

Full Length Research Article
The Effect of Drought Stress on the Superoxide Dismutase and Chlorophyll Content in Durum Wheat Genotypes
Majid Khayatnezhad*, Roza Gholamin
Adv. life sci., vol. 8, no. 2, pp. 119-123, February 2021
*- Corresponding Author: Majid Khayatnezhad (Email: Khayatnezhad@gmail.com)
Authors' Affiliations
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Drought stress is one of the most limiting factors of plant production around the world. So, finding a way for increasing genotypes resistance is so important. Free radicles and other dynamic subordinates of oxygen inactivate chemicals and significant plant cell parts. Superoxide dismutases (SODs) have been distinguished as essential parts in a creature's guard system.
Methods: This examination was carried out to examine the SOD movement in 8 durum wheat genotypes from Iran and Azerbaijan under two different conditions in 2015-2016 cropping year. The impact of dry season weight on SOD, chlorophyll content list (CCI), and chlorophyll debasement were examined. Critical contrasts among genotypes and the genotype × climate collaboration among SOD and CCI content were distinguished.
Results: The mean examination indicated that the substance of SOD and CCI diminished in susceptible genotypes, while tolerant genotypes SOD and CCI stayed unaltered or increased. For measuring drought tolerance, the stress tolerance index (STI) used. The correlation between STI for Chlorophyll and Chlorophyll CI in drought was significant at 0.01 levels. The pressure resilience list (STI) for SOD and CCI characterized safe and defenseless genotypes into unmistakable gatherings.
Conclusion: Hence, these 2 characters can be utilized as a Selection index for screening dry spell safe plant materials.
Keywords: Durum wheat, Drought stress, SOD, STI
Introduction![]()
Tetraploid durum wheat, or (T. durum), is the basic ingredient of scrumptious semolina-based products like pasta and grits, and it’s the main material used for flour production [1, 2]. Despite the high disease-resistant and stress-resistant properties of tetraploid wheat, more area of land is allocated to the cultivation of hexaploid wheat [3]. Given the importance of multiple genes in yield and the considerable effect of heredity on it, genotype screening attempts should investigate components related to yield [4, 5]. DS can be one of the many environmental factors that hamper photosynthesis [6, 7]. The significant sensitivity of two photic systems II (PS II) to limiting tensions induced by the environmental factors causes DS to damage these systems that are reaction spots. Chlorophyll fluorescence techniques reveal that metabolism and the processes of production are not balanced [8]. ROSs are the unfavored products of the biochemical changes that exposure to environmental stress generates in plants [9]. Nevertheless, ROS can result in oxidative stress by elevating some types of ROS, including O2-, H2O2-, and OH- [10]. The toxicity of H2O2 is particularly more highlighted in the chloroplasts, as even low concentrations of H2O2 can disrupt the active enzymes in the Calvin cycle and impair the photosynthetic assimilation of carbon dioxide [11]. As with the variable and unstable nature of drought as an environmental phenomenon, it is necessary to make the required changes in policies to adapt our cultivation in these areas to drought changes and minimize the difference between actual production what crops can potentially produce [12]. The range of influential factors on chlorophyll amounts includes:
(A) The light intensity can not only affect the chlorophyll content amounts in the leaf but also it even can influence different chloroplasts array within the cells of the leaf. Shady plants have more chlorophyll compared with high light plants.
(B) The role of temperature in chlorophyll efficiency and yield is prominent, as a plant with four carbons at 30°C to 45°C and another with three carbons at 10°C to 25°C have the highest chlorophyll yield. (C) Photosynthetic activity rises in the early stages of leaf growth, going at a peak when the leaf is fully grown, then slowly diminishing as the leaf passes maturity, indicating the positive relationship (at least to some extent) between leaf age and its chlorophyll content [13]. Besides, cultivars with more sensitivity to DS exhibited a considerably higher decrease in the chlorophyll a/b ratio, while in the more drought-resistant cultivars, the reported decrease was insignificant [14]. Chlorophyll is the primary pigment in many plant species, and insufficiency in its density may lead to impaired growth, lower yield, and, more specifically, a relatively serious condition called chlorosis [15]. Plants’ solution for tackling the challenge of oxidative stress is developing and activating an efficient defensive line able to either eliminate free radicals or neutralize them. This mechanism of this defensive is split into two groups: enzymatic and non-enzymatic. The former is composed of SOD, catalysis, APX, and GR, while the latter consists of tocopherol, carotenoids, ascorbate, and other compounds (that is, flavonoids, mannitols, and polyphenols) [6]. The diversity of reactive oxygen sorts allows for the multiplicity and the high number of defensive mechanisms (http://tse-co.blogfa.com) in cells and different under-cell sectors. Also, these reactive oxygen sorts have multiple distinguishing features, such as diffusion capability, solubility, and high reactivity. As a result, the presence of a range of interconnected molecules with defensive functions in both the organic phase and the cell membrane seems essential for the ideal elimination of radicals in real-time as they are synthesized. Research studies also associate the presence of antioxidants with the shortage of water [10]. Reportedly, plants harboring higher antioxidant content are more resistant to oxidative damage, regardless of the inherent or induced origin of the antioxidants [16]. Adaptive processes to drought-induced stress often correlate with using the antioxidant system to maintain the almost low levels of ROS. This study discusses the evaluation of the DS impacts on the activity of SOD and CCI in durum wheat genotypes, which have various levels of stress tolerance, as well as the relationship between these two traits and the stability of chlorophyll.
Methods![]()
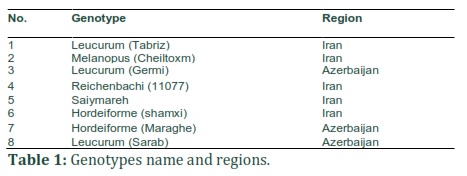
This study selected eight different durum wheat genotypes whose DS resistance was split into these levels: tolerant, semi-tolerant, and susceptible. The study took the seed samples each representing a single plant from northwestern Iran and The Republic of Azerbaijan Republic, as depicted in Table 1, to grow under stress-free (irrigated) and stress (under drought) conditions at the AES in the Islamic Azad University. From each genotype, ten leaf samples were collected. Seedlings were sampled on days 30, 35, and 39 of water stress. The study used CCM-200 to measure CCI made by Opti-sciences.
Determination of SOD activity
The procedure to detect levels of SOD activity in the genotypes included: Sampling leaf tissues for enzyme extraction and assays, freezing them in liquid nitrogen, grinding them, and maintaining them at -20 °C. Then, the study used 10 mol of 50 ml potassium phosphate buffer (pH 7.8) containing 1 μM EDTA (0.1 mM EDTA) and 1% (w/v) PVPP to homogenize 0.5 g of the frozen powder. Next, for centrifugation, the frozen powder was put into a centrifuge for 20 minutes at a temperature of 4 °C at 15,000 rpm. Also, the supernatant was immediately used as an enzyme source.
The study utilized the method proposed by Fridovich to assay the SOD activity (18), which, following the proposed method of Beyer and Fridovich, was defined based on the inhibition of NBT photochemical reduction (17). The following materials were included in the assay medium: 57 μM of NBT, 50 mM of phosphate buffer (pH 7.8), 9.9 mM of L-methionine, 0.025% (v/v) Triton-X 100, and 20 ml of enzyme extract. To start the reactions, we added a10 μl of aqueous riboflavin solution (44 μg ml-1) and put the tubes in a lined box wrapped with aluminum foil. The light source used included 20 W fluorescent lamps, which were switched on for seven minutes. We ran a parallel control where we had substituted buffer for the sample. Measurements indicated that with illumination, the absorbance level of the solution was 560 nm. By using a spectrophotometer to check the absorbance at 560 nm, the study managed to identify one unit of enzyme activity: it equaled the enzyme amount to reach a 50% NBT inhibition reduction rate (Shimadzu UV-120- 02) [10].
Statistical analysis
This study used ANOVA to analyze the data obtained from CCI and SOD activity and utilized the least significant difference (LSD) at a 0.05 probability level to differentiate between different treatments means. The study then used the formula and the STI suggested in a relevant study to measure the genotypes' drought resistance [17]:
STI= ((Ypi xYsi)/Yp2)
Ysi= yield in stress condition, Ypi= yield in normal condition
This study used HCA to divide genotypes based on DTI in the environment of SPSS 26.
Results![]()
Enzyme Activities
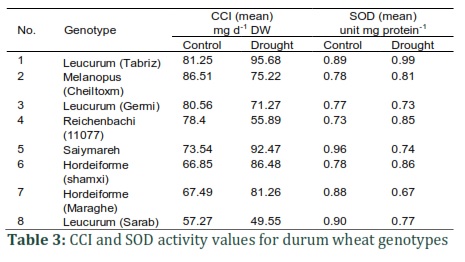
In the experiment, the genotypes showed significantly different levels of SOD activity. Moreover, as depicted in Table 2, the response to the environmental conditions varied, meaning that the variants used in the experiment showed different reactions to normal and DS conditions. Based on the findings of this study, genotype 5 had the highest SOD activity (0.96 unit mg protein–1) under normal stress conditions. Accordingly, as indicated in Table 3, genotypes 8 and 7 respectively showed 0.90 and 0.89 unit mg protein–1 activity rates. Leucurum (also known as Tabriz cultivar) represented higher SOD activity under normal farming conditions, whereas in the rest of the genotypes, enzyme activity declined. Famously, plants develop mechanisms specifically to help cope with ROS-induced stress; at the forefront of this defense line, SOD detoxifies the superoxide radicals. Based on observations, it is hypothesized that in genotypes with more significant elevation in SOD activity, the toxic concentration of O2- radicals may have decreased more efficiently than in genotypes without the mentioned elevation. Two factors are potentially associable with SOD activity reduction in response to drought: decline in production or the higher intensity of enzyme degradation.
Chlorophyll Content Index (CCI)
CCI variance analysis results indicated that genotypes showed a non-significant difference concerning the CCI under normal and stressful conditions. Despite that observation, as presented in Table 2, genotypes' interactions with the environment showed significant differences concerning CCI. It is hypothesized that drought stress causes the more significant CCI changes in genotypes, as depicted by Table 3. The study did a comparison regarding genotypes' considerable interactions with the environment, identifying genotypes 1 and 8 to respectively have the highest and lowest average CCI values in normal, non-stress conditions. After 39 days under stressful (drought) conditions, genotypes' leaf chlorophyll content exceeded the control level in this order: 1, 5, 6, and 7. Genotype 6 showed the highest increased percentage, followed by 7 and 1.
Stress tolerance index
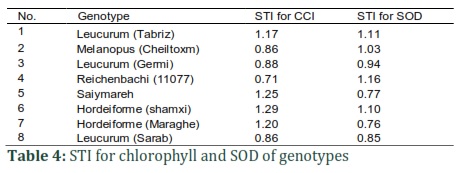
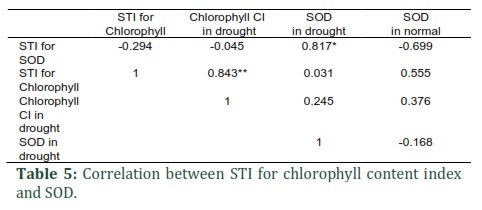
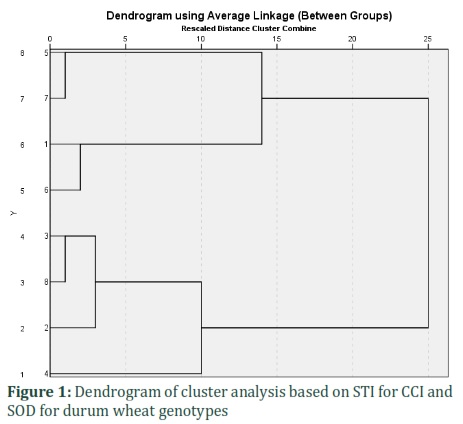
Table 4 depicts the SOD-based calculated STI and chlorophyll. Both traits showed similar results. Concerning the STI for SOD, the study identified the genotypes with the highest resistance and ordered them as the following: 4, 1, 6, 2, and 3; however, in terms of chlorophyll stress tolerance index, these genotypes were respectively identified as the most resistant ones: 6, 5, 7, 1, and 3. This indicates that SOD STI and chlorophyll STI's potential as identifying tools for finding varieties with significant resistance. Table 5 depicts the PCC results of the assessed physiologic traits. The observations indicated a significant (0.05) correlation of STI for SOD and SOD in drought. However, measurements of STI for Chlorophyll and Chlorophyll CI in response to drought were considerable at 0.01 level. As presented in Fig.1, the study divided the genotypes in the experiment into two main clusters based on cluster analysis results concerning CCI and SOD STI, chlorophyll content, and stress-induced SOD.
While genotypes 3, 8, 2, and 4 are identified as susceptible ones, as presented in the dendrogram, 5, 7, 1, and 6 put in the first and second clusters, indicating the application of this index for the identification of more resistant genotypes. The investigation of the considerable interaction between the genotypes and normal and stressful environmental conditions concerning the chlorophyll and SOD revealed that under drought and normal irrigation conditions, genotypes synthesized different amounts of SOD and chlorophyll.
Figures & Tables

Discussion![]()
Based on observations, most of the genotypes in the experiment exhibited lower under-stress CCI values in comparison with the control genotypes. The genotypes represented relatively similar patterns of SOD activity response and CCI changes under DS, except for few cases, suggesting the possible role of that drought in chlorophyll photo oxidation mediated by oxy-iradicals [18]. Genotypes’ average CCI in response to stress was lower than under normal conditions, associable with the relative DS-induced chlorophyll degradation. [19], in their study, proved that leaves’ chlorophyll content decreased in reaction to DS and shortage of water. The increasing levels of radicals and other ROSs that undermine chloroplasts, leading to chlorophyll degradation. However, stamen leaf represented elevated SOD values in response to drought and its associated stress, which is associable with SOD synthesis being part of the plant’s defensive mechanism against oxidative stress, triggered under drought and salinity conditions. Resistant verities exhibited higher stamen leaf SOD levels in comparison with the more susceptible varieties. In less resistant varieties, CCI and SOD STI have a corroborative negative correlation. As depicted by Table 5, though, more resistant genotypes have higher SOD and in-leaf CCI values, compared with the susceptible genotypes, making the SOD activity a practical index for detecting varieties with optimal drought resistance. Our findings are consistent with those reported by [10, 20-24]. STI selects genotypes with high performance under both stress and normal conditions [10, 25-29]. This study reported a considerable positive correlation between STI, SOD index, and chlorophyll. Moreover, the indexes this research study used appeared promising to be screening tools for distinguishing the more drought-resistant durum wheat varieties from the more sensitive ones due to this near-perfect distinguishing performance and the significant correlation mentioned. Nevertheless, it is necessary to explore the aboriginal species to identify plants with higher resistance to drought. Similarly, more research is needed to investigate the application of native genotypes in breeding more stress-resistant varieties.
Author Contributions
Majid Khayatnezhad and Roza Gholamin conducted, planned, analyzed the data, wrote manuscript and interpreted the results and involved in manuscript preparation.
The authors declare that they have no competing interests.
References![]()
- Gholamin R, Khayatnezhad M. Study of Bread Wheat Genotype Physiological and Biochemical Responses to Drought Stress. Helix, (2020); 10 (05): 87-92.
- Khayatnezhad M, Gholamin R. Study of Durum Wheat Genotypes' Response to Drought Stress Conditions. Helix, (2020); 10(05): 98-103.
- Khayatnezhad M, Gholamin R. Effect of nitrogen fertilizer levels on different planting remobilization of dry matter of durum wheat varieties Seimareh. African Journal of Microbiology Research, (2012); 6(7): 1534-1539.
- Khayatnezhad M, Gholamin R. The effect of drought stress on leaf chlorophyll content and stress resistance in maize cultivars (Zea mays). African Journal of Microbiology Research, (2012); 6 (12): 2844-2848.
- Almansouri M, Kinet JM, Lutts S, Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant and soil, (2001); 231 (2): 243-254.
- Khayatnezhad M. Evaluation of the reaction of durum wheat genotypes (Triticum durum Desf.) to drought conditions using various stress tolerance indices. African Journal of Microbiology Research, (2012); 6 (20): 4315-4323.
- Gholamin R, Khayatnezhad M. Effect of different levels of manganese fertilizer and drought stress on yield and agronomic use efficiency of fertilizer in durum wheat in Ardabil. Journal of Food, Agriculture & Environment, (2012); 10 (2 part 3): 1326-1328.
- Khayatnezhad M, Gholamin R. The effect of drought stress on leaf chlorophyll content and stress resistance in maize cultivars (Zea mays). African Journal of Microbiology Research, (2012); 6(12): 2844-2848.Dat J. Dual action of the active oxygen species during plant stress responses. Cellular and Molecular Life Sciences CMLS, (2000); 57 (5): 779-795.
- Zaefyzadeh M. The effect of the interaction between genotypes and drought stress on the superoxide dismutase and chlorophyll content in durum wheat landraces. Turkish Journal of biology, (2009); 33 (1): 1-7.
- Takeda T, Yokota A, Shigeoka S. Resistance of photosynthesis to hydrogen peroxide in algae. Plant and cell physiology, (1995); 36 (6): 089-1095.
- ORT DR. Chilling-induced limitations on photosynthesis in warm climate plants: contrasting mechanisms. Environment Control in Biology, (2002); 40(1): 7-18.
- Gholamin R, Khayatnezhad M. Assessment of the Correlation between Chlorophyll Content and Drought Resistance in Corn Cultivars (Zea Mays). Helix, (2020); 10 (05): 93-97.
- Loggini B. Antioxidative defense system, pigment composition, and photosynthetic efficiency in two wheat cultivars subjected to drought. Plant physiology, (1999); 119 (3): 1091-1100.
- Khosh K, Ando B. Effect of food environments, particularly sodium ion on the synthesis of chlorophyll and plant growth C4. in Abstracts Third Crop Science Congress of Iran. Tabriz University. (1995).
- Boo YC, Jung J. Water deficit induced oxidative stress and antioxidative defenses in rice plants. Journal of Plant Physiology, (1999); 155 (2): 255-261.
- Fernandez GC. Effective selection criteria for assessing plant stress tolerance. Adaptation of food crops to temperature and water stress, (1993); 410: 257-270.
- Wise RR, Naylor AW. Chilling-enhanced photooxidation: the peroxidative destruction of lipids during chilling injury to photosynthesis and ultrastructure. Plant physiology, (1987); 83 (2): 272-277.
- Zhang M. Uniconazole-induced tolerance of soybean to water deficit stress in relation to changes in photosynthesis, hormones and antioxidant system. Journal of Plant Physiology, (2007); 164 (6): 709-717.
- Moradi F, Ismail AM. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Annals of botany, (2007); 99(6): 1161-1173.
- Arora A, Sairam R, Srivastava G. Oxidative stress and antioxidative system in plants. Current science, (2002); 82 (10): 1227-1238.
- Gholamin R, Khayatnezhad M. The Study of Path Analysis for Durum wheat (Triticum durum Desf.) Yield Components. Bioscience Biotechnology Research Communications, (2020); 13(4): 2139-2144.
- Gholamin R, Khayatnezhad M. The Effect of Dry Season Stretch on Chlorophyll Content and RWC of Wheat Genotypes (Triticum Durum L.). Bioscience Biotechnology Research Communications, (2020); 13(4): 1829-1833.
- Khayatnezhad M, Gholamin R. A Modern Equation for Determining the Dry-spell Resistance of Crops to Identify Suitable Seeds for the Breeding Program Using Modified Stress Tolerance Index (MSTI). Bioscience Biotechnology Research Communications, (2020); 13(4): 2114-2117.
- Si X, Gao L, Song Y, Khayatnezhad M, Minaeifar AA. Understanding population differentiation using geographical, morphological and genetic characterization in Erodium cicunium. Indian Journal of Genetics, (2020); 80(4): 459-67.
- Jia Y, Khayatnezhad M, Mehri S. Population differentiation and gene flow in Erodium cicutarium: a potential medicinal plant. Genetika, (2020); 52 (3): 1127-1144.
- Karasakal A, Khayatnezhad M, Gholamin R. The Effect of Saline, Drought, and Presowing Salt Stress on Nitrate Reductase Activity in Varieties of Eleusine coracana (Gaertn). Biosc. Bioscience Biotechnology Research Communications, (2020); 13(4): 2087-2091.
- Karasakal A, Khayatnezhad M, Gholamin R. The Durum Wheat Gene Sequence Response Assessment of Triticum durum for Dehydration Situations Utilizing Different Indicators of Water Deficiency. Bioscience Biotechnology Research Communications, (2020); 13(4): 2050-2057.
- Rafay M, Abid M, Malik Z, Madnee M, Basit H, Ghaffar MU. Salinity tolerance and phytoremediation of Na+ and K+ ions by using halophytes from Cholistan Rangeland, Pakistan. Agrobiological Records, (2020); 2: 31-37.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


