

Full Length Research Article
NaCl affects lipids peroxidation and oxygen free radicals scavenging machinery in callus tissues of a cultivated (Solanum macrocarpon L.) and a wild Eggplant (Solanum dasyphyllum L.)
Sami Hannachi*1,2, Insaf Bahrini1, Nawaf I. Ibrahim1, Abdelmuhsin Abdelgadir1, Hira Affan Siddiqui3
Adv. life sci., vol. 8, no. 4, pp. 396-405, December 2021
*- Corresponding Author: Sami Hannachi (Email: sami.hannachi@gmail.com)
Authors' Affiliations
2. Department of Plants and Crops, Faculty of Bioscience Engineering, Ghent University, Coupure links, 653, 9000 Ghent – Belgium
3. Department of Physics, College of Science, University of Hail, PO Box 2440. Ha'il – 81451 – Kingdom of Saudi Arabia
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: In vitro techniques are an efficient tool to select salt tolerant lines in several plant species. The impacts of increasing salt concentration on membrane lipids oxidation and oxygen free radicals scavenging enzymes were evaluated in the callus of a cultivated eggplant (Solanum macrocarpon L.) and Wild Eggplant (Solanum dasyphyllum L.)
Methods: A salt stress ranging from 40 to 160 mM NaCl was imposed to callus of S. macrocarpon ‘Akwaseho’, a cultivated African eggplant and callus of S. dasyphyllum var dasyphyllum, a putative wild ancestor, for 40 days. Selected callus physiology and biochemistry features were investigated after 40 days of growing both species in MS medium added with NaCl at the concentration of 0 (control), 40, 80, 120 and 160 mM.
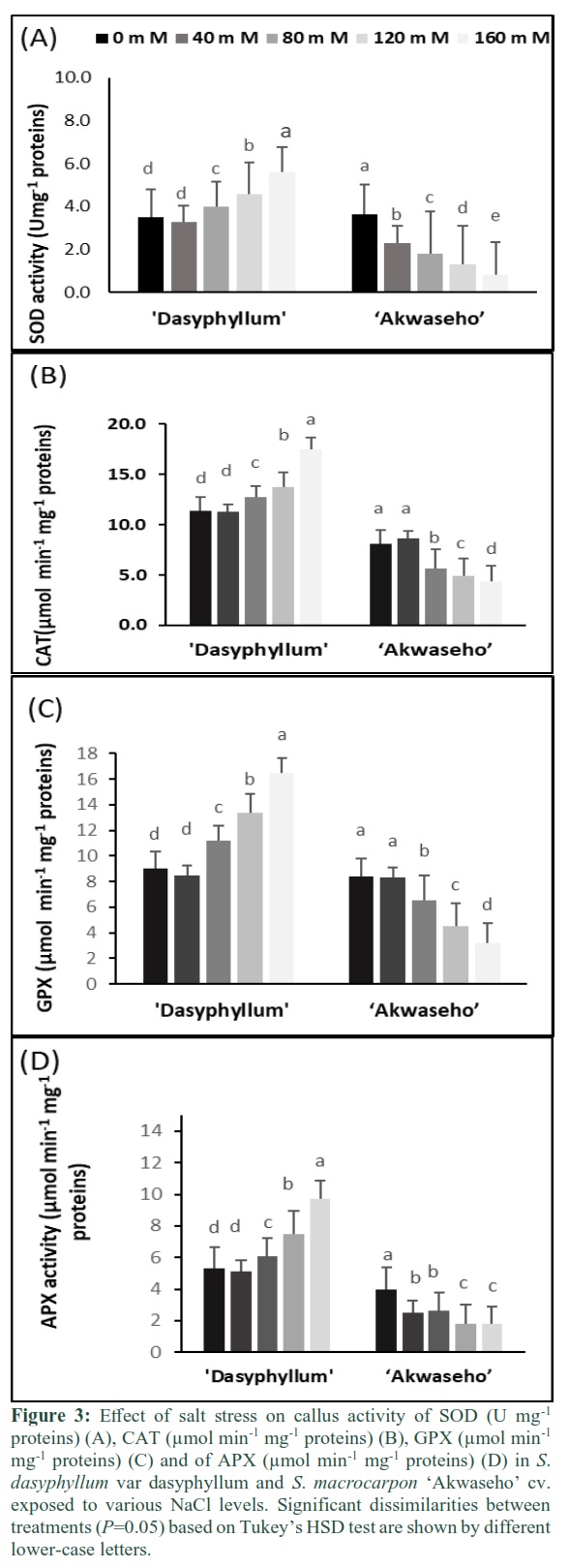
Results: A close correlation was observed between rising salinity level and the enhancement of proline accumulation. Callus of S. dasyphyllum var dasyphyllum succeeded efficiently in keeping higher K+, lower Na+ rate and Na+/K+ ratio than S. macrocarpon L. Ion content can be considered as useful tool to select salt tolerant callus tissue. Under saline condition, S. dasyphyllum L. showed lower amounts of malondialdehyde (MDA) and H2O2 but higher activity for superoxide dismutase (SOD), catalase (CAT), guaiacol peroxidase (GPX) and ascorbate peroxidase (APX) than S. macrocarpon L. in both case control and NaCl treatments.
Conclusion: The current study concluded that the two species responded differently to salinity induced oxidative damage. S. dasyphyllum L. callus showed more effective antioxidant defense system, which contributes, to better adaptive and protective capacity against salinity induced oxidative impairment by keeping more intense antioxidant enzymes activities than S. macrocarpon L. callus.
Keywords: Salinity; Tissue Culture; Oxygen Free Radicals; Eggplant
Introduction![]()
As a major global problem appearing in arid and semiarid area, soil salinity reduces plant growth, and significantly decreases crop yield [1,2]. Salt stress induces several and contrasting morphological, physiological and biochemical responses in plants [3]. The exposition of in vitro cultures to salinity has induced callus growth decline [4,5], ion imbalance and mineral deficiency, osmolytes accumulation and water status reduction [4]. Subjection to salt stress has been shown to induce the reduction in callus growth [4, 5], the accumulation of proline [4] the increase of sodium and the decrease of potassium ion concentrations [4].
In vitro culture of plant cells or tissues on medium containing selective agents constitutes an effective tool to investigate the salt induced response at callus level, decrease the time to select specific traits and minimize environmental interactions [6]. Moreover, In vitro techniques have been reported previously by several authors as an efficient tool to select and produce salt tolerant lines in numerous plant species [7]. Also in vitro procedure applications offer a deep knowledge of the mechanisms involved in stress tolerance in the entire plant [8, 9] and at callus level [10,11]. Recently, many researchers have investigated the impact of salinity in several callus biochemical process [12,13]. There is evidence that increasing salinity contributes to a substantial and continuous formation of cytotoxic oxygen free radicals which leads to an oxidative damage affecting DNA, lipids and proteins [14, 15]. In order to protect themselves from oxidative deterioration, plants have adopted an antioxidant defense machinery including both non-enzymatic (ascorbate, glutathione, α-tocopherol, etc.) and enzymatic such as APX, GPX and CAT [16].
Peroxidases (APX and GPX) catalyze the reduction of H2O2 to H2O. APX utilizes ascorbate as the electron donor in the first step of the ascorbate–glutathione cycle and is known to be the best plant peroxidase in H2O2 detoxification [17, 18]. Catalases (CAT) catalyze the reduction of H2O2 to water [19]. Several researchers reported that plants salinity tolerance is deeply linked to an effective antioxidant defense system [16]. Depending on the genotype (more tolerant or more sensitive), several differences have been found at the levels of expression and efficiency of antioxidative enzymes [9]. Lipid peroxidation being induced by free radicals has been considered as useful method assessing both oxidative deterioration and cell membrane integrity under stressful environment [17].
Eggplant is an important agronomical non-tuberous solanaceous crop. It represents an important vegetable crops all over the world and is moderately sensitive to salinity [20]. Solanum macrocarpon L. (African eggplant) is the cultivated form of the wild Solanum dasyphyllum L.
Solanum dasyphyllum L., the African ancestry of Solanum macrocarpon, are spiny wild forms which can be present in the tropical non-arid area of Africa and their fruits are still being consumed irregularly as vegetable. However, the cultivated forms (Solanum macrocarpon), called ‘gboma’ in West Africa makes an important fruit and leaf vegetable, which is oriented towards the market. Local cultivars grown for the leaves are familiar in the West and Central of Africa, while the fruit types are mainly limited to the humid coastal region of West Africa [21]. Cultivated eggplant is potentially crossable with its wild ancestor for introgression breeding aiming at the incorporation of desired specific traits [22]. Consequently, there is crucial need to identify sources of variation for tolerance to salinity among eggplant wild relatives growing in salty environments [23]. Previously, many in vitro reports have been done based on cultivated eggplant dealing with callus antioxidant enzymes activity in Solanum melongena [24] and antioxidative responses of eggplant seedlings to salinity stress [25]. However, the responses of S. macrocarpon .L and S. dasyphyllum L. under salt stress conditions have not yet been studied in callus tissue level. Nevertheless, investigating the stress tolerance level of callus tissue is essential for somaclonal selection process. On the one hand, the present work aimed at assessing the impact of the salinity on a Cultivated (S. macrocarpon L.) and a Wild Eggplant (S. dasyphyllum L.) using a biochemical and physiological approach in order to detect the mechanisms underlying the salt tolerance in both cultivars. Therefore, we analyzed several parameters such as callus growth, proline content, sodium and potassium accumulation, protein, hydrogen peroxide, lipid peroxidation, and antioxidant machinery in callus tissue grown under NaCl stress. On the other hand, another goal of this study was to focus on to what extend antioxidant machinery is associated with salt tolerance of the callus tissue in both of the cultivated and in Wild eggplant seedlings.
Methods![]()
Initiation and stock
Seeds of S. macrocarpon ‘Akwaseho’, a cultivated African eggplant (originated from Ghana) and seeds of S. dasyphyllum var dasyphyllum, a putative wild ancestor (originated from Ghana) were used as plant material, which was provided by the Biology Department of University of Hail. Thiram pre-treated seeds of both species were rinsed with 70% of alcohol for some seconds and then laved by distilled water. Later on they were subjected to a first sterilization during 20 min in a 5% HazTab solution (1,3,5 Dichloro-Triazine-Trionedihydrate-Dichlorosodium) and 0.02% Dreft (5-15% non-ionic surfactants, 15-30% anionic surfactants), then a second sterilisation in 0.5% mercuric chloride during 10 min was achieved. After being rinsed three times by sterile distilled water, the germination of seeds was performed in glass vessels (0.7 L) containing 0.8 % agar-solidified MS medium [26] with 3% (w/v) sucrose. Before autoclaving, the adjustment of the pH to 5.8 was done using 1 N NaOH. Stock plants were kept in a growth chamber at 28±2°C and a 16-h photoperiod regime which was furnished by cool-white fluorescent lamps along with a photon flux density of 36 µmol m–2 s–1.
Callus induction and salt treatment
25- to 30-day-old seedlings were selected to excise leaf segments (50 mm2). These explants were grown during 30 days on 0.8 % agar MS medium (3 % sucrose) [62] added with 0.2 µM thidiazuron (TDZ). The culture was kept in a growth chamber at 28±2°C and a 16-h photoperiod regime provided by cool-white fluorescent lamps and a photon flux density of 36 µmol m–2 s–1. Four explants per glass vessel were used and replicated four times. Calli showing better fresh weight obtained for 0.2 µM TDZ were subjected to fragmentation and then moved to MS medium containing 0.4 μM TDZ and five NaCl concentrations: 0 (control), 40, 80, 120, or 160 mM. In each glass vessel, we grew five explants, this in four repetitions. 40 days-old calli of both species were chosen for evaluating selected parameters dealing with callus physiology and biochemistry.
Growth and water relation parameters
The investigation of water status was achieved via assessing the relative water content (RWC). RWC was quantified according to [27]. Briefly, after being kept in distilled water (50ml) during 4 h a fresh callus sample (0.5g) was dried in an oven for 48h at 70°C. Then the dry weight (DW) and fresh weight (FW) were used for determination of RWC value as follow:
RWC (%) = [(FW− DW)/(TW−DW)] ×100, (TW is the turgid weight).
Proline content
We quantified proline according to [28]. In short, 0.5g of fresh callus tissue was extracted with 3% (w/v) sulfosalicylic acid. The determination of proline was carried out using a calibration curve and expressed as µg proline g-1 FW.
Ionic assay (Na+ and K+)
The extraction and the estimation of Na+ and K+ content was achieved according to [29]. In short, collected calli were dried at 60°C for 5 days, then 10 mg of grinded callus was digested with 10 ml 3% (w/v) aqueous sulfosalicylic acid for 24 h at 4°C, after that sample extract was purified with Whatman No. 1 filter paper and finally Na+ and K+ contents were determined by flame photometer (Jenway Model PEP7, UK).
Hydrogen peroxide quantification
The determination of hydrogen peroxide amount was achieved as described by [30]. The homogenization of fresh callus (1 g) was performed using 5 ml of 0.1% (w/v) trichloroacetic acid (TCA) in an ice bath. After that, the centrifugation of the homogenate was done at 12,000×g for 15 min, the 0.5 ml of the extract supplemented to 0.5 ml of 10 mM potassium phosphate buffer (pH 7.0) and 1 ml of 1 M KI. Finally, we calculated the H2O2 content via a standard curve after measuring the absorbance of the supernatant at 390 nm.
Lipid peroxidation
Lipid peroxidation was investigated through the determination of MDA [31]. In short we extracted 1 g callus tissue in 80% ethanol. The determination of MDA was based on the reaction with thiobarbituric acid (TBA) and the absorbance was measured at λ=440 nm, 532 nm and 600 nm by spectrophotometer (InfiniteM200 TECAN Group Ltd., Switzerland). Malondialdehyde (MDA) equivalents were estimated according to [31].
Protein extraction and quantification and enzyme assays
For protein and enzyme extractions, Fresh callus (1 g) was homogenized with 50 mM potassium phosphate buffer (pH 7.8) containing 1 mM EDTA-2Na and 7 % (w/v) polyvinylpolypyrrolidone (PVPP). The whole extraction procedure was carried out at 4°C. Centrifugation of the homogenates was performed at 4 °C for 15 minutes at 13,000g, and enzyme activity was measured using the supernatants. Protein was quantified as described by [32], utilizing bovine serum albumin as a standard.
Superoxide dismutase (SOD; EC 1.15.1.1) activity was determined according to [33] through the measuring of inhibition in the photochemical reduction of nitro blue tetrazolium (NBT) spectrophotometrically at 560 nm. The reaction mixture contained 50 mM potassium phosphate buffer (pH 7.8) with 0.1 mM ethylene diamine tetra acetic acid (EDTA), 75 μM NBT, 13 mM methionine, 2 μM riboflavin and 20 μl of protein extract. Reactions were performed for 10 min under a light intensity of 300 μmol−1 m−2s−1. The non-irradiated reaction mixture served as control and was deducted from absorption at 560 nm. One unit of SOD was defined as the amount of enzyme, which caused 50% inhibition of NBT reduction under the assay condition. The results were expressed as U mg −1 protein.
According to [14], we quantified the guaiacol peroxidase (GPX) activity (EC 1.11.1.7). The reaction solution includes 100 µl of plant extract supplemented by 700 µl of 0.05 M phosphate buffer (pH 7.8) and 200 µl of guaiacol (25 mM). The reaction begins by adding 100 µl of H2O2. The absorbance elevation generated by oxidation of guaiacol to tetra guaiacol was recorded for 3 min at 436 nm. POD activity was estimated from the extinction coefficient, Ɛ = 25.5 mM-1 cm-1. Results were expressed as µmol oxidized guaiacol min-1 mg-1 proteins
Ascorbate peroxidase (APX) activity (EC 1.11.1.11) was determined according to [34]. The reaction mixture contained 50 mM of potassium phosphate buffer (pH 7.0), 4.4 µl ascorbate (1 mM) and 10 µl EDTA-2Na (0.5 M). Adding H2O2 started the reaction and ascorbate oxidation was determined at 290 nm for 1 min. Activity was quantified using the extinction coefficient, e = 2.8 mM-1 cm-1. Each sample was measured in three repetitions. Results were expressed as µmol oxidized ascorbate min-1 mg-1 proteins.
According to [35], we assayed Catalase (CAT) activity (EC 1.11.1.6) by determination of the level of decomposition of H2O2 (Ɛ = 2.3 mM-1 cm-1) at 240 nm. This activity was measured in a reaction mixture containing 1,900 µl of potassium phosphate buffer (50 mM, pH 7.0 not containing EDTA), 100 µl sample and 1,000 µl H2O2 (30 mM). CAT activity is expressed as µmol H2O2 decomposed min-1 mg-1 proteins.
Statistical analysis
A completely random design was used in achieving all analyses. Significant differences between the treatments or varieties were detected via the software of SPSS Statistics 21 after subjecting all collected data to a one-way analysis of variance (ANOVA). Means comparison was made using the Tukey’s multiple range test (P=0.05). Mean values are presented with the standard errors.
Results
![]()
Growth and water relation
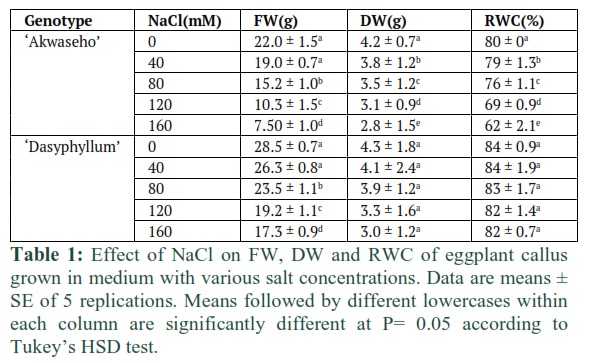
To investigate the impact of the rising salt concentration on the callus growth, leaf explants callus was initiated from 40 days old seedlings of S. macrocarpon ‘Akwaseho’ cv and S. dasyphyllum var dasyphyllum. Within 10 days we produced about 90% of callus from leaf explants of both species on solid MS basal medium containing 0.4 µM TDZ. The initial callus derived calli were maintained on MS medium added with 0.4 μM TDZ, 3% sucrose, and 0.8% agar–agar, pH 5.8. The developed calli were moved to the same solid MS medium supplemented with NaCl in five concentrations: 0 (Control), 40, 80, 120, or 160 mM NaCl. The increase of salinity contributes to a progressive diminution in callus growth in the two lines. Callus fresh weight was reduced, when compared to control, by 13.6 % to 65.9% in S. macrocarpon ‘Akwaseho’ cv and by 7.7% to 39.2% in S. dasyphyllum var dasyphyllum at 40 mM to 160 mM NaCl, respectively (Table 1). Callus cultured on control medium showed good cell proliferation in both lines. Up to 80 mM NaCl callus tissue occurred morphologically similar to control callus with no phytotoxicity symptoms in S. dasyphyllum var dasyphyllum; whereas a low cell proliferation and great portions displaying a brownish colour indicating cell necrosis were found in S. macrocarpon ‘Akwaseho’ cv grown under salt stress. When exposed to increasing NaCl levels, and compared to control, callus dry weight was reduced by 9.5% and 33.3% in S. macrocarpon ‘Akwaseho’ cv and by 4.6% and 30.2% in S. dasyphyllum var dasyphyllum at 40 mM to 160 mM NaCl, respectively (Table 1). Furthermore, RWC was significantly affected in calli of S. macrocarpon ‘Akwaseho’ cv by increasing salt concentration. Indeed, salinity contributes to the decline of the RWC from 80% at 0 mM NaCl medium to 62% at 160 mM NaCl medium. In contrast, RWC was hardly affected by salt stress in calli of S. dasyphyllum var dasyphyllum and no major effect was shown (Table 1).
Proline content
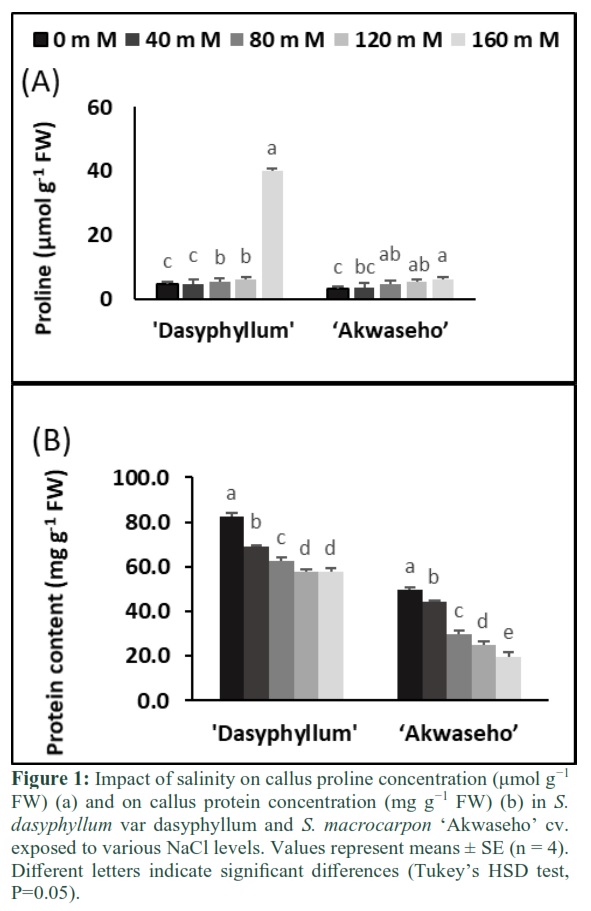
Rising NaCl concentration boosted up callus proline in both species. In S. dasyphyllum var dasyphyllum, a strong increase of callus proline was closely associated to the severity of the salinity. At 160 mM NaCl concentration callus proline increased was 10 times compared to the controls. While in S. macrocarpon ‘Akwaseho’ cv, there is a moderate callus proline increase of 1.2-fold which was noticed (Figure 1A).
Total protein accumulation
Salt stress reduced significantly the total protein content in both species. In S. macrocarpon ‘Akwaseho’ cv, the total soluble protein decrease was more drastic than in S. dasyphyllum var dasyphyllum, for all treatments. Increasing salinity concentration leads to less aggravated reduction in S. dasyphyllum var dasyphyllum, which maintained a significant higher soluble protein content. The highest salt level (160 mM NaCl) reduced protein accumulation by 30% and 60% respectively in S. dasyphyllum var dasyphyllum and S. macrocarpon ‘Akwaseho’ cv (Figure 1B).
Na+ and K+ concentration
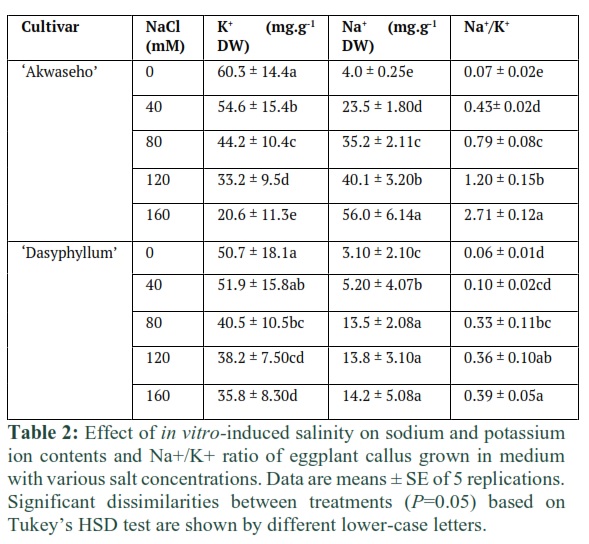
Our results pointed out that boosting up salinity content increased the Na+ accumulation in both species callus tissues. However, S. macrocarpon ‘Akwaseho’ cv exhibited more pronounced rising of Na+ content than S. dasyphyllum var dasyphyllum. The highest concentration of K+ was recorded from the control calli in S. macrocarpon ‘Akwaseho’ cv (60.3 mg.g-1 DW) and from 40 mM NaCl concentration in S. dasyphyllum var dasyphyllum (51.9 mg.g-1 DW). Whereas the lowest content of K+ were found at the 160 mM NaCl level in S. macrocarpon ‘Akwaseho’ cv (20.6 mg.g-1 DW) and in S. dasyphyllum var dasyphyllum (35.8 mg.g-1 DW) (Table 2). Na+/K+ ratio rose progressively with increasing salinity level in both genotypes. There was a genotype effect on this cation ratio contributing to higher callus Na+/K+ in S. macrocarpon ‘Akwaseho’ cv than in S. dasyphyllum var dasyphyllum (Table 2).
Lipid peroxidation and H2O2 accumulation
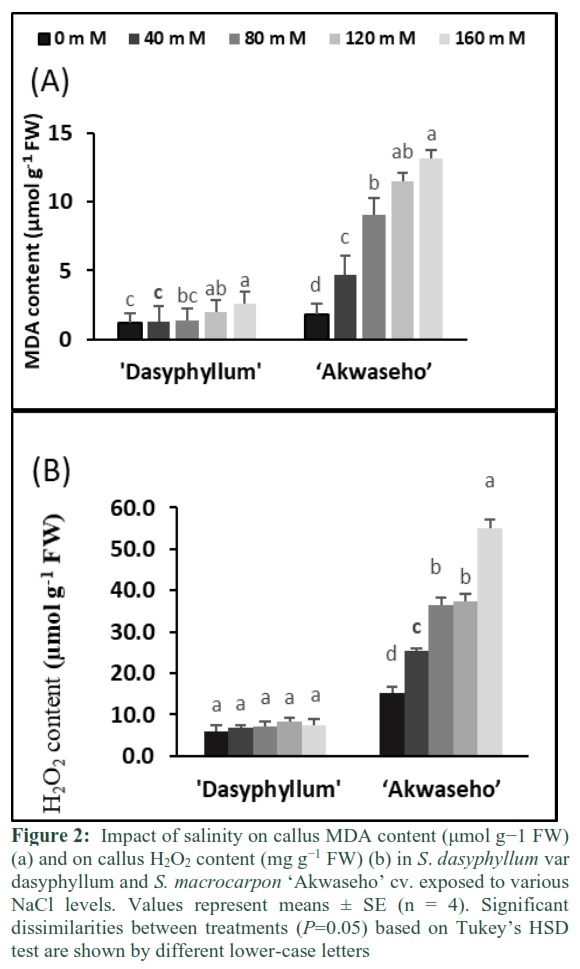
MDA content rose significantly with increasing salinity concentration in both species. S. macrocarpon ‘Akwaseho’ cv has shown a substantial augmentation at 40 mM NaCl while in S. dasyphyllum var dasyphyllum a moderate increase had only been noticed at 80 mM NaCl. Callus MDA content was enhanced respectively, 6-fold and 2-fold in S. macrocarpon ‘Akwaseho’ cv and in S. dasyphyllum var dasyphyllum at 160 mM NaCl, when compared to control (Figure 2A). The ROS scavenging machinery of callus tissues grown in salinity conditions was investigated through the determination of H2O2 content. Under salt stress, S. macrocarpon ‘Akwaseho’ cv clearly showed higher H2O2 content than S. dasyphyllum var dasyphyllum for all salt concentrations. H2O2 content was substantially increased in S. macrocarpon ‘Akwaseho’ cv and was 360% higher at 160 mM NaCl relative to the control. However, in S. dasyphyllum var dasyphyllum the callus H2O2 content increase was less (25% higher compared to control at the same salinity level) and no significant change was noticed (Figure 2B).
Antioxidant enzyme activity
Increasing salt concentration resulted in a significant reduction of the callus SOD enzyme activity in S. macrocarpon ‘Akwaseho’. This reduction ranged from 37% at 40 mM NaCl to 77.2% at 160 mM NaCl compared to the control. In S. dasyphyllum var dasyphyllum, 40 mM NaCl caused a non-significant decrease of the callus SOD enzyme activity. In contrast, rising salinity levels to 120 and 160 mM NaCl contributed significantly in enhancing the callus SOD enzyme activity by respectively 24% and 37.5% when compared to the control (Figure 3A). The two species showed two contrasting CAT activity under salt stress. The increase of salinity concentration to 80 mM NaCl and 160 mM NaCl caused callus CAT activity to be significantly elevated by 10.3% and 34.8%, respectively in S. dasyphyllum var dasyphyllum relative to control. Contrariwise, rising salinity level to 80 mM NaCl and 160 mM NaCl contributed to the reduction of callus CAT activity by 30.8% and 45.7%, respectively in S. macrocarpon ‘Akwaseho’ compared to control. It is noteworthy that S. dasyphyllum var dasyphyllum displayed higher callus catalase activity for all NaCl concentrations (Figure 3B). Increasing salinity level to 120 mM NaCl and 160 mM NaCl engendered the reduction of callus GPX activity in S. macrocarpon ‘Akwaseho’ cv by 46.4% and 61.9% relative to control. In contrast, rising salinity concentration to 120 mM NaCl and 160 mM NaCl enhanced callus GPX activity in S. dasyphyllum var dasyphyllum by 32.3% and 45.4, respectively when compared to control (Figure 3C). In S. dasyphyllum var dasyphyllum, salt stress enhanced callus APX activity from 80 mM NaCl to 160 mM NaCl. The rise ranged from 15% in 80 mM NaCl, 41.5% in 120 mM NaCl to 80% in 160 mM NaCl compared to control. However, callus APX activity declined in S. macrocarpon ‘Akwaseho’ cv under saline condition. For instance, the reduction ranged from 36% in 40 mM NaCl, to 54.5% in 160 mM NaCl, compared to control (Figure 3D).
Figures & Tables
Discussion![]()
Salt tolerance involves the development of several physiological and biochemical mechanisms by plant cell tissue in order to cope with salinity. Many authors emphasized the role of plant tissue culture as a promising and useful technique leading to the establishment and selection of salt tolerant somaclones or lines [5,13,36]. Growth decline under salinity conditions has been reported in several plant species grown in vitro [4,37]. In our experiment, callus growth parameters were adversely affected by increasing salt stress. However, S. dasyphyllum var dasyphyllum was more performant than S. macrocarpon ‘Akwaseho’ cv by showing less decrease in term of callus FW and DW (Table 1) and maintaining a quite stable callus RWC (Table 1) under stressful environment. The growth reduction might be due to the generated dehydration [36] combined to high toxic ion uptake such as Na+ and Cl−. According to the previous work [9], growth decrease appears in two specific steps: the osmotic stress fast reaction pursued by slow reaction induced by Na influx in cell tissue. Our results showed the capacity of S. dasyphyllum var dasyphyllum callus tissue in maintaining enhanced turgor and cell expansion using an effecient plasmolytic mechanism in order to overcome salt stress [4, 5, 38]. According to [38] proline accumulation in salt-tolerant tissues is more enhanced than in salt-sensitive ones. In S. dasyphyllum var dasyphyllum, callus accumulated higher proline amount than in S. macrocarpon ‘Akwaseho’ cv depending on NaCl level (Figure 1A). Consequently, the osmoregulation mechanism via proline synthesis was more effective to provide S. dasyphyllum var dasyphyllum better ability to adapt and tolerate salinity. In contrast, this osmoregulation process was inefficient in S. macrocarpon ‘Akwaseho’ cv. Plant adaptation to salinity is closely related to the osmoregulation efficiency. It is generally accepted that the majority of salt tolerant plants are provided with an effective osmoregulation system through a high capacity of accumulating organic or inorganic osmolytes, for example proline [ 39, 40]. In the present work callus of S. dasyphyllum var dasyphyllum displayed the lowest Na+ associated to the highest K+ content and consequently the lowest Na+/K+ ratio (Table 2). Thus contributing to higher FW, DW and RWC in all treatments (Table 1). This salt tolerance behavior could be explained by the fact that S. dasyphyllum var dasyphyllum developed a successful osmotic adjustment through the accumulation of K+ in the cells and maintained stable callus growth and water status and consumed less energy in producing and accumulating organic osmolytes. Similar findings has been reported earlier by [4, 38].
Interestingly, a low NaCl level was combined with slight elevated nutritional uptake of both Na+ and K+ by callus tissues in S. dasyphyllum var dasyphyllum. It is evident that low salt stress level provided a more optimal environment for cellular functions of eggplant callus tissue, as previously found with other species [41]. In contrast, S. macrocarpon ‘Akwaseho’ cv exhibited the highest Na+ associated with lowest K+ concentration and Na+/K+ ratio which generated lower growth rate (FW and DW) and RWC in all salinity levels. This may be caused by the high toxic ions accumulation in the cytoplasm and the depletion of elevated energy level in accumulating these ions in the vacuole [4, 16].
Marcum et al., [42] emphasized that the high Na+/K+ ratio engenders cytoplasmic enzymatic processes disruption. Under salinity condition, salt-tolerant callus tissue kept low cytosolic Na+ with elevated cytosolic K+ (via the extrusion and/or intracellular compartmentalization and consequently higher K+/Na+ ratio than salt sensitive callus tissue [5, 43].
Salt stress reduced significantly the total protein content in both species. In S. macrocarpon ‘Akwaseho’ cv, the total soluble protein decrease was more pronounced than in S. dasyphyllum var dasyphyllum, for all treatments (Figure 1B). The decline in total protein accumulation under stressful environment is in coherence with previous findings [36, 38]. [44] emphasized that the reduced protein level in plant subjected to salt stress might be due to a down regulation of protein synthesis via a rising protein hydrolyzing enzymes activities. The examination of lipid peroxidation level constitutes an effective tool to evaluate the oxidative damage in response to salt stress [45]. Callus of S. macrocarpon ‘Akwaseho’ cv had a relative high level of MDA and H2O2 content at 160 mM NaCl. For callus of S. dasyphyllum var dasyphyllum, less lipid peroxidation and oxidative stress were generated in callus tissue (Figure 3) and this should make less oxidative damage. This pointed out that S. dasyphyllum var dasyphyllum tissue showed better ability to cope with salt induced oxidative damage. This tolerance could be due to a stronger antioxidant enzymes machinery in S. dasyphyllum var dasyphyllum callus tissue (Figure 3). Our findings are in coherence with the results of earlier studies conducted on callus [16, 38].
Increasing salinity level caused SOD activity to be significantly declined in in S. macrocarpon ‘Akwaseho’ cv and substantially increased in S. dasyphyllum var dasyphyllum. Consequently, S. dasyphyllum var dasyphyllum showed a more performant scavenging system than S. macrocarpon ‘Akwaseho’ cv. Our results are in line with several previous reports [16,17] stating that salt tolerant species showed higher capacity in enhancing SOD activity than salt susceptible species.
Pessarakli et al., [44] emphasized that intracellular H2O2 levels is mostly regulated by CAT, GPX, APX, and GR under saline conditions. In the present work, the aggravated CAT activity reduction in S. macrocarpon ‘Akwaseho’ cv is closely linked to high H2O2 accumulation, while the CAT activity enhancement in S. dasyphyllum var dasyphyllum is related to low H2O2 level (Figure 2B and Figure 3B). This is in agreement with previous results reported by [46].
GPX activity followed the same trend as CAT activity. For instance, under saline condition GPX activity significantly rose in S. dasyphyllum var dasyphyllum and decreased in S. macrocarpon ‘Akwaseho’ cv (Figure 3C). This could be explained by higher capacity of decomposing H2O2 in S. dasyphyllum var dasyphyllum callus tissue than in S. macrocarpon ‘Akwaseho’ cv. This is in coherence with previous studies reported by [43].
Similar to CAT and GPX activity, rising salt stress declined APX activity in S. macrocarpon ‘Akwaseho’ cv callus tissue and increased it in S. dasyphyllum var dasyphyllum callus tissue (Figure 3D). As emphasized by [47] APX has an effective contribution to the regulation of the plant intracellular H2O2 level. It is evident that the combined enhancement of SOD and APX activity in S. dasyphyllum var dasyphyllum callus tissue efficiently led to overcoming oxidative stress. Our finding is in concordance with earlier results reported by [48] who stated that rising APX activity seems to be an adaptive mechanism giving the best regulation of H2O2 induced by a high SOD activity under saline conditions.
S. dasyphyllum var dasyphyllum callus tissue (seems more tolerant) was capable of exploiting its antioxidant enzyme machinery more actively than S. macrocarpon ‘Akwaseho’ cv callus tissue (seems more susceptible). Depending on the genotype (more tolerant or more susceptible), several differences have been found in levels of expression and activity of antioxidative enzymes [9]. The antioxidative enzymes activity was found to be up regulated during salt stress in many plants like chickpea [49], Cicer arietinum [50], melon [51], rice [52], rice [16] and tomato [53].
In conclusion, salt stress affects differentially the antioxidant enzyme machinery in the two studied species. S. dasyphyllum var dasyphyllum callus tissue occurs with higher ROS scavenging system, enhanced K+ content, low H2O2, and MDA accumulation, thus highlighting its better ability to tolerate salt stress than S. macrocarpon ‘Akwaseho’ cv which seems to be less protected and more susceptible to salt stress. The present work indicates that callus tissue offers a useful tool for future strategies to select salt-tolerant somaclonal variants in eggplant.
Author Contributions
Sami Hannachi: designed and performed experiments discussed the results and wrote the paper.
Insaf Bahrini: designed and performed experiments discussed the results and wrote the paper.
Nawaf I. Ibrahim: revised the manuscript. Abdelmuhsin Abdelgadir: revised the manuscript. Hira Affan Siddiqui: corrected the manuscript.
Acknowledgment
This research has been funded by Scientific Research Deanship at University of Hail, Saudi Arabia through project number RG-191346.
The authors declare that they have no competing interests.
References
- Mohamed Elhag. Evaluation of Different Soil Salinity Mapping Using Remote Sensing Techniques in Arid Ecosystems, Saudi Arabia, Journal of Sensors, (2016); 2016: 8.
- Munns R, Gilliham M. Salinity tolerance of crops—what is the cost? New Phytoligist, (2015); 208:668–673.
- Forni C, Duca D, Glick BR. Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil, (2017); 410: 335–356
- Golkar P., Amooshahi, F., Arzani, A. The effects of salt stress on physio-biochemical traits, total phenolic and mucilage content of Plantago ovata Forsk under in vitro conditions. Journal of Applied Botany and Food Quality, (2017); 90, 224 – 231.
- Al‑Khateeb AA, Sattar MN, Mohmand AS. Induced in vitro adaptation for salt tolerance in date palm (Phoenix dactylifera L.) cultivar Khalas. Biological Research, (2020); 53: 37.
- Aghaleh M, Niknam, V, Ebrahimzadeh, H, Razavi, K. Salt stress effects on growth, pigments, proteins and lipid peroxidation in Salicornia persica and S. europaea. Biologia Plantarum, (2009); 53: 243–248.
- Nikam AA, Devarumath RM, Shitole MG. Gamma radiation, in vitro selection for salt (NaCl) tolerance, and characterization of mutants in sugarcane (Saccharum officinarum L.). In Vitro Cellular & Developmental Biology – Plant, (2014); 50: 766–776.
- Dita MA, Rispai N, Prats E, Rubiales D, Singh KB. Biotechnology approaches to overcome biotic and abiotic stress constraints in legumes. Euphytica, (2006); 147: 1–24.
- Munns R, Tester M. Mechanisms of salinity tolerance. Annual Review of Plant Biology, (2008); 59: 651–681.
- Koc NK, Bas B, Koc M, Kusek M. Investigations of in vitro selection for salt tolerant lines in sour orange (Citrus aurantium L.). Biotechnology, (2009); 8: 155–159.
- Rout GR, Senapati SK, Panda JJ. Selection of salt tolerant plants of Nicotiana tabacum L. through invitro and its biochemical characterization. Acta Biologia Hung, (2008); 59: 77–92.
- Vaidyanathan H, Sivakumar P, Chakrabarty R, Thomas G. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.) differential response in salt-tolerant and sensitive varieties. Plant Science, (2003); 165: 1411–1418.
- Zhang F, Yang YL, He WL, Zhao X, Zhang X. Effects of salinity on growth and compatible solutes of callus induced from Populus euphratica. In Vitro Cell Development Biology-Plant, (2004); 40: 491–494.
- Chance B, Maehly AC. Assay of catalases and peroxidases. Methods of Biochemical Analysis, (1955); 2:764–775.
- Locato V, De Gara L. Plant programmed cell death: an overview. Methods in Molecular Biology, (2018); 1743: 1–8.
- Formentin E, Sudiro C, Ronci MB, Locato V, Barizza E, Stevanato P, Ijaz B, Zottini M, De Gara L, Lo Schiavo F. H2O2 Signature and Innate Antioxidative Profile Make the Difference Between Sensitivity and Tolerance to Salt in Rice Cells. Front Plant Science, (2018); 9:1549.
- Azevedo Neto AD, Prisco JT, Eneas-Filho J, de Abreu CEB, Gomes-Filho E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salttolerant and salt-sensitive maize genotypes. Environmental and Experimental Botany, (2006)5: 87–94.
- Kusvuran S, Ellialtioglu S, Yasar F, Abak K. Effects of salt stress on ıon accumulations and some of the antioxidant enzymes activities in melon (Cucumis melo L.), International Journal of Food and Agriculture, (2007); 2(5): 351-354.
- Sudhakar C, Lakshmi A, Giridarakumar S. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant Science, (2001); 161:613–619.
- Ünlükara A, Kurunç A, Kesmez GD, Yurtseven E, Suarez DL. Effects of Salinity on Eggplant (Solanum Melongena L.) Growth and Evapotranspiration. Journal of Irrigation and Drainage Engineering, (2008); 59: 203-214
- Bukenya ZR, Hall JB. Six cultivars of Solanum macrocarpon (Solanaceae) in Ghana. Bothalia, (1987); 17: 91–95.
- Bukenya ZR, Carasco JF. Crossability and cytological studies in Solanum macrocarpon and Solanum linnaeanum (Solanaceae). Euphytica, (1985); 86: 5–13.
- Knapp S, Vorontsova MSA. revision of the “African non-spiny” clade of Solanum L. (Solanum sections Afrosolanum Bitter, Benderianum Bitter, Lemurisolanum Bitter, Lyciosolanum Bitter, Macronesiotes Bitter, and Quadrangulare Bitter: Solanaceae). PhytoKeys, (2016); 66: 1–142.
- Yasar F, Ellialtioglu S. Antioxidative Responses of Some Eggplant Genotypes to Salinity Stress YYU Journal of Agricultural Science, (2013); 23: 215-221
- Brenes M, Pérez J, González-Orenga S, Solana A, Boscaiu M, Prohens J, Plazas M, Fita A, Vicente O. Comparative Studies on the Physiological and Biochemical Responses to Salt Stress of Eggplant (Solanum melongena) and Its Rootstock S. torvum. Agriculture, (2020); 10: 328.
- Murshige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Plant Physioly, (1962); 15:473-97.
- Sairam RK, Srivastava GC. Changes in antioxidant activity in subcellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Science, (2002); 162: 897–904.
- Bates LS, Waldran RP, Teare ID. Rapid determination of free proline for water stress. Plant Soil, (1973); 39, 205-208.
- Skoog DA, West DM, Holler FJ, Crouch SR. Analytical Chemistry: An Introduction, New Age International PVT, UK, (1973); 594-631.
- Velikova V, Yordanov I, Edreva A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: protective role of exogenous polyamines. Plant Science, (2000); 151: 59–66
- Hodges DM, Delong JM, Forney FC, Prange RK. Improving the thiobarbituric acidreactive- substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta, (1999); 207:604– 611.
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, (1976); 72: 248–254.
- Beauchamp C, Fridovich I. Superoxide dismutase improved assays and an assay applicable to acrylamide gels. Analytical Biochemistry, (1971); 44:276–287.
- Nakano Y, Asada K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiology, (1981); 22:867–880
- Aebi H. Catalases. In: Bergmeyer H. U. (ed) Methods of enzymatic analysis, vol. 2. Academic Press, New York, (1974); 673–684.
- Abed Alrahman NM, Shibli RA, Ereifej KI, Hindiyeh MY. Influence of salinity on growth and physiology of in vitro grown cucumber (Cucumis sativus L.). Jordanian Journal of Agricultural Science, (2005); 1: 93–106.
- Sotiropoulos TE, Dimassi KN, Tsirakoglou V, Therios IN. Response of two Prunus rootstocks to KCl induced salinity in vitro. Biologia Plantarum, (2006); 50: 477-480.
- Ibrahim MA, Jerry AN, Khalil AI. Effect of salt stress on some chemicals characteristics of callus for three cultivars of potato plant (Solanum tuberosum L.). AAB Bioflux, (2018); 10(2):87-96
- Rejeb KB, Abdelly C, Savouré A. How reactive oxygen species and proline face stress together. Plant Physiology and Biochemistry, (2014); 80: 278–284.
- Santangeli M, Capo C, Beninati S, Pietrini F, Forni C. Gradual Exposure to Salinity Improves Tolerance to Salt Stress in Rapeseed (Brassica napus L.). Water, (2019); 11: 1667.
- Beloualy N, Bouharmont J. NaCl-tolerant plants of Poncirus trifoliata regenerated from tolerant cell lines. Theoretical and Applied Genetics, (1992); 83: 509–514
- Marcum KB, Yensen NP, Leake JE. Genotypic variation in salinity tolerance of Distichlis spicata turf ecotypes. Australian Journal of experimental Agriculture, (2007); 47(12): 1506-1511
- Ganie SA, Molla KA, Henry RJ, et al. Advances in understanding salt tolerance in rice. Theoretical and Applied Genetics, (2019); 132:851–70.
- Pessarakli M, Tucker TC. Dry Matter Yield and Nitrogen-15 Uptake by Tomatoes under Chloride Stress. Soil Fertility and Plant Nutrition, (1988); 52: 698-700.
- Masood A, Ahmad SN, Zeeshan M, Abraham G. Differential response of antioxidant enzymes to salinity stress in two varieties of Azolla (Azolla pinnata and Azolla filiculoides). Environmental and Experimental Botany, (2006); 58: 216–222.
- Sevengor S. Investigations on antioxydant enzyme activities under in vitro and in vivo conditions to obtain salt tolerance in squash (Cucurbita pepo L.). Ph.D.Thesis, Ankara Universitiy, Graduate School of Natural and Applied Sciences, Ankara, (2010); p. 17.
- Shigeoka S, Ishikawa T, Tamoi M, Miyagawa Y, Takeda T, Yabuta Y, Yoshimura K. Regulation and function of ascorbate peroxidase isoenzymes. Journal of Experimental Botany, (2002) ;53:1305–1319
- Lopez MV, Satti SME. Calcium and Potassium- Enhanced Growth and Yield of Tomato Under Sodium Chloride Stress. Plant Science, (1996); 114: 19-27.
- Kukreja S, Nandwal AS, Kumar N, Sharma SK, Unvi V, Sharma PK. Plant water status, H2O2 scavenging enzymes, ethylene evolution and membrane integrity of Cicer arietinum roots as affected by salinity. Biologia Plantarum, (2005); 49: 305–308.
- Eyidogan F, Öz MT. Effect of salinity on antioxidant responses of chickpea seedlings. Acta Physiologia Plantarum, (2007); 29: 485–493.
- Kusvuran S. Relationships between physiological mechanisms of tolerances to drought and salinity in melons. PhD Thesis, Department of Horticulture, Institute of Natural and Applied Sciences, University of Cukurova, Turkey, (2007); p.356
- Roychoudhury A, Basu S, Sengupta DN. Antioxidants and stress related metabolites in the seedlings of two indica rice varieties exposed to cadmium chloride toxicity. Acta Physiologia Plantarum. (2012c); 34: 835–847.
- Gapiñska M, Skłodowska M, Gabara B. Effect of short-and longterm salinity on the activities of antioxidative enzymes and lipid peroxidation in tomato roots. Acta Physiologia Plantarum, (2008); 30, 11–18.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


