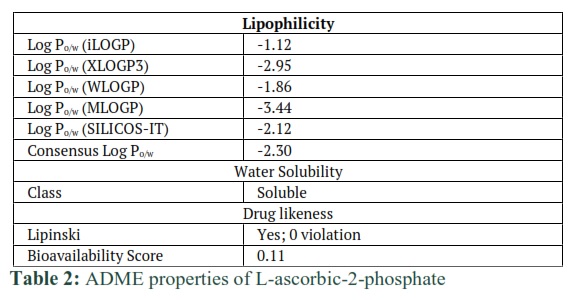
Background: Bone is a mineralized dynamic tissue, helps to protect and support the body. Osteoarthritis damages the cartilage and is responsible for the degeneration of the bone. Many cell-based therapies are available to repair the damage however, the non-availability of autologous cells and slows healing during regeneration of the damaged bone present major constraints. Hence, there is a need to search for a convenient and easily available cell source that can not only be used to repair the bone but can also enhance its regenerative potential. β-glycerophosphate, dexamethasone, and L-ascorbic-2-phosphate can differentiate mesenchymal stem cells (MSCs) into osteocytes. So far, the interaction of these compounds with osteocytes-specific proteins has not been studied. In this study, in silico analysis was performed to investigate the interaction of proteins with osteocytes specific compounds at the amino acids level.
Methods: 3D structures of Dexamethasone and L-ascorbic-2-phosphate (ascorbic acid) were drawn using Molecular Operating Environment (MOE). Then absorption, distribution, metabolism, and excretion (ADME) analysis was achieved using an online tool of “Swiss Package”. By Ramachandran plot, the predicted model of ALPL, MMP13, Osteonectin, and RunX2 proteins were evaluated. Then docking of these proteins with Dexamethasone and L-ascorbic-2-phosphate was performed.
Results: L-ascorbic-2-phosphate and Dexamethasone docked within the binding pockets of ALPL, RunX2, MMP13, and Osteonectin proteins, expressed in the bone cells. These compounds also showed good drug-likeness and pharmacokinetics properties.
Conclusion: It is concluded that β-glycerophosphate, dexamethasone, and L-ascorbic-2-phosphate are novel substrates for osteogenic differentiation. These compounds could increase the healing and regenerative potential of bone cells by enhancing the expression of osteocytes specific proteins.
Keywords: Bone; Osteoarthritis; β-glycerophosphate; Dexamethasone; L-ascorbic-2-phosphate; Docking; Differentiation; Mesenchymal stem cells (MSCs); Osteonectin
Introduction![]()
Bone gives shape to the body and also protects internal delicate organs against external insult [1]. Bone is affected by different types of stimuli and factors [2, 3] such as cell-cell communication via gap junction, paracrine, endocrine, and autocrine factors [4]. The remodeling process is controlled by organized and coordinated interactions between different bone cells [5]. Bone cells consist of osteocytes, osteoclasts, and osteoblasts [6]. Osteocytes are present in abundant numbers and found in the lacunar space of the bone matrix. Lacunar-canalicular network is made by osteocytes and dendrites [7, 8] which connect osteocytes as well as with other cells on the bone surface and thus communicate extra and intracellular [9].
Osteoarthritis is a degenerative disease of joints that affects millions of people globally [10]. Articular cartilage covers the ends of joints and helps in the frictionless movement of bones. Degradation of articular cartilage results in osteoarthritis [11]. Degraded cartilage also contributes to bone degeneration. Extracellular matrix (ECM) maintains the intact cartilage. Disturbed homeostasis of ECM is the main cause of the degradation of cartilage and bones. Other factors responsible for osteoarthritis and ultimately degeneration of cartilage and bones are age [12], wear and tear, nutrient deficiency, environmental factors [13], and oxidative stress [14]. Properties of bone change with age, environmental factors, and deficiency of minerals and vitamins. Aging is a common factor for a high prone risk of bone diseases. It was reported that older persons have 10-fold greater risks of bone diseases compared to younger ones [15]. Bones have slow healing and an inefficient regenerative process that further slows down with age.
Stem cell-based therapies are a new hope to repair and regenerate damaged bone. Osteocytes are the main types of bone cells as they make 95% of bone [16] and differentiate into osteoblasts. Isolation of autologous osteocytes is an invasive method. We can use alternate methods to differentiate stem cells into osteocytes. Adipose stem cells are strong candidates as they have great potential to differentiate into osteocytes as described previously [17]. Triggering the osteogenic pathway requires inhibition of the adipogenic pathway [18]. β–glycerophosphate, dexamethasone, and ascorbic acid-2-phosphate play a key role in this purpose. In vitro experiments have confirmed osteogenic differentiation using these compounds but in silico interactions of osteocyte-specific proteins with these synthetic compounds have not been investigated yet.
We selected osteocytes specific proteins, RunX2, MMP13, Osteonectin, and ALPL. In silico approach was used to study the interaction of osteocyte-specific proteins with in vitro utilized differentiated compounds like Dexamethasone and L-ascorbic-2-phosphate. Molecular docking of ligands with target proteins was performed using different web-based and desktop-based bioinformatics tools. 3D structures of proteins were predicted, and 3D structures of compounds were prepared by desktop-based tool “MOE” (Molecular Operating Environment (MOE), 2019.01; Chemical Computing Group ULC). Structures were validated and docking was performed using “MOE”. Results showed that RunX2, MMP13, Osteonectin, and ALPL proteins interact with ligands at the molecular level.
This study was performed to examine the amino acids interactions of osteocytes specific proteins with β–glycerophosphate, dexamethasone, and L-ascorbic-2-phosphate. Docking results manifested the role of ligand compounds to enhance the expression of osteocytes specific proteins. In the future, results of this in silico study would help in better understanding the role of dexamethasone and L-ascorbic-2-phosphate in the regeneration of damaged bone by targeting the ALPL, MMP13, Osteonectin, and RunX2 proteins.
Methods![]()
Preparation of ligand structure
Dexamethasone and L-ascorbic-2-phosphate (ascorbic acid) were used as ligands. Molecular Operating Environment (MOE) was used to draw their 3D structures from their molecular formulas [19]. Hydrogen atoms with minimized energy were added to ligands structures and ligand database files were prepared. Potential energy was calculated.
ADME analysis
Absorption, Distribution, Metabolism, and Excretion (ADME) analysis of dexamethasone and L-ascorbic-2-phosphate was performed by using an online tool of “Swiss Package” [19].
Retrieval of amino acids Sequences
Protein sequences of ALPL, MMP13, Osteonectin, and RunX2 of rat were retrieved from NCBI under GenBank ID CAA34160.1, EDL78538.1, CAA74042.1, and NP_001265412.1 respectively.
Prediction and evaluation of the 3D structure of proteins
For docking, 3D structures of ALPL, MMP13, Osteonectin, and RunX2 were predicted using an online prediction tool. Optimized parameters were used to predict the three-dimensional structures of these proteins. Threading and homology modeling approaches were used. Predicted models were evaluated by the Ramachandran plot.
Interaction of proteins with ligands
Protein structures were used as the receptors. 3D structures of these proteins were opened in MOE. Hydrogen atoms were connected to them, and structures were analyzed to identify ligand binding sites. Then ligands were docked by opening the database file of ligand structures.
Results
![]()
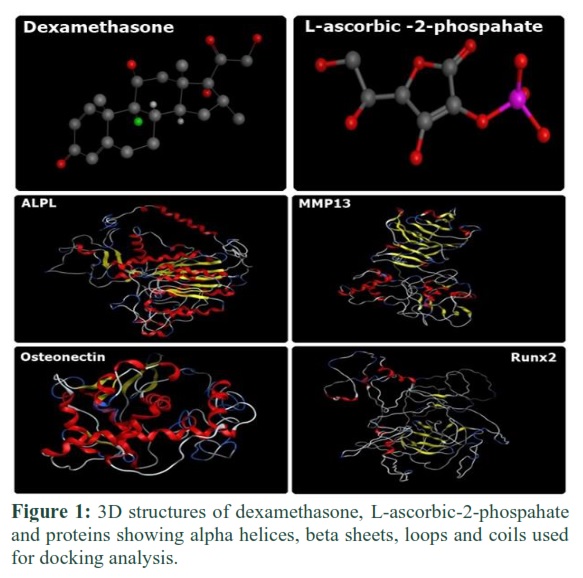
3D structures of proteins and ligands
MOE generated 3D structures of ligand and receptors molecules are shown in Figure 1. 3D structures of proteins showed beta-sheets, alpha helices, loops, and coils.
ADME analysis of ligands
Computed ADME properties of dexamethasone and L-ascorbic-2-phosphate were shown in tables 1 and 2 respectively. Lipophilicity, water solubility, and drug likeliness of dexamethasone and L-ascorbic-2-phosphate were checked against threshold values that are drawn from algorithms and saved in the database. ADME results have shown that these compounds are safe to use (Table 1 and 2).
Evaluation of predicted 3D structures of proteins
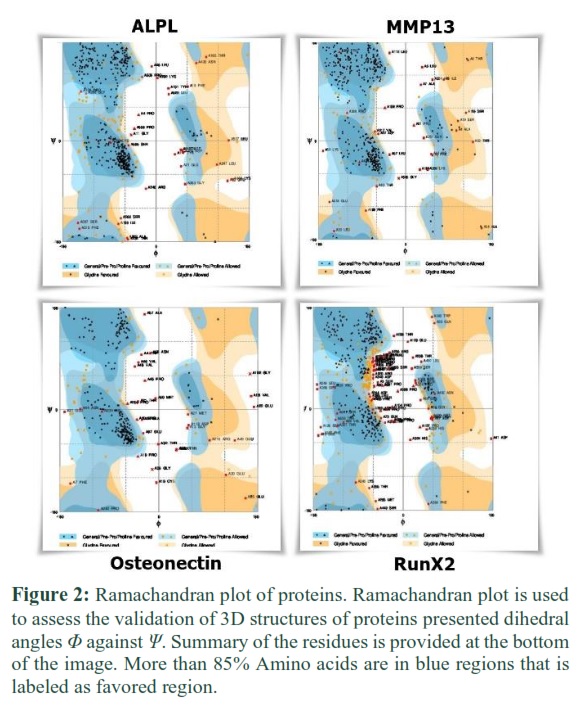
Predicted 3D structures of proteins were evaluated based on the Ramachandran plot. It was determined by the Ramachandran plot that 418 amino acids (80.1%) of ALPL protein were in the favored region and only 32 (6.1%) amino acids were in the outlier region. 383 (81.5%) amino acids of MMP13 protein were in the favored region and 30 amino acids (6.4%) were in the outlier region respectively. 222 (74.2%) amino acids of Osteonectin protein were in the favored region while 33 (11.0%) amino acids were in the outlier region. 306 (58.1%) amino acids of RunX2 were in the favored and 69 (13.1%) in the outlier region. Based on threshold criteria, all the structures were validated as more than 85 % amino acids of all the structures were in the favored and allowed region. (Figure 2)
Protein ligand docking
Ligands were docked with receptors molecules via 30 different angles. This different angular geometry was used by MOE in order to best fit the ligand inside the proximity of the receptor’s binding pocket.
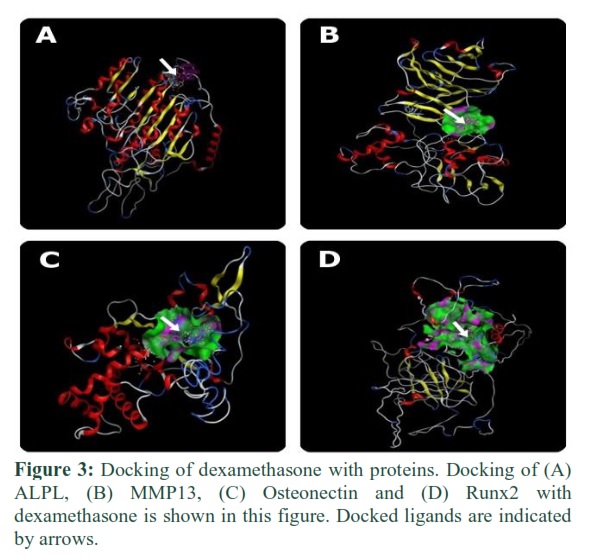
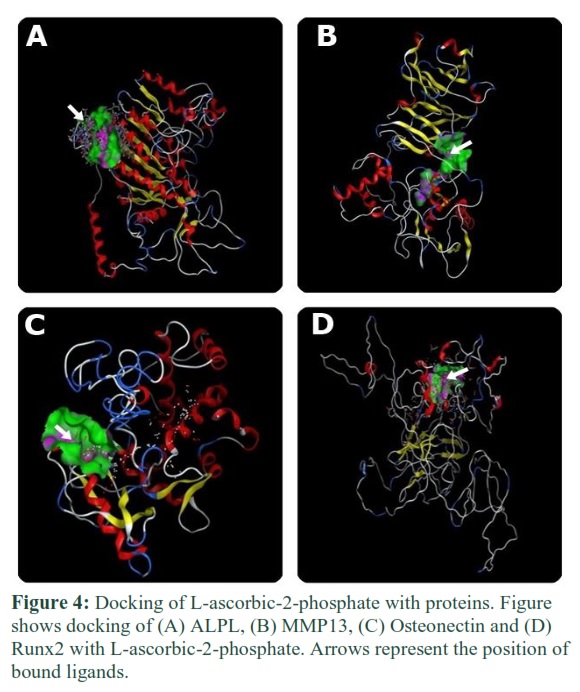
Docking of ALPL, MMP13, Osteonectin, and Runx2 with dexamethasone and L-ascorbic phosphate
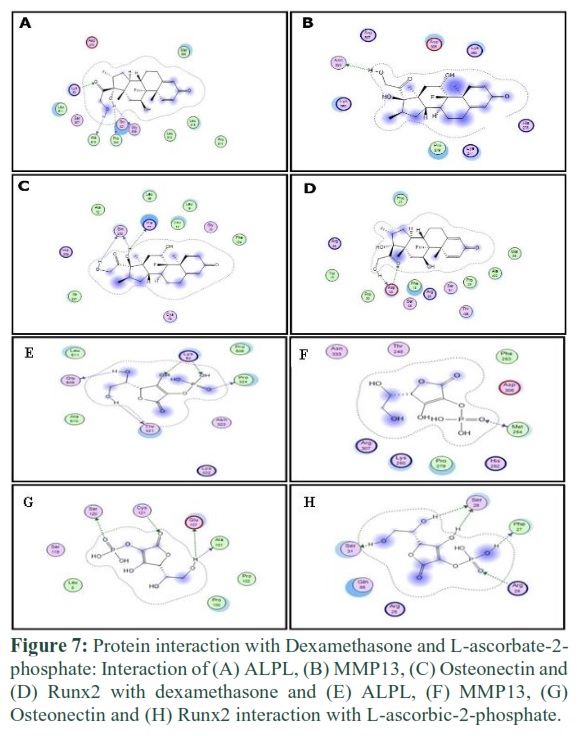
Dexamethasone and L-ascorbic-2-phosphate were bound to proteins via specific amino acid residues. These amino acids made the binding pocket of proteins and came into contact with dexamethasone and L-ascorbic-2-phosphate. In figures 3 and 4, green bubbles represent the binding pocket, and the position of docked ligands is indicated by arrows.
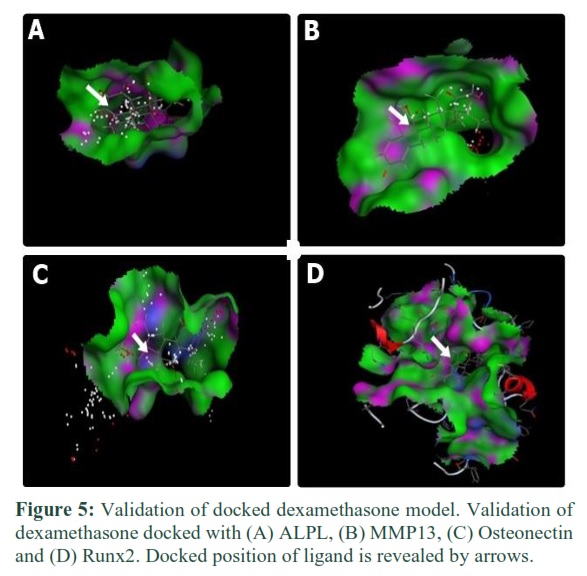
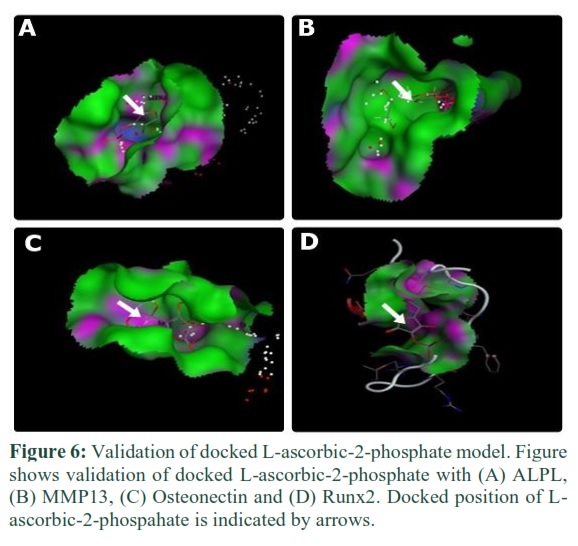
Validation
Single docking interaction gave 30 docked models. Validation was done to investigate that which model of protein has ligand within the proximity of the binding site. Each model was examined one by one on account of its docked position inside the binding pocket. For ALPL, among 30 docked models, model no. 4 of dexamethasone and model no. 1 of L-ascorbic-2-phosphate was selected by the validation process. For MMP13, model no.6 of dexamethasone and model no.2 of L-ascorbic-2-phosphate and for Osteonectin, model no.2 of dexamethasone and model no.1 of L-ascorbic-2-phosphate were selected. For Runx2, model no.20 of dexamethasone and model no.6 of L-ascorbic-2-phosphate was selected by the same validation process. Validation is shown in figures 5 and 6.
Protein ligand interaction
Protein ligand interaction was performed using MOE to determine the number and name of amino acids that interacted with dexamethasone and L-ascorbic-2-phosphate.
ALPL interaction with dexamethasone and L-ascorbic-2-phosphate
Lys 52, Ala 510, Pro 506, Thr 321, and Gly 509 of ALPL interacted with dexamethasone (Figure 7A). While amino acids Gly 509, Thr 321, Pro 324, and Lys 52 of ALPL showed interaction with L-ascorbic-2-phosphate.
MMP13 interaction with dexamethasone and L-ascorbic-2-phosphate
Amino acid Asn 333 and amino acid Met 254 of MMP13 interacted with dexamethasone and L-ascorbic-2-phosphate respectively (Figure 7B and 8B).
Osteonectin interaction with dexamethasone and L-ascorbic-2-phosphate
Amino acids Gln 238, His 77, and amino acids Ser 120, Cys 121, Glu 107, and Ala 101 of Osteonectin protein were interacted with dexamethasone and L-ascorbic-2-phosphate respectively (Figure 7C and 8C).
RunX2 interaction with dexamethasone and L-ascorbic-2-phosphate
Amino acid Asp 20 and amino acids Ser 31, Ser 28, Phe 27, and Arg 26 of RunX2 protein were interacted with dexamethasone and L-ascorbic-2-phosphate respectively (Figure 7D and 8D).
Figures & Tables
Osteogenic differentiation of adipose stem cells requires dexamethasone, L-ascorbic-2-phosphate, and β-glycerophosphate. Dexamethasone, a synthetic glucocorticoid, is an anti-inflammatory substance and was used for osteogenic differentiation [20]. L-ascorbic-2-phosphate enhances collagen accumulation, the proliferation of cells, and the ratio of GAGs content. β-glycerophosphate acts as a phosphate donor [21].
It was reported that dexamethasone triggers the osteogenic differentiation of MSCs by upregulating the expression of RunX2 dependent WNT/ β-catenin singling [22]. It has been reported that up-regulation of RunX2 resulted in the high expression of collagen type I [23]. Besides WNT/ β-catenin, TAZ (transcriptional co-activator with PDZ-binding motif) can also be activated by dexamethasone [24-26] and it can enhance the expression of RunX2 by repressing the expression of adipogenic differentiation markers of stem cells [24]. Dexamethasone also interacts with the mitogen-activated protein kinase phosphatase (MKP-1) pathway and induces osteocytes differentiation. By the MKP-1 pathway, RunX2 enhances the expression of some other bone-related proteins such as bone sialo protein and osteocalcin [27]. In vitro studies have reported the role of dexamethasone, L-ascorbic-2-phosphate, and β-glycerophosphate in molecular pathways of osteogenic differentiation but how exactly they bind and interact is not yet so clear.
In this study, in silico approach was used to explicate the binding interaction of dexamethasone and L-ascorbic-2-phosphate with osteocytes specific proteins and transcription factors. The binding of dexamethasone and L-ascorbic-2-phosphate was investigated with RunX2 (Figure 3D and 4D). Dexamethasone binds with the amino acids of RunX2 which are Phe, Arg, and Ser, whereas L-ascorbic-2-phosphate interacts with proline and lysine amino acids and can enhance the secretion of collagen type I [28]. These compounds can bind and change the 3D conformation of RunX2 protein that could enhance its activation. Β-Glycerophosphate did not bind with any protein as it has an indirect role. It gives phosphates that are required in osteocytes specific pathways, initiated in response to dexamethasone and L-ascorbic-2-phosphate.
Osteonectin protein is also called SPARC [29]. It is responsible for normal remodeling of bone as well as maintenance of bone mass [30]. Its expression must be increased in osteogenic differentiation to increase the regeneration of bone. It could be achieved by hyper-activating the expression of this protein using the osteogenic-specific compounds. Docking has described the binding of L-ascorbic-2-phosphate and dexamethasone with Osteonectin (Figure 3C and 4C).
MMP13 plays a role in fracture resistance and in the maintenance of bone quality. Expression of MMP13 is required for the development of long bone [31]. ALPL is responsible for bone calcification. It was observed that dexamethasone and L-ascorbic-2-phosphate bind with these proteins (Figure 3B, 4B, and Figure 3A, 4A) so that they can play their role to differentiate adipose stem cells into osteocytes.
Besides interaction, it was also necessary to check the drug likeliness, toxicity, absorption, metabolism, distribution, and excretion properties of dexamethasone and L-ascorbic-2-phosphate. ADME analysis showed that these substances are water-soluble and safe to use as they have no violation of the Lipinski rule of five, a standard algorithm to check the drug-like properties of a compound. This study reveals the interaction of osteocytes-specific proteins and compounds at the molecular level. In the future, it would be helpful to modify the structures of these compounds or to mutate the osteocytes-specific proteins in such a way that could increase the regenerative potential of bone.
Author Contributions
Conceptualization, Javaid MS and Latief N; Methodology, Kaul H and Fazal N; Validation, Yaqub F and Naseer N; Writing-original draft preparation, Javaid MS and Hanif M; Writing-Review and editing, Javaid MS, Kaul H, and Latief N; Supervision, Latief N.
The authors declare that there is no conflict of interest regarding the publication of this paper.
References
- Phan T, Xu J, Zheng M. Interaction between osteoblast and osteoclast: impact in bone disease. Histology and histopathology, (2004); 19(4): 1325-44.
- Sims NA, Walsh NC. Intercellular cross-talk among bone cells: new factors and pathways. Current osteoporosis reports, (2012); 10(2): 109-117.
- Stains JP, Civitelli R. Gap junctions in skeletal development and function. Biochimica et Biophysica Acta (BBA)-Biomembranes, (2005); 1719(1-2): 69-81.
- Watkins M, Grimston SK, Norris JY, Guillotin B, Shaw A, et al. Osteoblast connexin43 modulates skeletal architecture by regulating both arms of bone remodeling. Molecular biology of the cell, (2011); 22(8): 1240-1251.
- Del Fattore A, Teti A, Rucci N. Bone cells and the mechanisms of bone remodelling. Front Biosci (Elite Ed), (2012); 42302-2321.
- Neve A, Corrado A, Cantatore FP. Osteoblast physiology in normal and pathological conditions. Cell and tissue research, (2011); 343(2): 289-302.
- Clarke B. Normal bone anatomy and physiology. Clinical journal of the American Society of Nephrology, (2008); 3(Supplement 3): S131-S139.
- Pajevic PD. Regulation of bone resorption and mineral homeostasis by osteocytes. Ibms Bonekey, (2009); 663.
- Komori T. Functions of the osteocyte network in the regulation of bone mass. Cell and tissue research, (2013); 352(2): 191-198.
- Akkiraju H, Nohe A. Role of chondrocytes in cartilage formation, progression of osteoarthritis and cartilage regeneration. Journal of developmental biology, (2015); 3(4): 177-192.
- Glasson SS, Askew R, Sheppard B, Carito B, Blanchet T, et al. Deletion of active ADAMTS5 prevents cartilage degradation in a murine model of osteoarthritis. Nature, (2005); 434(7033): 644-648.
- Hochberg MC (2002) Prevention of lower limb osteoarthritis: data from the Johns Hopkins Precursors Study. The many faces of Osteoarthritis: Springer. pp. 31-37.
- Spector TD, MacGregor AJ. Risk factors for osteoarthritis: genetics. Osteoarthritis and cartilage, (2004); 1239-44.
- Altindag O, Erel O, Aksoy N, Selek S, Celik H, et al. Increased oxidative stress and its relation with collagen metabolism in knee osteoarthritis. Rheumatology international, (2007); 27(4): 339-344.
- Kanis JA. Diagnosis of osteoporosis and assessment of fracture risk. The Lancet, (2002); 359(9321): 1929-1936.
- Bonewald LF, Johnson ML. Osteocytes, mechanosensing and Wnt signaling. Bone, (2008); 42(4): 606-615.
- Chen Q, Shou P, Zheng C, Jiang M, Cao G, et al. Fate decision of mesenchymal stem cells: adipocytes or osteoblasts? Cell Death & Differentiation, (2016); 23(7): 1128-1139.
- Dai F, Zhang F, Sun D, Zhang Z, Dong S, et al. CTLA4 enhances the osteogenic differentiation of allogeneic human mesenchymal stem cells in a model of immune activation. Brazilian Journal of Medical and Biological Research, (2015); 48629-636.
- Javaid MS, Latief N, Ijaz B, Ashfaq UA. Epigallocatechin gallate as an anti-obesity therapeutic compound: An in silico approach for structure-based drug designing. Natural product research, (2018); 32(17): 2121-2125.
- Oshina H, Sotome S, Yoshii T, Torigoe I, Sugata Y, et al. Effects of continuous dexamethasone treatment on differentiation capabilities of bone marrow-derived mesenchymal cells. Bone, (2007); 41(4): 575-583.
- Boskey AL, Guidon P, Doty SB, Stiner D, Leboy P, et al. The mechanism of β‐glycerophosphate action in mineralizing chick limb‐bud mesenchymal cell cultures. Journal of Bone and Mineral Research, (1996); 11(11): 1694-1702.
- Gaur T, Lengner CJ, Hovhannisyan H, Bhat RA, Bodine PV, et al. Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. Journal of Biological Chemistry, (2005); 280(39): 33132-33140.
- Hamidouche Z, Haÿ E, Vaudin P, Charbord P, Schüle R, et al. FHL2 mediates dexamethasone‐induced mesenchymal cell differentiation into osteoblasts by activating Wnt/β‐catenin signaling‐dependent Runx2 expression. The FASEB Journal, (2008); 22(11): 3813-3822.
- Hong D, Chen H-X, Xue Y, Li D-M, Wan X-C, et al. Osteoblastogenic effects of dexamethasone through upregulation of TAZ expression in rat mesenchymal stem cells. The Journal of steroid biochemistry and molecular biology, (2009); 116(1-2): 86-92.
- Kanai F, Marignani PA, Sarbassova D, Yagi R, Hall RA, et al. TAZ: a novel transcriptional co‐activator regulated by interactions with 14‐3‐3 and PDZ domain proteins. The EMBO journal, (2000); 19(24): 6778-6791.
- Hong J-H, Hwang ES, McManus MT, Amsterdam A, Tian Y, et al. TAZ, a transcriptional modulator of mesenchymal stem cell differentiation. Science, (2005); 309(5737): 1074-1078.
- Phillips JE, Gersbach CA, Wojtowicz AM, García AJ. Glucocorticoid-induced osteogenesis is negatively regulated by Runx2/Cbfa1 serine phosphorylation. Journal of cell science, (2006); 119(3): 581-591.
- Vater C, Kasten P, Stiehler M. Culture media for the differentiation of mesenchymal stromal cells. Acta biomaterialia, (2011); 7(2): 463-477.
- Yiu GK, Chan WY, Ng S-W, Chan PS, Cheung KK, et al. SPARC (secreted protein acidic and rich in cysteine) induces apoptosis in ovarian cancer cells. The American journal of pathology, (2001); 159(2): 609-622.
- Delany AM, Kalajzic I, Bradshaw AD, Sage EH, Canalis E. Osteonectin-null mutation compromises osteoblast formation, maturation, and survival. Endocrinology, (2003); 144(6): 2588-2596.
- Page-McCaw A, Ewald AJ, Werb Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nature reviews Molecular cell biology, (2007); 8(3): 221-233.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0