

Full Length Research Article
Metabolic and molecular effects of edible oils on PPAR modulators in rabbit liver
Syed Nasir Ahamed1, Johra Khan2*, Syed Rahamathulla1, T.C. Venkateswarulu1, S. Krupanidhu1
Adv. life sci., vol. 8, no. 2, pp. 154-159, February 2021
*- Corresponding Author: Johra Khan (Email: j.khan@mu.edu.sa)
Authors' Affiliations
2. Department of Medical Laboratory Sciences, Majmaah University, Majmaah – Saudi Arabia
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Peroxisome proliferator-activated receptors are the receptors that switch-on DNA responding elements as heterodimers with nuclear retinoic acid. These are the transcription factors and belong to the family of nuclear receptors and key regulators for lipid and glucose metabolism.
Methods: The study was designed to investigate the effect of edible oil on PPAR Alpha & Gamma expression on rabbit liver.
Results: Reduced Glutathione (GSH) activity showed a significant increase and glutathione peroxidase (GPx) activity showed a significant decrease in batch II and batch III in comparison to control group was observed. Out of the total 3 batches, an increase was significantly increased in batch III (160%) in comparison to batch II (151%). Inedible oil-treated Batches, PPAR α, and PPAR γ levels were found to decrease significantly for the control batch. The decrease in PPAR alpha levels in batch II and batch III was 1.7 and 2.2 fold with p<0.01. In the case of PPAR gamma, the corresponding values increased by 1.6 and 2.4 fold respectively with p<0.01.
Conclusion: The study demonstrated that edible oil supplementation significantly increases mRNA levels up to two-fold in comparison to the control batch.
Keywords: PPARα; PPARγ; Edible oil; Reduced glutathione; Glutathione peroxidase
Introduction![]()
Lipids are one of the vital components of the diet and major sources of energy next to carbohydrates [1]. Lipids are a diversified and ever-present group of compounds, which are Fatty substances found within and between each cell and in the myelin sheath that coats and protects the nerves [2]. The liver is the key organ in controlling entire lipid metabolism like lipoprotein synthesis, secretion, and authorization, intracellular lipid metabolism in the rabbit body [3].
Several pathways that are mutually dependent and cross-regulated are involved in lipid metabolism. Fatty acids are common circulating forms of energy and triacylglycerols are the normal non-toxic fatty acid forms. Cytoplasmic triacylglycerol stores, de novo lipogenesis, fatty acids of lipoprotein remnants that are released by adipose tissues will be taken up via plasma non-esterified fatty acids (NEFA) [4]. Carnitine palmitoyltransferase (CPT), acetyl-CoA carboxylase (ACC), fatty acid synthase (FAS) are the major regulating elements in fatty acid production and oxidation [5]. The rate-limiting steps are hormone-sensitive lipase and lipoprotein lipase enzymes for the yield of adipose tissue fatty acids [5]. The cellular uptake and transport of fatty acids are regulated by the fatty acid-binding protein (FABP) and the fatty acid transport protein (FATP) [6].
Peroxisome proliferator-activated receptors (PPARs) perform the function in a ligand-dependent manner. The expression pattern of these transcription factors is indicative of the tissue functions [7]. Both β and ω oxidation pathways include PPAR-α in fatty acid catabolism, which is liver, adipose tissue [8]. PPAR-γ shows restriction in expression patterns and its primary sources include mammary glands, adipose tissues, and macrophages [9]. PPAR-γ is meant for the differentiation of adipose tissue in vivo and in vitro [10]. A few genes that are regulated by PPARγ include adipocyte fatty acid-binding protein, acyl-CoA synthetase, stearoyl-CoA synthetase, and phosphoenolpyruvate carboxykinase [11].
The innovation of the PPAR family of transcription factors has exposed the mechanism of the strong link among lipid/glucose availability. It shows long-term metabolic adjustment. PPARα plays a significant role in lipid metabolism, the well-known target genes are concerned with liver inflammation, lipoprotein metabolism, glucose metabolism, amino acid metabolism, and hepatocyte proliferation [11-13]. The role of synthetic agonists which are used in the clinical management of dyslipidemia is to decrease the plasma triglycerides and to increase plasma high-density lipoprotein levels [14,15]. The study of these contents would reveal the insights in exploring new physiological roles of PPARs and the identification of novel and advanced PPAR agonist drugs. It was found that, in a rabbit atherosclerosis model, PPAR-α activation can ease the atherosclerosis effects by the inhibition of major inflammatory factor expression [16]. The observations of have provided a molecular basis to understand the mechanism of how fibrates and glitazones affect lipid metabolism [17,18]. This identifies PPARs as important targets for generating a potential lipid-lowering drug [19]. Edible oil which is part of food in most southeast Asian countries and European countries is the cause of a high rate of hyperlipidemia as the lipids produced here are not utilized in urgent energy production, which means the lipids are stored in adipose tissues (fat depots) in the body and inside the liver causing fatty liver disease [19].
It was also reported that the perfluorooctanoic acid (PFOA) is a determined organic pollutant and established an in vivo peroxisome proliferator-activated receptor alpha silencing model in chicken embryo [20,21]. PPARα protects against liver toxicity made by AOM and CCl4 signifying that this receptor is hepato-protective in conflict to environmental chemicals which are metabolized in these tissue [22].
Methods![]()
Male albino rabbits of 1750 g each were kept in laboratory conditions with the temperature maintained between 15-21oC, humidity 45±10%, and 12 hr alternate light and dark period for 21 days with a mixed diet of pellets, green leafy vegetables, and water. All animals were divided into 3 batches with 6 rabbits in each batch. The batch I was kept as control, with a normal diet and water supplied regularly. Batch II was supplemented with 2 mL/kg of body weight dose of edible oil for 21 days, and batch III animals were given 5 mL/kg edible oil. After 21 days period, they were sacrificed and extracted liver tissues were homogenized.
Estimation of Protein using Bradford method:
This method is used to assess total protein concentration in homogenized tissue samples [23]. The standard protein buffer of concentration ranging from 0.1-1.4 mg/ml was used with the BSA standard. In a 96 well ELISA plate, 5ml protein samples were loaded and mixed with sample mixture prepared by mixing 1.2mg.ml sample with 250 µL of Bradford reagent, followed by incubation in shaker for 39 sec followed by 30 min incubation and measurement of absorbency at 595nm. A standard protein curve was used to assess protein concentration.
Biochemical Assays:
Glutathione Peroxidase (GPx) Assay:
GPx was measured by washing liver tissues with a phosphate buffer of pH 7.4. GPx was then estimated using the GPx Assay Kit [24].
Reduced Glutathione Assay:
For GSH assay tissue samples were collected from all three batches and estimation of GSH was done using Cayman Chemical Assay Kit.
Superoxide Dismutase estimation:
Superoxide dismutase (SOD) is an important class of metalloenzymes that obstruct the harmful effects of superoxide free radicals. The estimation of SOD was performed by keeping liver tissues in 0.9% NaCl for removing RBC, followed by blot drying tissues [25]. After weighing these tissues were homogenized and SOD was measured using SD assay kit (Oxis research, USA SOD- 525™).
ELISA based assessment:
PPARα and PPARγ levels in the liver were tested using a PPARα and PPARγ assay kit (R&D Systems, Minneapolis) as per company's instructions. Samples and diluted series of standards were subjected to incubation with monoclonal antibody of the mouse to PPARα and PPARγ for 3 hr at room temperature [26]. After a thorough wash, the addition of a substrate solution and stop solution followed by the optical density calculation at 450 nm was performed.
Assessment using Western Blotting technique:
Homogenized tissue extraction was performed using lysis buffer composed of RIPA-, 5 µg of Sodium deoxycholate, 10 µg of NP-40, and 10 µg of 0.1% of
RNA isolation, cDNA synthesis, and Real-Time PCR
Isolation of RNA and cDNA synthesis
Liver tissue samples of all three batches were exposed to liquid nitrogen, followed by RNA isolation using triazole under aseptic conditions. Nanodrop technique was used to check RNA concentration in sample and cDNA was produced using applied biosystem (cat no- 4368814) following manufactures instructions.
Real-Time Rt-PCR
The RT Mix was prepared as per the manual by preparing 10 μl of 2 X RT master mix having 10μl of RNA sample, 10X RT buffer, 10X RT Random Primers, 25X dNTP Mix, MultiScribe™ Reverse Transcriptase, RNAse Inhibitor, and Nuclease-free water. On a Real-Time PCR System 7500 (Applied Biosystems), the reaction was performed. The sense and anti-sense primers used are tabulated (Table 1).
Results![]()
The wet lab experiments were conducted with three different batches of animals and the data was analyzed for mean ± SD. One way ANOVA was performed followed by Dunnet’s test for comparison between control and respective batches, using sigmablot software. At a 5% level, the statistical significance was calculated. The total protein, when estimated using Bradford reagent, in rabbit liver, was found to be 3.74µg/µL.
Biochemical assay
Results show a significant decrease in batch II and batch III GPx activity in comparison to the control group emphasizing the presence of OS (oxidative stress) and reduced the anti-oxidative defense systems. And like in the case of
PPARα and PPARγ levels in rabbit liver
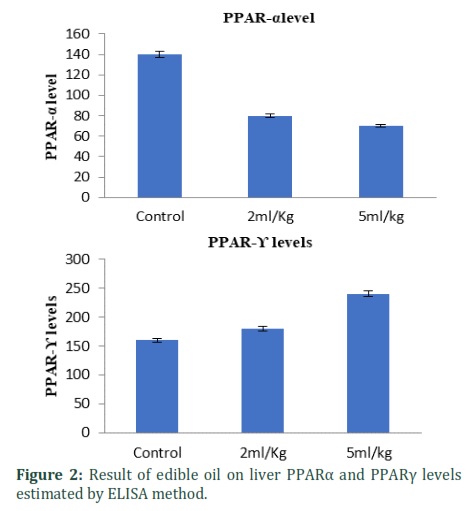
The effect of oil supplemented on liver PPARα and PPARγ levels were assessed by ELISA. Enhancement in liver PPARα and PPARγ levels was observed when oil was fed for the long term. Precisely, it was 2.1 and 1.2 fold (p < 0.01) decreased production by medium and low-dose of oil respectively in liver PPARα levels. Similarly, 1.6 and 2.4 fold (p < 0.01) increase in liver PPARγ levels by medium and high-dose of oil respective that of control batch (Fig. 2).
Effect of edible oil on PPARα and PPARγ expression
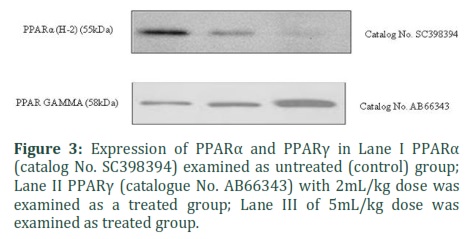
PPARα and PPARγ protein concentration were assessed in homogenate liver tissue of control and batch II and III. Homogenate tissues were exposed to Western blot using anti-PPARα and PPARγ and anti-Actin antibodies. The findings shows that PPARα expression was decreased as is seen in lane II and Lane III when compared to Lane I and PPARγ expression was increased as is seen in lane II and Lane III (Fig. 3). Actin was determined by Western blot with definite antibodies in the Rabbit liver. Effect of Lane II and Lane III on Beta Actin have seen as significantly no changes in comparison to Lane I (Fig. 4)
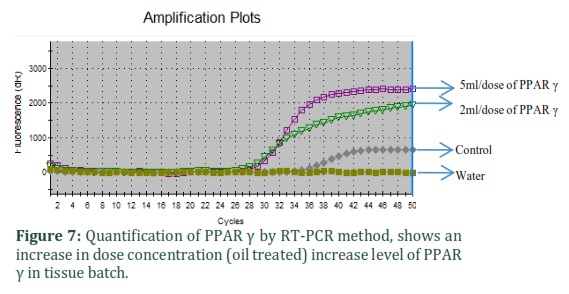
Quantitative RT-PCR
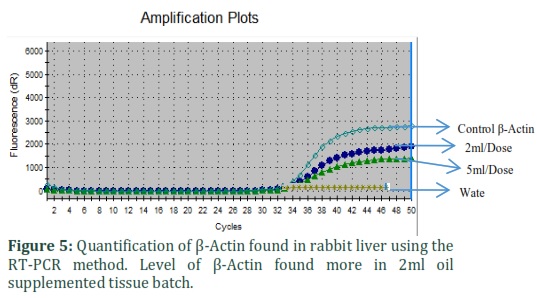
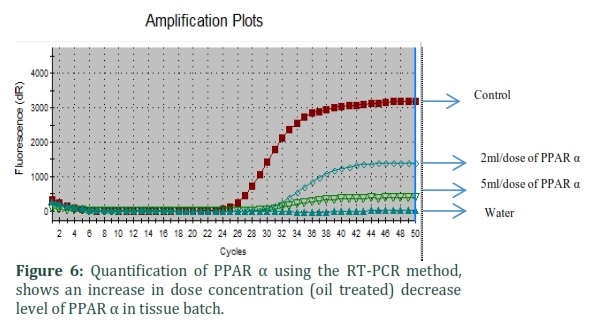
Results from quantitative RT-PCR assays showed that mRNA for PPARα, PPARγ, and β-actin were present in rabbit liver tissue. Concentrations of RT-PCR products for PPARα were significantly decreased and PPARγ were expressively increased in the tissues of oil complemented batch.
Figures & Tables



Discussion![]()
In this study, the biochemical and molecular changes in rabbits fed on oils excessively for about 21 days were illustrated. This study shows that there is significant decrease in the activities of SOD, and GPx in group II and group III when compared to the control group. The down-regulation and upregulation of PPAR-α and PPAR γ respectively in oil-fed rats indicates profuse adipogenesis in the liver, increased oxidative stress which finally leads to conditions like diabetes, obesity, and cardiovascular diseases. PPARs belonging to the superfamily of nuclear receptors are the transcription factors existing in three isoforms. In the present research, we studied the role of edible oils in altering the metabolic and molecular events that occur in the rabbit liver via PPAR α & γ. Bradford method and antioxidant content (
As we know SOD functions as the first line of defense especially in the response of oxidative stress [27], and play a significant part in the disputation of superoxide anions from H2O2. GPx breaks down H2O2 to H2O and O2. The rabbit feed with edible oil was found to have a significant decrease in these enzymes which confirms the function of edible oil in the decomposition of H2O2 and superoxide. The decrease in GSH concentration shows the reduced antioxidant system in liver tissues. The possible reason for oxidative stress is the accumulation of fat in the adipose tissues of the rabbit liver. Fatty acid and fat accumulation can increase OS by liberating ROS in tissues.
Further in our studies, it was observed that PPAR α mRNA and in turn its protein levels were significantly decreased whereas PPAR- γ mRNA and its protein levels showed a significant increase in oil-fed rabbits. This indicates increased adipogenesis, and the modulation of gene expression (involved in lipid metabolism) by the transcription factors, PPARs. PPARγ, a type II nuclear receptor controls various cellular processes. It is included in the generation of various diseases like diabetes, obesity, cancer, and atherosclerosis [28]. The date indicates the abundant expression of PPAR in mature adipocytes and animal tissues [29]. PPARγ and its gene CD36 are up-regulated in the rabbit liver after two weeks of the feeding of a high-fat diet.
The hypothesis of PPARγ inhibiting the accumulation of lipids and the formation of foam cells by cholesterol efflux activation from macrophages via PPARα in vivo is supported by the current correlated expression of PPARα and PPARγ. Therefore, supplementation of oil induces oxidative damage of hepatocyte which will not affect the oxidative damage related proteins and genes inducing the signaling network being evoked by infrasound as PPARα and PPARγ are involved in infrasonic hepatic oxidant-antioxidant balance.
Author Contributions
SNA, JK: data collection and manuscript drafting, SNA: tables and figures, TCV, SK, SR: study design and supervision
Financial Support and Sponsorship
The author would like to thank the Deanship of Scientific Research at Majmaah University for supporting this work under Project Number No. 64/24172.
Authors declare no conflict of interest in this work.
References![]()
- Mohdaly AA, Smetanska I, Ramadan MF, Sarhan MA, Mahmoud A. Antioxidant potential of sesame (Sesamum indicum) cake extract in stabilization of sunflower and soybean oils. Industrial Crops and Products, (2011); 34(1): 952-959.
- Hussain SA, Hameed A, Ajmal I, Nosheen S, Suleria HAR, et al. Effects of sesame seed extract as a natural antioxidant on the oxidative stability of sunflower oil. Journal of food science and technology, (2018); 55(10): 4099-4110.
- Lei L, Xiaoyi S, Fuchang L. Effect of dietary copper addition on lipid metabolism in rabbits. Food & nutrition research, (2017); 61(1): 1348866.
- Taskinen M-R, Borén J. New insights into the pathophysiology of dyslipidemia in type 2 diabetes. Atherosclerosis, (2015); 239(2): 483-495.
- Khan J, Alaidarous MA, Naseem A. Dyslipidemia Relationship with Socioeconomic Status in East Champaran Population. International Journal of Pharmaceutical Research & Allied Sciences, (2020); 9(2).
- Khan J. Magnesium Deficiency and its Correlation with Insulin Resistance in Obese Females in Majmaah. International Journal of Pharmaceutical Sciences Review and Research, (2018); 49(2): 117-119.
- Umemoto T, Fujiki Y. Ligand‐dependent nucleo‐cytoplasmic shuttling of peroxisome proliferator‐activated receptors, PPARα and PPARγ. Genes to Cells, (2012); 17(7): 576-596.
- Pirat C, Farce A, Lebègue N, Renault N, Furman C, et al. Targeting peroxisome proliferator-activated receptors (PPARs): development of modulators. Journal of medicinal chemistry, (2012); 55(9): 4027-4061.
- Smeets PJ, Planavila A, Van Der Vusse GJ, Van Bilsen M. Peroxisome proliferator‐activated receptors and inflammation: take it to heart. Acta Physiologica, (2007); 191(3): 171-188.
- Cipolletta D, Feuerer M, Li A, Kamei N, Lee J, et al. PPAR-γ is a major driver of the accumulation and phenotype of adipose tissue T reg cells. Nature, (2012); 486(7404): 549-553.
- Lage R, Diéguez C, Vidal-Puig A, López M. AMPK: a metabolic gauge regulating whole-body energy homeostasis. Trends in molecular medicine, (2008); 14(12): 539-549.
- Boitier E, Gautier J-C, Roberts R. Advances in understanding the regulation of apoptosis and mitosis by peroxisome-proliferator activated receptors in pre-clinical models: relevance for human health and disease. Comparative hepatology, (2003); 2(1): 1-15.
- Kliewer SA, Sundseth SS, Jones SA, Brown PJ, Wisely GB, et al. Fatty acids and eicosanoids regulate gene expression through direct interactions with peroxisome proliferator-activated receptors α and γ. Proceedings of the National Academy of Sciences, (1997); 94(9): 4318-4323.
- Calabresi L, Villa B, Canavesi M, Sirtori CR, James RW, et al. An ω-3 polyunsaturated fatty acid concentrate increases plasma high-density lipoprotein 2 cholesterol and paraoxonase levels in patients with familial combined hyperlipidemia. Metabolism, (2004); 53(2): 153-158.
- Guan Y, Breyer M. Targeting peroxisome proliferator-activated receptors (PPARs) in kidney and urologic disease. Minerva urologica e nefrologica, (2002); 54(2): 65-80.
- Zambon A, Gervois P, Pauletto P, Fruchart J-C, Staels B. Modulation of Hepatic Inflammatory Risk Markers of Cardiovascular Diseases by PPAR–α Activators: Clinical and Experimental Evidence. Arteriosclerosis, Thrombosis, and Vascular Biology, (2006); 26(5): 977-986.
- Barbier O, Torra IP, Duguay Y, Blanquart C, Fruchart J-C, et al. Pleiotropic actions of peroxisome proliferator–activated receptors in lipid metabolism and atherosclerosis. Arteriosclerosis, thrombosis, and vascular biology, (2002); 22(5): 717-726.
- Lalloyer F, Staels B. Fibrates, glitazones, and peroxisome proliferator–activated receptors. Arteriosclerosis, thrombosis, and vascular biology, (2010); 30(5): 894-899.
- Shearer BG, Billin AN. The next generation of PPAR drugs: do we have the tools to find them? Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids, (2007); 1771(8): 1082-1093.
- Steenland K, Fletcher T, Savitz DA. Epidemiologic evidence on the health effects of perfluorooctanoic acid (PFOA). Environmental health perspectives, (2010); 118(8): 1100-1108.
- Bonato M, Corrà F, Bellio M, Guidolin L, Tallandini L, et al. PFAS environmental pollution and antioxidant responses: an overview of the impact on human field. International Journal of Environmental Research and Public Health, (2020); 17(21): 8020.
- Ross SA, Davis CD. The emerging role of microRNAs and nutrition in modulating health and disease. Annual review of nutrition, (2014); 34305-336.
- Wang W, Tai F, Chen S. Optimizing protein extraction from plant tissues for enhanced proteomics analysis. Journal of separation science, (2008); 31(11): 2032-2039.
- Yoshida S, Kumakura F, Komatsu I, Arai K, Onuma Y, et al. Antioxidative glutathione peroxidase activity of selenoglutathione. Angewandte Chemie, (2011); 123(9): 2173-2176.
- Bafana A, Dutt S, Kumar A, Kumar S, Ahuja PS. The basic and applied aspects of superoxide dismutase. Journal of Molecular Catalysis B: Enzymatic, (2011); 68(2): 129-138.
- Syversen U, Stunes AK, Gustafsson BI, Obrant KJ, Nordsletten L, et al. Different skeletal effects of the peroxisome proliferator activated receptor (PPAR) α agonist fenofibrate and the PPARγ agonist pioglitazone. BMC Endocrine Disorders, (2009); 9(1): 10.
- Jain MR, Giri SR, Trivedi C, Bhoi B, Rath A, et al. Saroglitazar, a novel PPARα/γ agonist with predominant PPARα activity, shows lipid‐lowering and insulin‐sensitizing effects in preclinical models. Pharmacology Research & Perspectives, (2015); 3(3): e00136.
- Inoue M, Ohtake T, Motomura W, Takahashi N, Hosoki Y, et al. Increased expression of PPARγ in high fat diet-induced liver steatosis in mice. Biochemical and biophysical research communications, (2005); 336(1): 215-222.
- Lefebvre P, Chinetti G, Fruchart J-C, Staels B. Sorting out the roles of PPARα in energy metabolism and vascular homeostasis. The Journal of clinical investigation, (2006); 116(3): 571-580.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


