

Full Length Research Article
Response of antioxidants and Reactive oxygen species at various exogenous PEG and proline levels in rice callus
Muhammad Ilyas Khokhar*, Imran Habib, Muhammad Jamshaid Anwar, Sajid ur Rehman
Adv. life sci., vol. 8, no. 4, pp. 374-380, December 2021
*– Corresponding Author: Muhammad Ilyas Khokhar (Email: ilyasabri@yahoo.com)
Authors' Affiliations
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Rice is a water loving crop and drought stress at any physiological stage can hamper the growth as well as its yield. Hence this research was conducted to assess the drought tolerance capability of elite basmati rice cultivars of Pakistan.
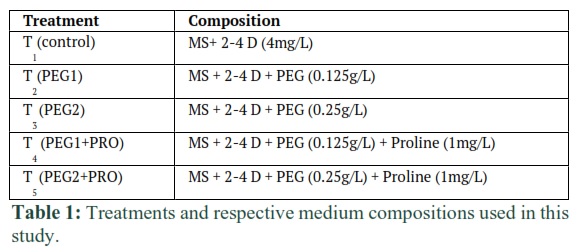
Methods: Polyethylene glycol (PEG) mediated in vitro experiment was conducted to estimate various water stress related parameters like callogenesis, crude protein, total antioxidant 2,2-Diphenyl-1-Picrylhydrazyl (DPPH), hydrogen peroxide (H2O2) and proline accumulation in callus. Dehusked seeds of five rice genotypes i.e. Basmati-2000, Super Basmati, Basmati-515, Basmati-385 and PS-2 were cultured for callogenesis on Murashige and Skoog (MS) medium having 2,4-Dichlorophenoxyacetic acid (2,4-D) @ 4mg/L, proline @ 1mg/L and two levels of PEG (0.125g/L and (0.25g/L).
Results: In case of relative performance under all treatments, maximum callus fresh weight (0.308g) was observed in PS-2 followed by Basmati-515 (0.281g) and minimum (0.198g) in Basmati-385 after 30 days of culturing. Maximum proline (72.29 μmol/g FW), crude protein (5.89%) and total antioxidant DPPH (70.77%) were noted in Basmati-2000 while maximum H2O2 (75.55 μmol/g FW) was observed in PS-2 and minimum (37.22 μmol/g FW) in Basmati-2000. It was observed that under both PEG treatments, the rice cultivar Basmati-2000 was found to be highly drought tolerant. Maximum callus fresh weight was observed in the treatment without the addition of PEG and proline. Maximum production of Proline, protein and DPPH were observed at a low level of PEG (0.125g/L) in presence of proline @ 1mg/L.
Conclusion: Results revealed that application of proline in rice may be effective in mitigating the effect of drought stress in the presence of PEG under in-vitro conditions.
Keywords: Basmati rice; Callogenesis; DPPH; Hydrogen peroxide; Protein
Introduction![]()
Pakistan ranked at 10th position in Asian rice producing countries [1]. Rice is the second important staple food after wheat and major exportable commodity after cotton in Pakistan. It added 3.1% value in agriculture and contributed 0.6% GDP to the national economy of Pakistan (2019-20). It is cultivated on an area of 3,034,000 ha with production 7410,000 tons and an average yield of 2442 kg/ha [2]. Water scarcity and inadequate soil moisture are major issues in rice growing areas. Drought severely affected paddy yield [3]. Drought is a major threat in the rainfed zone and reduced 13–35% rice yield [4]. Aromatic rice has high demand in the national and international market due to its fragrant aroma [4,5]. Rice quality and yield are highly linked with drought tolerance [6]. There is a dire need to give special attention to this issue. Basmati rice genotypes are late maturing cultivars (5-6 months) and required more irrigation water due to their higher evapotranspiration. The research will be focused to develop short duration and drought tolerant high yielding rice varieties. Screening against drought tolerant rice genotypes through conventional breeding is laborious and time consuming, however, in vitro method is mostly used as it is simple and effective [7]. In vitro culture produced callus and regeneration in fine and coarse rice [8]. The callus is an undifferentiated mass of cells developed when 2, 4-D alone or in combination with cytokinins [9]. An efficient callus induction is also a selection against abiotic stress tolerance when various PEG doses are added in the MS medium [10]. El-Tayeb and Hassanein; Nepomuceno et al. [11,12] noted that PEG sharply reduced tissue moisture, used for drought stress and tolerant cell lines selection. Different osmatic agents i.e., mannitol, PEG and sorbitol were added in the medium of explants for creating artificial drought stress [7] but PEG is mostly used in rice screening as it has a wide range of molecular weights, non-ionic water-soluble polymer and modify osmotic potential of culture medium [10]. Reactive oxygen species (superoxide, H2O2, hydroxyl radicals & singlet oxygen) lead to oxidative degradation of lipid and protein membrane, deactivation of enzymes which cause the death of plants under water stress [3,13,14]. Joshi et al. [14] reported somaclones of PEG treated rice calli had more proline, chlorophyll, number of tillers and plant height comparative to control.
The study is subjected to find the response of basmati/fine rice varieties under various PEG and exogenous proline treatments for its impact on callus fresh weight, hydrogen peroxide, proline, and protein contents. Moreover, this study will help to screen out rice calli for drought tolerance among selected genotypes under in vitro conditions. Best drought tolerance genotype calli will be used in another experiment for somaclones development using standardized regeneration protocol [8].
Methods![]()
This research was conducted at cytogenetic laboratory, Agricultural Biotechnology Research Institute (ABRI), Faisalabad, Pakistan from October 2019 to September 2020.
Callus induction medium
Callus medium contained phyta gel (1.76 g/L) (ID: G434), Murashige and Skoog medium (4.43g/L MS; 1962; ID: M519), sucrose (30g/L) 2, 4-dichlorophenoxy acetic acid (4mg/L, ID: D299). Exogenous PEG (0.125 and 0.25g/L) and proline (1mg/L) were also added alone and with combination in callogenesis medium. All treatments used in the present study are mentioned in table 1. The pH of these media was maintained at 5.7 by adding 1N HCl or 1N NaOH. Chemicals used in this study with code IDs of USA Phyto Technology Laboratories were the major products. The medium was then autoclaved at 15 psi for 15 min at 120°C.
Explant culturing
The required research material was collected from Rice Research Institute, Kala Shah Kaku. Paddy seeds dehusked manually, washed with distilled water to remove dust and other particles, surface sterilized with 35% Clorox for 15 min on a shaker (3020 model, GFL, Burgwedel, Germany) at 154 rpm followed by 70% alcohol for 30 seconds, then rinsed 3 times for 5 minutes in autoclaved distilled water. All procedures related to sterilization and culture was performed in a laminar flow cabinet in the tissue culture room of ABRI. The laminar flow bench was illuminated with UV-light for 20 min before each operation. Hands and all instruments were regularly disinfected with 70% ethanol. Explants (surface sterilized dehusked seeds) were cultured on callus induction medium. Seeds were cultured at one seed/test tube and incubated for 20 days in the dark at 25°C, as suggested by Noor et al. [15]. Thereafter, test tubes were transferred under light (16-h photoperiod with a photosynthetic photon flux density (PPFD) of 83.6 µmol m-2 s-1 provided by white fluorescent tubes) for 10 days and incubated at 25 ± 2°C, based on Hussain et al. [16].
Data Collection
Data was noted for various parameters from rice callus after 30 days of seed culture.
- Callus fresh weight
Callus fresh weight noted with weight balancing machine after 30 days.
- Crude protein test
Kjeldahl method with conversion factor (6.625×3) was applied to determine crude protein content.
- 2, 2, diphenyl-2-picryl-hydrazyl (DPPH)
2, 2, diphenyl-2-picryl-hydrazyl method developed by Turkmen, Sari and Velioglu [17] was applied with some modification to detect Free radical scavenging activity of the sample.
- Hydrogen peroxide (H2O2) test
Fresh callus (0.2g) was frozen in liquid nitrogen and grounded to a fine powder with sterilized pestle mortar and H2O2 in rice callus was measured with modified method described by Patterson et al. [18].
- Proline test
Fresh callus (0.2g) was used for estimation of proline contents with a modified method described by Bates et al. [19].
Statistical analysis
The experiment was laid out with 3 replicates in a factorial complete randomized design having 50 samples per replication. MSTAT-C software was applied for data analysis to calculate the least significant difference test @ α=0.05.
Results
![]()
Analysis of variance revealed that the effect of treatments, genotypes and treatments x genotypes showed highly significant results for callus fresh weight, proline, protein, DPPH and H2O2 whereas the non-significant effect was observed for protein in the case of treatments x genotypes.
Callus fresh weight (g/kg)
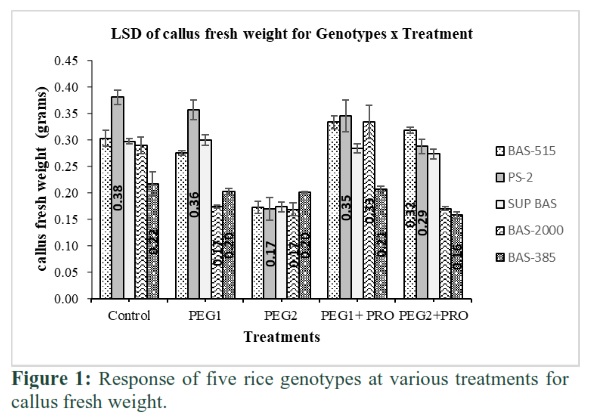
Maximum callus fresh weight (0.38g) was observed in PS-2 while minimum (0.22g) in Basmati-385 at treatment-1. Callus fresh weight was reduced in all genotypes with increase in PEG levels. When proline (1mg/L) was added in treatment 4 & 5, high response was noted in callus weight (0.35 & 0.28g/Kg) of PS-2 followed by Basmati-515 (0.33 & 0.32g/Kg) while low (0.21 and 0.16mg/Kg) in Basmati-385, detailed in Figure 1.
Protein
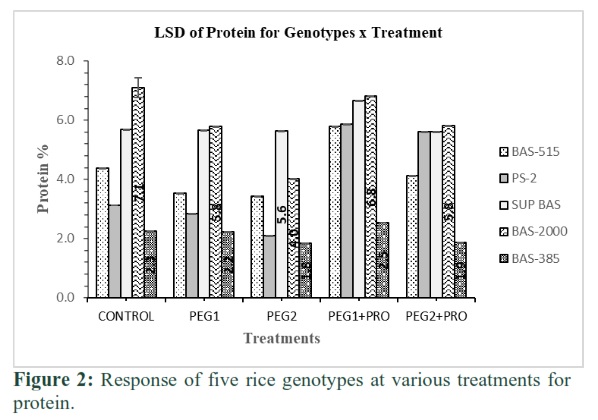
All genotypes behaved differently for protein percentage at various treatments as described in Figure 2. Overall, the maximum protein (7.1%) was found in Basmati-2000 at treatment-1 and minimum (1.9%) was found in Basmati-385 at treatment-5. Protein level considerably decreased in the callus of all studied genotypes under PEG treatment. Basmati-2000 and PS-2 performed at par at treatment-2 and lowest protein (2.2%) was noted in Basmati-385. PS-2 surpassed all others with maximum protein (5.6%) followed by Basmati-2000 (4%) at treatment-3. Protein levels were found at par in Basmati-2000 and PS-2 at treatments 4 and 5.
Proline
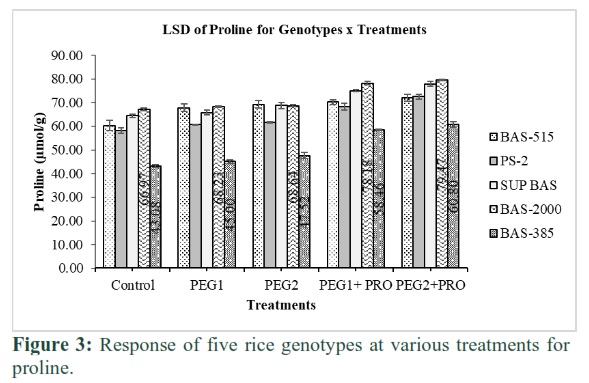
Figure 3 depicted various proline levels in the callus of rice genotypes. Maximum proline (67μmol/g) was noted in Basmati-2000 and minimum (43.1μmol/g) in Basmati-385 at treatment-1. Proline level increased as stress level enhanced and depicted maximum (69.2μmol/g) in Basmati-515 and minimum (47.5μmol/g) at treatment-3. Maximum callus proline (78.2μmol/g) and (79.5μmol/g) noted in Basmati-2000 at treatment 4 & 5 which observed at par Basmati-515.
2, 2-Diphenyl-1-picrylhydrazyl (DPPH)
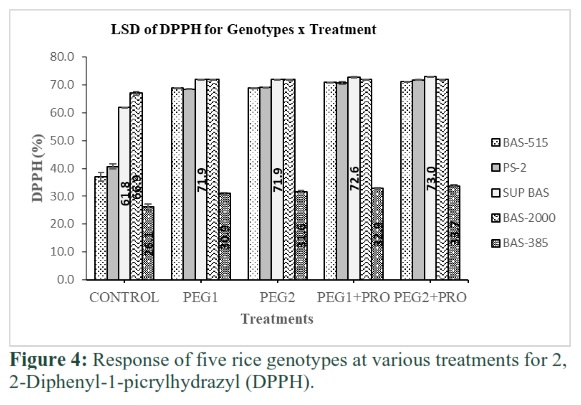
Five rice genotypes responded differently for DPPH at the control level. DPPH value (66.88%) observed maximum in Basmati-2000 and minimum (26%) for Basmati-385 at treatment-1. Basmati-515, PS-2, Super Basmati and Basmati-2000 behaved at par for DPPH at treatment-2 & 3 and minimum (31.6 and 30.9) in Basmati-385. DPPH value was enhanced in presence of proline at treatment-4 and 5. Maximum values (72.96% and 72.68%) were observed in Super Basmati at treatment-4 and 5, respectively.
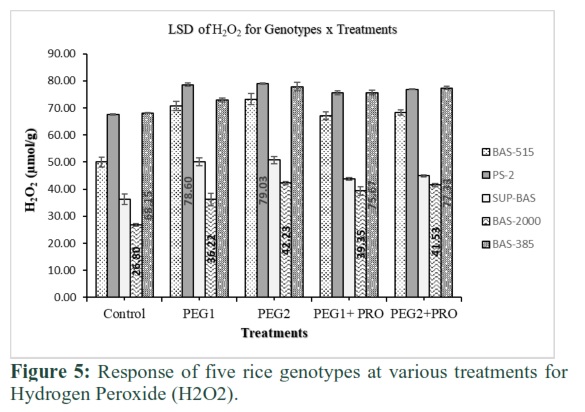
Hydrogen peroxide (H2O2)
H2O2 produced maximum in callus of all genotypes in PEG medium as revealed in Figure 5. The maximum quantity of H2O2 (79 μmol/g) was produced at PEG treatment-2 in PS-2 and 78.6μmol/g in PEG treatment-1. The minimum quantity of H2O2 was noted in Basmati-2000 at PEG treatment-1. Basmati-2000 produced 36.2 μmol/g and 42.2 μmol/g H2O2 when proline was added in PEG medium. H2O2 found maximum in Basmati-385 at treatment-4 and 5, respectively. Minimum H2O2 value was recovered in Basmati-2000 at treatment-4 and 5 presence of proline in PEG medium.
Figures & Tables
Discussion![]()
Joshi et al. [14] studied high PEG levels in medium reduced water contents, rice callus growth and volume which corroborates with our study. Callus fresh weight decreased in all genotypes in our study when PEG levels gradually increased. Al-Bahrany [20] exposed rice calli “Hassawi” at various PEG-8000 levels (0, 50, 100, 150, 200, 250 and 300g/L) and observed progressive reduction in callus fresh weight and water content whereas 200 g/L was found to have the highest inhibitory impact. In the present study, the increasing PEG levels also resulted in a decline in callus fresh weight especially in treatment-3 indicating a certain threshold level of varietal resistance to external PEG application. Zain et al. [21] exposed callus of aromatic rice variety “MRQ74” to radiation and later supplemented MS media with various PEG-6000 levels (1.5, 3.0, 4.5, 6.0 and 7.0%) for drought stress and observed maximum callus fresh weight linked with increased proline contents in drought resistant rice calli. The highest callus weight was observed in PS-2 in the present study, which may be a direct result of elevated proline concentration at cellular levels.
This study depicted that protein %age considerably declined under PEG treatments, and it was also genotype dependent. Genotype performed better under PEG treatment and may also be performed well in proline + PEG treatments. Performance of Basmati-2000 and PS-2 found best under all treatments compared to other genotypes. Under abiotic stress, various protective mechanisms are activated in plant cells. Some of these defensive substances have low molecular weight like osmolites and certain stress induced proteins i.e. heat shock proteins, aquaporins and late embryogenesis abundance proteins (LEAs) [22]. In the current study, despite the accumulation of protein dependent release of osmolites, under various treatments of PEG induced osmotic stress, overall protein contents of calli were reduced. However, considerable variation in protein contents was observed among studied rice genotypes at different PEG levels. This demonstrates that protein contents in plants under abiotic stress may be related to genotypes infrastructure as well as varying concentrations of PEG. Considerably lower protein contents of the cell may also be indicative of higher level of cellular catabolism as compared to anabolism under drought stress [23]. Similar to these findings, Hameed et al. [24] also observed reduced cell growth and protein contents in wheat under salinity stress.
This study presented proline content exceeded when externally added in medium. Proline level in callus is also genotype dependent and maximum proline observed under high PEG applications. Fine rice genotype (PS-2) performed better compared to basmati genotype under stress conditions. High proline levels may also be caused by better callus growth. Reduction of growth and higher cellular proline contents are the main responses of cells to various abiotic stresses under in-vitro callus culturing [25]. In rice, higher osmotic stress caused a gradual decline in calli growth and vigour. Proline acts as an osmolyte and is found extensively high in plant species which accumulates in great quantitates as compared to other amino acids in response to salinity or water deficit stresses [26]. Sometimes proline’s role in drought alleviation is marked as selection criteria for water deficit stress [27]. Proline released in cells is considered to be a mechanism of osmotic regulation and adjusting of rice calli water by osmolytic effects of proline under water stress [28]. Hence in the present study, significantly higher accumulation of proline in callus of rice variety Basmati-2000 may be an indication of adaptive survival mechanism to protect the precious cellular water contents by lowering the rate of cell growth [29].
DPPH values found less in Basmati-385 comparative to other genotypes in all treatments as presented in Figure 4. DPPH-free radical method is one of the most commonly used antioxidant detection assay that depends on the electron relocation and it generates violet colour when mixed in ethanol, this free radical is reduced by antioxidant molecules at stable ambient temperature and produced colourless solution [30]. In the present investigation, rice genotypes basmati-2000 and Super Basmati exhibited the highest DPPH activity under drought stress indicating the presence of efficient antioxidant and free radical scavenging activity in the calli of these varieties under PEG induced water limitation stress. Rahman et al. [31] also found a positive correlation between DPPH based free radical and reduced lipid peroxidation in Tabebuia pallida under a stress environment. Radical inhibition percentages of DPPH ranged from 66.9 -93.5% was observed in Gynura bicolor whereas the highest DPPH activity (84%) in calli and lowest scavenging was noted in the shoot of this plant [32].
All genotypes were affected under PEG stress medium as compared to the control condition. However, Basmati-2000 showed little effect as compared to other genotypes. Proline compensates the effect of PEG to produce H2O2 in callus of all rice genotypes. Reactive oxygen species are produced under abiotic stresses and their conversion into H2O2 caused cellular membrane peroxidation, protein aggregation, DNA denaturation or toxicity of various metabolic pathways in cell [33]. Various environmental stresses like drought, salinity, heat or chilling are known to stimulate H2O2 production in plants [34]. Excessive H2O2 in the cell is countered by releasing catalase (CAT) and ascorbate peroxidase (APX) which are responsible for the reduction or elimination of this injurious substance at a cellular level [35]. In the current study the highest level of H2O2 was released in PS-2 variety at both PEG treatment-1 and 2, indicating the drought susceptibility and inability to produce required levels of ROS scavengers under water limitation stress. In presently study interestingly on the addition of proline added to PEG treated media, Basmati-2000 managed to reduce H2O2 at less injurious levels. It indicated the role of proline as a compatible solute to adjust osmotic potential in the cell of stressed plants [36]. Similar exogenous proline supplementation was found to have a protective effect on maize seedlings under heat stress due to modulation of lipid peroxidation process [37].
Maximum callus fresh weight was observed in PS-2 and minimum in Basmati-385. Maximum proline, crude protein and total antioxidant DPPH were noted in Basmati-2000 while maximum H2O2 was observed in PS-2 and minimum in Basmati-2000. It is observed that under both PEG treatments, Basmati-2000 was found to be highly drought tolerant. Maximum callus fresh weight was observed in treatment without the addition of PEG and proline. Maximum production of proline and DPPH were observed at a low level of PEG in presence of proline, whereas maximum crude protein was observed at a higher PEG level. From this study, it is concluded that the application of proline may be effective in mitigating the effect of drought. Callus of Super Basmati and Basmati-2000 was found to be highly drought tolerant as it has high values of proline, DPPH and low production of H2O2 under stress treatments.
Author Contributions
Muhammad Ilyas khokhar planned, conducted the research work, collected the data and wrote-up the paper. Imran Habib assisted in the discussion portion. Muhammad Jamshaid Anwar helped in proof reading. Sajid ur Rehman supervised the experiment.
All authors declared no conflicts of interest.
Acknowledgment
All authors would like to express their gratitude to the Ayub Agricultural Research Institute, Faisalabad and its sub sections i.e., Soil Chemistry, Soil Biochemistry and STAT department for providing expertise and analysis facility required to conduct this research.
References![]()
- Shah MAA, Özel G, Chesneau C, Mohsin M, Jamal F, et al. A statistical study of the determinants of rice crop production in Pakistan. Pakistan Journal of Agricultural Research, (2020); 33(1): 97-105.
- Pakistan E, Survey, (2020) Agriculture, Pakistan Economic Survey 2020-21. Islamabad. 17-44 p.
- Akte J, Yasmin S, Bhuiyan M, Khatun F, Roy J, et al. In vitro screening of rice genotypes using polyethylene glycol under drought stress. Progressive agriculture, (2016); 27(2): 128-135.
- Lafitte H, Li Z, Vijayakumar C, Gao Y, Shi Y, et al. Improvement of rice drought tolerance through backcross breeding: evaluation of donors and selection in drought nurseries. Field Crops Research, (2006); 97(1): 77-86.
- Roychoudhury A, Basu S, Sarkar SN, Sengupta DN. Comparative physiological and molecular responses of a common aromatic indica rice cultivar to high salinity with non-aromatic indica rice cultivars. Plant cell reports, (2008); 27(8): 1395-1410.
- Degenkolbe T, Do PT, Zuther E, Repsilber D, Walther D, et al. Expression profiling of rice cultivars differing in their tolerance to long-term drought stress. Plant molecular biology, (2009); 69(1): 133-153.
- Kacem NS, Delporte F, Muhovski Y, Djekoun A, Watillon B. In vitro screening of durum wheat against water-stress mediated through polyethylene glycol. Journal of genetic engineering and biotechnology, (2017); 15(1): 239-247.
- Khokhar MI, Iqbal MZ, Teixeira da Silva J. Rapid callogenesis and plant regeneration of fine and coarse varieties of rice. BioTechnologia Journal of Biotechnology Computational Biology and Bionanotechnology, (2017); 98(4).
- Chakraborty A, Chowdhury S, Haque Z, Dash AK, Mosnaz A. In vitro shoot regeneration of rice (Oryza sativa L.) varieties under artificial drought stress. The Agriculturists, (2018); 16(1): 55-64.
- Wani SH, Sofi PA, Gosal SS, Singh NB. In vitro screening of rice (Oryza sativa L) callus for drought tolerance. Communications in Biometry and Crop Science, (2010); 5(2): 108-115.
- El-Tayeb M, Hassanein A. Germination, seedling growth, some organic solutes and peroxidase expression of different Vicia faba lines as influenced by water sterss. Acta agronomica hungarica, (2000); 48(1): 11-20.
- Nepomuceno A, Oosterhuis D, Stewart J. Physiological responses of cotton leaves and roots to water deficit induced by polyethylene glycol. Environmental and Experimental Botany, (1998); 40(1): 29-41.
- Phung T-H, Jung H-i, Park J-H, Kim J-G, Back K, et al. Porphyrin biosynthesis control under water stress: sustained porphyrin status correlates with drought tolerance in transgenic rice. Plant physiology, (2011); 157(4): 1746-1764.
- Joshi R, Shukla A, Sairam RK. In vitro screening of rice genotypes for drought tolerance using polyethylene glycol. Acta Physiologiae Plantarum, (2011); 33(6): 2209-2217.
- Noor A, Rashid H, Chaudhry Z, Mirza B. High frequency regeneration from scutellum derived calli of basmati rice cv. Basmati 385 and Super basmati. Pakistan Journal of Botany, (2005); 37(3): 673.
- Hussain Z, Khan MH, Bano R, Rashid H, Chaudhry Z. Protocol optimization for efficient callus induction and regeneration in three Pakistani rice cultivars. Pakistan Journal of Botany, (2010); 42(2): 879-887.
- Turkmen N, Sari F, Velioglu YS. The effect of cooking methods on total phenolics and antioxidant activity of selected green vegetables. Food chemistry, (2005); 93(4): 713-718.
- Patterson BD, MacRae EA, Ferguson IB. Estimation of hydrogen peroxide in plant extracts using titanium (IV). Analytical biochemistry, (1984); 139(2): 487-492.
- Bates LS, Waldren RP, Teare I. Rapid determination of free proline for water-stress studies. Plant and soil, (1973); 39(1): 205-207.
- Al-Bahrany AM. Callus growth and proline accumulation in response to polyethylene glycol-induced osmotic stress in rice, Oryza sativa L. Pakistan Journal of Biological Sciences (Pakistan), (2002).
- Che R, Kadhimi AA, Arshad NA, Anizan I, Azhar M, et al. Enhancing of drought-tolerant rice (Oryza sativa) variety MRQ74 through gamma radiation and in vitro pathway. Biotechnology, (2016); 15(6): 125-134.
- Büyük İ, Soydam-Aydın S, Aras S. Molecular responses of plants to stress conditions. Türk Hijyen ve Deneysel Biyoloji Dergisi, (2012); 69(2): 97-110.
- Dogan M. Effect of salt stress on in vitro organogenesis from nodal explant of Limnophila aromatica (Lamk.) Merr. and Bacopa monnieri (L.) Wettst. and their physio-morphological and biochemical responses. Physiology and Molecular Biology of Plants, (2020); 26(4): 803.
- Hameed A, Naseer S, Iqbal T, Syed H, Haq MA. Effects of NaCl salinity on seedling growth, senescence, catalase and protease activities in two wheat genotypes differing in salt tolerance. Pak J Bot, (2008); 40(3): 1043-1051.
- Al-Khayri JM. Growth, proline accumulation and ion content in sodium chloride-stressed callus of date palm. In Vitro Cellular & Developmental Biology-Plant, (2002); 38(1): 79-82.
- Niknam V, Razavi N, Ebrahimzadeh H, Sharifizadeh B. Effect of NaCl on biomass, protein and proline contents, and antioxidant enzymes in seedlings and calli of two Trigonella species. Biologia Plantarum, (2006); 50(4): 591-596.
- Ashraf M, Harris P. Potential biochemical indicators of salinity tolerance in plants. Plant science, (2004); 166(1): 3-16.
- Lokhande VH, Nikam TD, Penna S. Biochemical, physiological and growth changes in response to salinity in callus cultures of Sesuvium portulacastrum L. Plant Cell, Tissue and Organ Culture (PCTOC), (2010); 102(1): 17-25.
- Zhao X, Tan H, Liu Y, Li X, Chen G. Effect of salt stress on growth and osmotic regulation in Thellungiella and Arabidopsis callus. Plant Cell, Tissue and Organ Culture (PCTOC), (2009); 98(1): 97-103.
- Huang D, Ou B, Prior RL. The chemistry behind antioxidant capacity assays. Journal of agricultural and food chemistry, (2005); 53(6): 1841-1856.
- Rahman MM, Islam MB, Biswas M, Alam AK. In vitro antioxidant and free radical scavenging activity of different parts of Tabebuia pallida growing in Bangladesh. BMC research notes, (2015); 8(1): 1-9.
- Krishnan V, Ahmad S, Mahmood M. Antioxidant potential in different parts and callus of Gynura procumbens and different parts of Gynura bicolor. BioMed Research International, (2015); 2015.
- Sharma P, Jha AB, Dubey RS, Pessarakli M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. Journal of botany, (2012); 2012.
- Avsian-Kretchmer O, Gueta-Dahan Y, Lev-Yadun S, Gollop R, Ben-Hayyim G. The salt-stress signal transduction pathway that activates the gpx1 promoter is mediated by intracellular H2O2, different from the pathway induced by extracellular H2O2. Plant Physiology, (2004); 135(3): 1685-1696.
- Yang Y, Shi R, Wei X, Fan Q, An L. Effect of salinity on antioxidant enzymes in calli of the halophyte Nitraria tangutorum Bobr. Plant Cell, Tissue and Organ Culture (PCTOC), (2010); 102(3): 387-395.
- Bartels D, Sunkar R. Drought and salt tolerance in plants. Critical reviews in plant sciences, (2005); 24(1): 23-58.
- Ali Q, Anwar F, Ashraf M, Saari N, Perveen R. Ameliorating effects of exogenously applied proline on seed composition, seed oil quality and oil antioxidant activity of maize (Zea mays L.) under drought stress. International Journal of Molecular Sciences, (2013); 14(1): 818-835.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0