

Full Length Research Article
Acclimatization of drought tolerance with Somaclonal variants of sugarcane (Saccharum officinarum L.)
Rashda Naheed1,2, Muhammad Arfan1, Fozia Farhat2*, Siddra Ijaz3, Hamza Khalid4
Adv. life sci., vol. 8, no. 1, pp. 57-62, November 2020
*- Corresponding Author: Fozia Farhat (Email: foziafarhat@gcwuf.edu.pk)
Authors' Affiliations
2. Department of Botany, Government College Women University, Faisalabad – Pakistan
3. Centre of Agricultural Biochemistry and Biotechnology, University of Agriculture, Faisalabad – Pakistan
4. Department of Microbiology, University of Agriculture, Faisalabad – Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Sugarcane is basically a water loving crop and scarcity of water is not only affecting its overall production but its total sugar contents as well. Somaclonal variation is an intriguing phenomenon to study various genetic and cellular mechanism under in vitro conditions. The induction of somaclonal variation generates useful variability without sexual reproduction in sugarcane.
Methods: CPF-248, a drought sensitive variety was used to induce stress tolerance using somaclonal variation approach. Various combinations of callogenesis and regeneration media were used to induce soma clonal variations. The plants, thus achieved, were subjected to drought selection pressure using different concentrations of PEG, the selected plants were referred as in vitro selected putative somaclonal variants 1 & 2 (IPSV1 & IPSV2). These two plants were then subjected to acclimatize under greenhouse conditions by planting them in soil filled plastic pots. The in vitro grown seedlings named as in vitro selected putative somaclonal variants along with parent plants were subjected to drought conditions by withholding watering for 8 days. After 10 days of treatment, data for photosynthetic and biochemical attributes were recorded and subjected to comparison. The activity of antioxidants enzymes like superoxide dismutase (SOD), ascorbate peroxidase (APX), catalase (CAT), peroxidase (POD), ascorbate and carotenoid contents was analyzed using standard protocols.
Results: A significant increase in chlorophyll a, b and carotenoid contents were recorded in IPSV1 and IPSV2 compared to parent genotype. Antioxidant enzymes (SOD, APX, CAT, POD) also increased significantly in somaclonal variants and displayed tolerance under drought condition. Moreover, osmotic adjustment was observed in the form of higher rate of total free amino acid and proline contents in both somaclonal variants.
Conclusion: The finding suggests that sugarcane (CPF-248) somaclones generated through tissue culture approach are found to be drought tolerant with improved photosynthesis and antioxidant response. Further, somaclonal variant IPSVI, exhibited better response than other variant (IPSV2).
Keywords: Drought; Antioxidants; Osmoprotectants; Somaclones; Sugarcane
Introduction![]()
As an important cash crop, sugarcane is a major contributor of sugar industry in the world. It is a non-cereal member of family Poaceae. Sugarcane belongs to the C4 group of plants. Although it has a more advanced type of energy conversion system as a tropical crop yet, it requires an optimal amount of water (1200 – 2500 mm) to provide optimum yields [1].
Water scarcity is among the several abiotic stresses that most drastically reduces the functioning of sugarcane cultivation, which is the main reason of falls in yield [2]. There is increasing demand for the sugarcane cultivars showing better improvements or tolerance against the more severe and prolonged drought conditions. This urges the better understanding of the physiological and biochemical responses against drought stress [3].
The ROS production enhances due to the exposure of the plant to the drought stress which causes an imbalance of redox system of the cell thus causing, unsettling of the electron transport system and the initiation of oxidative damage [4]. Plants adopted various mechanisms to detoxify the reactive oxygen species [5]. Several antioxidants are working for the scavenging the ROS. These antioxidants are either of enzymatic (CAT, SOD, POD, APX) or non-enzymatic (ascorbic acid, carotenoids) nature [6]. When a comparison is made between the drought tolerant and drought sensitive varieties, it becomes efficient that the tolerant varieties have a preference over the sensitive genotypes for readily recovery.
Plants or seedlings produced as a result of in vitro culturing from undifferentiated cells have been testified to reveal a high frequency of variations. The variations induced through tissue culture are best regarded to as somaclonal variations and are defined as the genetic/epigenetic variations [7,8]. Without any doubt, it is appropriate to recommend somaclonal technique to manipulate genetic makeup and for in vitro selection of desired traits in plants [9]. The primary aim of the present study was to screen the somaclones of sugarcane sensitive genotype (CPF-248) for drought tolerance using PEG (6000) as a selection component. In our second trial, examined the drought tolerant somaclones in the green house. The tolerance in sugarcane somaclones (IPSVI, IPSV2) were assessed through photosynthetic and antioxidant response compared to parent genotype (CPF-248) under prevailing drought conditions in the green house.
Methods![]()
Current research was carried out in Centre of Agriculture and Biochemistry in collaboration with Department of Botany, University of Agriculture, Faisalabad. Prior to this particular part of experiment, genotype CPF-248, a drought sensitive genotype of sugarcane was collected from Ayub Agriculture Research Institute (AARI), Faisalabad and used to induce stress tolerance by somaclonal techniques in a separate experiment. Various Callogenesis and regeneration media were used to induce somaclonal variations [10]. The plants thus achieved were exposed to drought selection pressure using different concentrations of PEG, the selected plants were referred as in vitro selected putative somaclonal variants 1 & 2 (IPSV1 & IPSV2).
These somaclonal variant (IPSV1, IPSV2) were then imperiled to acclimatization by shifting them in greenhouse environment. For acclimatization, plants were shifted into peat moss filled polythene bags and then transferred to green house. As the plants were established, they were subjected to drought stress by withholding water for 8 days. After two days of re-watering, the data was collected for different biochemical parameters in order to evaluate the drought tolerance capacities of the in vitro selected putative somaclonal variants and compared their results with the control (parent plant subjected to similar conditions).
Determination of Chlorophyll contents
The chlorophyll contents were determined with the method as described by Arnon [11]. Fresh leaves (0.5g) of both somaclonal variants and parent genotypes were chopped and dipped in 80% acetone (5mL). Solvent mixture was placed in dark for 24h. The resultant extract was subjected to record its absorbance at 480, 645 and 663 nm using a spectrophotometer (IRMECO U2020). Chlorophyll a, b and carotenoids were determined by applying respective formulas.
Determination of Antioxidants
To extract antioxidant enzymes, fresh leaves (0.5 g) were homogenized in 5 mL of 50 mM cooled phosphate buffer (pH 7.8). The homogenate was centrifuged at 15000 rpm for 20 min at 4°C. The supernatant was used to access activities of antioxidant enzymes. The activity of SOD was determined by measuring its ability to inhibit the photoreduction of nitroblue tetrazolium (NBT) by following the method of Ginnopolitis and Rice [12]. The absorbance of the irradiated solution was recorded at 560 nm using a spectrophotometer (IRMECO U2020). One unit of SOD activity was defined as the amount of enzyme that inhibited 50% of NBT photoreduction.
Activities of catalase (CAT) and peroxidase (POD) were measured using a slightly modified method of Chance and Maehly [13]. Change in absorbance was recorded at 240nm with an interval of 20s for 2 minutes. One unit CAT activity was defined as an absorbance change of 0.01 units per minute. One unit POD activity was defined as an absorbance change of 0.01 units per minute [14].
The ascorbate peroxidase (APX) activity was determined by adopting method of Nakano and Asada [15]. The activity of APX was accessed in 50 mM potassium phosphate buffer (pH 7.0) containing 0.5 mM AsA and 0.1 mM hydrogen peroxide and enzyme extract (0.1µL) in a total volume of 1 ml (Nakano and Asada, 1981). The hydrogen peroxide-dependent oxidation of AsA was followed by monitoring the decrease in absorbance at 290 nm assuming an absorption coefficient of 2.8 mM–1 cm–1.
The proline content in the fresh leaf was estimated according to the method of Bates [16]. Ground 0.5g tissue in 5 ml of 3% sulfosalicylic acid, filtered and mixed 2 ml filtrate with ninhydrin reagent (2Ml) and glacial acetic acid (2 ml), let the mixture heated in a boiling water bath for an hour. The heated mixture was cooled immediately in an ice bath, added 4 ml toluene, separated the chromosphere in the toluene fraction and optical density was recorded at 520 nm with a spectrophotometer. Prepared standard graph using 10, 20, 40, 80 and 100 µg/ml of proline. Calculated µg proline/ g fresh weight (Fr Wt.) by using the formula (µg/ml proline x vol. of toluene x volume of SS acid)/ (g Fr. Wt x 115.5).
Statistical analysis
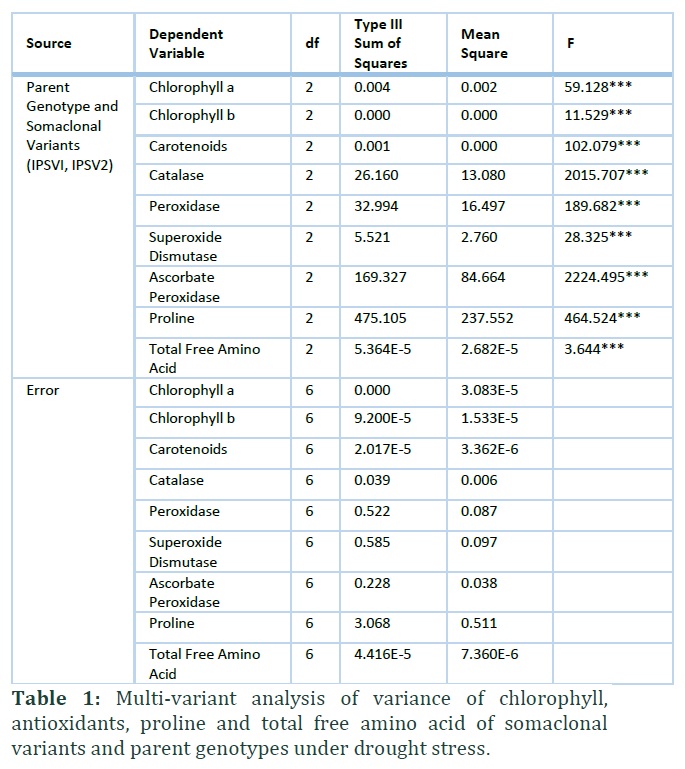
Somaclonal variants were laid down in greenhouse with completely randomized design (CRD) with three replicates. Multivariate analysis of variance (MANOVA) was performed with SPSS (23.0) at significance difference of p≤0.05.
Results![]()
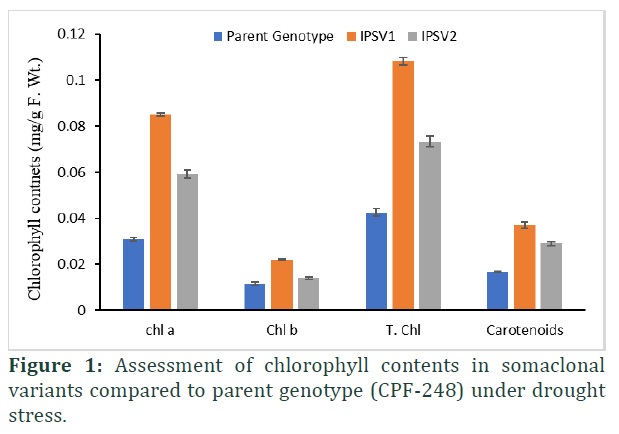
Data of somaclonal variants and parent genotype showed a significant (p≤0.00) difference in their chlorophyll contents under drought stress (Table 1). However, it was observed that parent genotype is extremely sensitive to drought treatment but somaclonal variants induce significance tolerance against drought and showed enhanced activity of chl a, chl b and total chlorophyll contents along with carotenoids (Figure 1). Approximately, 63% and 46% increase in chl a and chl b contents under drought stress in IPSV1 was observed compared to parent plant (Figure 1). The same trend was observed with respect to carotenoid contents that exhibited 54% and 42% increase in IPSV1 and IPSV2 respectively under drought stress (Figure 1). Chlorophyll contents exhibited a good success rate of acclimatizing under greenhouse conditions through somaclonal variants produced under in vitro growth.
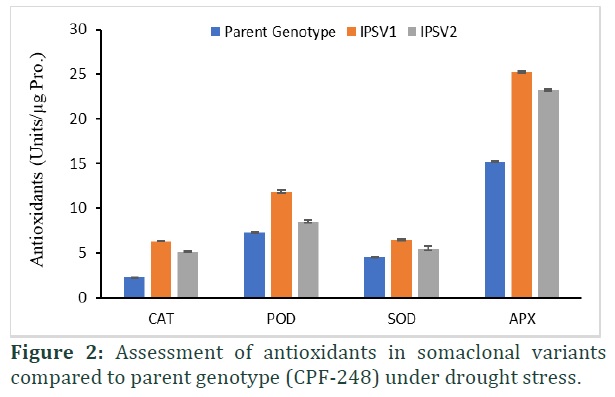
Antioxidant activity provides strong evidence for their tolerance against drought stress. Current experiment showed a significant (p≤0.00) difference in their antioxidants in both somaclonal variants and parent genotypes (Table 1). Ascorbate peroxidase (APX) activity increase (39% and 34%) substantially in both somaclonal variants with limited supply of water among all antioxidant enzymes studied in current study (Figure 2). Moreover, catalase (64%, 59%), peroxidase (38%, 14%) and superoxide dismutase (29%, 17%) also showed comprehensive increase under drought stress in IPSVI and IPSV2, respectively (Figure 2). Somaclonal variant displayed mitigation response against limited water supply in greenhouse environment and showed successful acclimatization as well.
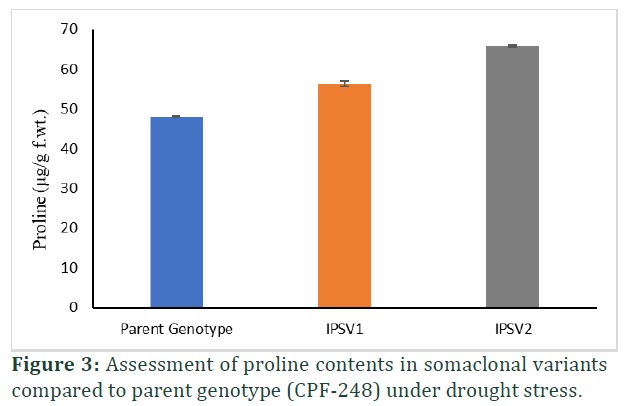
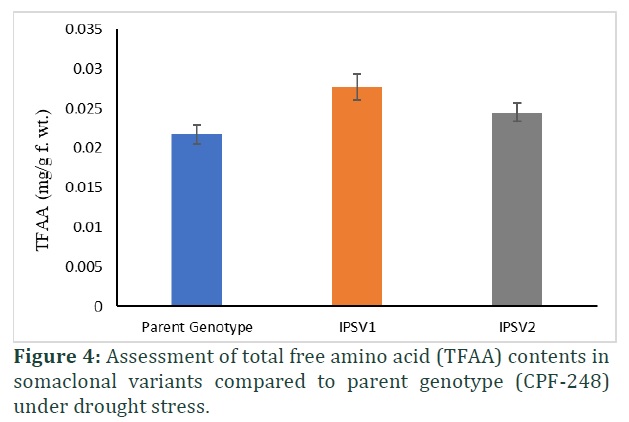
Total free amino acid and proline contents maintain metabolism to ensure survival of plants under stress. Proline contents increased significantly (p≤0.00) in both somaclonal variants compared to parent genotype under drought stress (Table 1). Somaclonal variant (IPSV2) showed 27% increase in proline contents followed by 14% increase in IPSV1 with drought stress (Figure 3). Total free amino acid also increased significantly (p≤0.00) in both somaclonal variants compared to parent genotype under drought stress (Table 1). It is observed that TFAA enhanced 21% and 11% in somaclonal genotypes (Figure 4).
Figures & Tables
Discussion![]()
Plant has to survive in a continuously changing environment where it has to face many biotic and abiotic stresses which are translated in to loss of vigor, reduced yields and much morphological and physiological impairment. The production of stress tolerant plants is the very first priority of the agronomists and researchers. For this purpose, among the different adapted techniques, in vitro selection of tissue culture based plants is the most cost effective and rapid practice. The selected plants may show genetic stability but will be the plants, cells or tissues that have shown tolerance against stress conditions [17].
Photosynthesis is a vital process for plants to grow. This process of imperative nature is under the control of different photosynthetic pigments like chlorophyll a, chlorophyll b and carotenoids play a pivotal role in harvesting of light energy and also in the running of Calvin Cycle. Like all other organic compounds these pigments are also susceptible to water deficit conditions [18]. A diminution in the levels of chlorophyll content is obvious when plants are exposed to water deficit [17, 18]. The chlorophyll a content of drought sensitive varieties showed a more decline than those of drought tolerant varieties. The drought stress not only caused the reduction in chlorophyll a and chlorophyll b contents but also changed the ratio of the two pigments [19]. The decrease in chlorophyll contents under drought stress is the result of oxidative damage which resulted in enhanced activity of antioxidants like carotenoids, ascorbates and flavonoids etc. [20].
Plants are continuously exposed to different types of environmental stresses resulting in the production of ROS (reactive oxygen species) which, due to their highly toxic and reactive nature causes the damage to the vital organic molecules and organelles of the cell. Different types of enzymatic and non-enzymatic antioxidants are produced by the plants to scavenge these toxic molecules of ROS. Superoxide dismutase forms the very first line of defense against reactive oxygen species by dismutation of O2 thus preventing the production of OH ions [21]. As a C4 plant sugarcane has the ability of concentrating CO2 even during stomatal closure, a first defensive approach adapted by the plants against drought stress. So the effect of stomatal closure is less in sugarcane as compared to C3 plants [22]. In this research study, an enhanced SOD activity was observed in IPSV1 and IPSV2 as compared to parent plant showing their preference in combating the drought stress over the parent plant. Increased activity of SOD is translated in better adaptations of the plant to fight against the oxidative stress. An increase in SOD activity against drought and salinity stress has been observed in Glycyrrhiza uralensis Fisch [23]. Water stress in Trifolium repens L. was reported to cause increased levels of SOD activity in the total SOD of leaf and SOD of chloroplast [24]. There is the evidence that the plants having foreign genes for improving stress tolerance showed the improvement in the SOD activity as a sign of tolerance of these transgenic plants over their wild relative [25,26]. An increase in the SOD activity is the indicator of drought tolerance and vice versa [27].
Ascorbic acid also known as Vitamin C is the most powerful non-enzymatic antioxidant. As a most abundant and water soluble antioxidant it is mostly present in photosynthetic and meristematic cells in a reduced form [28, 29]. High levels of ascorbic acid have been observed in seedlings of P. asperata [30] under drought and high light intensity. Maintenance and enhancement of APX and CAT is also related with the oxidative stress. The concentrations of these antioxidants is inversely proportional to the damage caused by oxidative stress in sugarcane [31, 32]. The results of this study also showed an increased activity of ASH, CAT and APX by IPSV1 and IPSV2. DaCosta and Huang (2007) and Chagas et al .m, (2008) also observed the increase in activity of CAT and SOD in drought tolerant genotypes of sugarcane as compare to sensitive genotypes. Cia et al., (2012) and Boaretto et al., (2014) observed the quenching role of these antioxidants against ROS production.
Since long proline has been known as osmoprotectant but its role as antioxidant is also being established now. Free proline has a variety of roles vital for the plant survival like its role as metal chelator, protein stabilizer, inhibitor of lipid per oxidation [37,38] and most importantly as osmoprotectant. As a nonenzymatic antioxidant, it is present in almost all types of living organisms to mitigate the deleterious effects of reactive oxygen species [39]. In the present study, the increase in concentration of proline in IPSV1 and IPSV2 over parent plant is also an indicator of their drought tolerance ability. Various studies revealed that increase in proline concentration is associated with stress tolerance capacity of the plant which in turn is related with the ability to maintain the compatible ratio of NADP : NADPH [40-44]. Improved proline synthesis under water stress has been concerned as a means of improving cytoplasmic acidosis. The degree of tolerance to drought stress is strongly co related with the increase in concentration of proline. Many researchers in their studies have also observed an increase in free proline concentrations under drought stress as an indicator of drought tolerant phenomena [45]. Free amino acids also played their role as osmoprotectants [45].
Increased activities of antioxidants and high accumulation of osmoprotectants as compared to parent genotype are the indicators of more stress tolerance capacities of IPSV1 &IPSV2 over parent genotype.
Author Contributions
Dr. Rashda Naheed (Principle Author) conducted the research trial in the laboratory as well as in greenhouse and got reproducible data of antioxidants and photosynthetic attributes of both sugarcane somaclonal variants and parent genotype. I’m highly honored to have Dr. Arfan as supervisor, who provided all the technical support to design the experiment. Dr. Siddra Ijaz (Assistant Professor, CABB, UAF) has facilitated laboratory conditions for smooth conduction of experiment. Dr. Fozia Farhat is a kind colleague and research fellow, who helped me in collecting data and compiling the manuscript in its final form. I’m also thankful to Mr. Hamza Khalid for his unconditional support to accomplish research work.
There is no conflict of interest regarding the publication of this paper.
References![]()
- Singh PN, Shukla SK, Bhatnagar VK. Optimizing soil moisture regime to increase water use efficiency of sugarcane (Saccharum spp. hybrid complex) in subtropical India. Agricultural Water Management, (2007); 90: 95-100.
- Dedemo GC, Rodrigues FA, Roberto PG, Neto CB, Franca SC, Zingaretti SM. Osmoprotection in sugarcane under water deficit conditions. Plant Stress, (2013); 7: 1-7.
- Ashraf MFMR, Foolad M. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental and experimental botany, (2007); 59: 206-216.
- Gratao PL, Monteiro CC, Antunes AM, Peres LEP, Azevedo RA. Acquired tolerance of tomato (Lycopersicon esculentum cv. Micro‐Tom) plants to cadmium‐induced stress. Annals of Applied Biology, (2008); 153: 321-333.
- Kellos T, Timar I, Szilagyi V, Szalai G, Galiba G, Kocsy G. Stress hormones and abiotic stresses have different effects on antioxidants in maize lines with different sensitivity. Plant Biology, (2008); 10: 563-572.
- Martins S, Mussatto SI, Martínez-Avila G, Montanez-Saenz J, Aguilar CNJA and Teixeira. Bioactive phenolic compounds: production and extraction by solid-state fermentation. A review. Biotechnology advances, (2011); 29: 365-373.
- Bairu MW, Aremu AO, Staden JV. Somaclonal variation in plants: causes and detection methods. Plant Growth Regul, (2011); 63: 147–173.
- Currais L, Loureiro J, Santos C, Canhoto JM. Ploidy stability in embryogenic cultures and regenerated plantlets of tamarillo. Plant Cell Tissue Organ Cult, (2013); 114: 149–159.
- Roopadarshini V, Gayatri MC. Isolation of somaclonal variants for morphological and biochemical traits in Curcuma longa (Turmeric) Res Plant Biol, 2012; 2: 31–37.
- Naheed R, Arfan M, Ijaz S, Shahbaz M. Induction of somaclonal variation in selected drought sensitive genotype of sugarcane (sachharum officinarum). Int. J. Agric. Biol. (2018); 20: 777-783.
- Arnon DI. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant physiology, (1949); 24: 1.
- Ginnopolitis CN, Rice SK. Superoxide dismutase purification and quantitative relationship with water soluble protein in seedling. Plant Physiol, (1977); 59: 315-318.
- Chance, M, Maehly AC. Assay of catalases and peroxidases. Methods Enzymol, (1955); 2: 764-817.
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry, (1976); 72: 248-254.
- Nakano Y, Asada K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant and cell physiology, (1981); 22: 867-880.
- Bates LS, Waldren RP, Teare ID. Rapid determination of free proline for water-stress studies. Plant and soil, (1973); 39: 205-207.
- Rai MK, Kalia RK, Singh R, Gangola MP, Dhawan AK. 2011. Developing stress tolerant plants through in vitro selection-an overview of the recent progress. Environ. Ex. BoT, (2011); 71:.89-98.
- Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SMA. (2009). Plant drought stress: effects, mechanisms and management. In Sustainable agriculture (pp. 153-188). Springer, Dordrecht.
- Massacci A, Nabiev SM, Pietrosanti L, Nematov SK, Chernikova TN, et al. Response of the photosynthetic apparatus of cotton (Gossypium hirsutum) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging. Plant Physiology and Biochemistry, (2008); 46: 189-195.
- Tahkokorpi M, Taulavuori, K, Laine K, Taulavuori E. After-effects of drought-related winter stress in previous and current year stems of Vaccinium myrtillus L. Environmental and experimental botany, (2007); 61: 85-93.
- Jaleel CA, Manivannan PARAMASIVAM, Wahid A, Farooq M, Al-Juburi HJ, Somasundaram RAMAMURTHY, Panneerselvam R. Drought stress in plants: a review on morphological characteristics and pigments composition. Int. J. Agric. Biol, (2009); 11: 100-105.
- Egert M, Tevini M. Influence of drought on some physiological parameters symptomatic for oxidative stress in leaves of chives (Allium schoenoprasum). Environmental and Experimental Botany, (2002); 48: 43-49.
- Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant physiology and biochemistry, (2010); 48(12), 909-930.
- Osborne CP, Sack L. Evolution of C4 plants: a new hypothesis for an interaction of CO2 and water relations mediated by plant hydraulics. Philosophical Transactions of the Royal Society B: Biological Sciences, (2012); 367: 583-600.
- Pan Y, Wu LJ, Yu ZL. Effect of salt and drought stress on antioxidant enzymes activities and SOD isoenzymes of liquorice (Glycyrrhiza uralensis Fisch). Plant Growth Regulation, (2006); 49: 157-165.
- Wang CQ, Li RC. Enhancement of superoxide dismutase activity in the leaves of white clover (Trifolium repens L.) in response to polyethylene glycol-induced water stress, Acta Physiol. Plant, (2008); 30: 841-847.
- Melchiorre M, Robert G, Trippi V, Racca R, Lascano HR. Superoxide dismutase and glutathione reductase overexpression in wheat protoplast: photooxidative stress tolerance and changes in cellular redox state. Plant growth regulation, (2009); 57: 57.
- Yang Z, Wu Y, Li Y, Ling HQ, Chu C. OsMT1a, a type 1 metallothionein, plays the pivotal role in zinc homeostasis and drought tolerance in rice, Plant Mol. Biol, (2009); 70: 219-229.
- Basu S, Roychoudhury A, Saha PP, Sengupta DN. Differential antioxidative responses of indica rice cultivars to drought stress. Plant Growth Regulation, (2010); 60: 51.
- Smirnoff N. Ascorbate, tocopherol and carotenoids: metabolism, pathway engineering and functions. Antioxidants and reactive oxygen species in plants, (2005); issue no? 53-86.
- Ashraf M, Atha, HR, Harri, PJC, Kwon TR. Some prospective strategies for improving crop salt tolerance. Advances in agronomy, (2008); 97, 45-110.
- DaCosta M, Huang B. Changes in antioxidant enzyme activities and lipid peroxidation for bentgrass species in response to drought stress. Journal of the American Society for Horticultural Science, (2007); 132: 319-326.
- Chagas RM, Silveira JA, Ribeiro RV, Vitorello VA, Carrer H. Photochemical damage and comparative performance of superoxide dismutase and ascorbate peroxidase in sugarcane leaves exposed to paraquat-induced oxidative stress. Pesticide Biochemistry and Physiology, (2008); 90: 181-188.
- Cia MC, Guimaraes ACR, Medici LO, Chabregas SM, Azevedo RA. Antioxidant responses to water deficit by drought‐tolerant and‐sensitive sugarcane varieties. Annals of Applied Biology, (2012); 161: 313-324.
- Boaretto LF, Carvalho G, Borgo L, Creste S, Landell MG, Mazzafera P, Azevedo RA. Water stress reveals differential antioxidant responses of tolerant and non-tolerant sugarcane genotypes. Plant Physiology and Biochemistry, (2014); 74: 165-175.
- Farooq M, Wahid A, Kobayashi N, Fujita DBSMA, Basra SMA. (2009). Plant drought stress: effects, mechanisms and management. In Sustainable agriculture (pp. 153-188). Springer, Dordrecht.
- Anjum S, Tanveer M, Ashraf U, Hussain S, Shahzad B, Khan I, Wang L. Effect of progressive drought stress on growth, leaf gas exchange, and antioxidant production in two maize cultivars. Environmental Science and Pollution Research, (2016); 23: 17132-17141.
- Ashraf MFMR, Foolad M. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental and experimental botany, (2007); 59: 206-216.
- Trovato M, Mattioli R, Costantino P. Multiple roles of proline in plant stress tolerance and development, Rendiconti Lincei (2008); 19: 325-346.
- Chen C, Dickman MB. Proline suppresses apoptosis in the fungal pathogen Colletotrichum trifolii. Proceedings of the National Academy of Sciences, (2005); 102: 3459-3464.
- Abbas SR, Ahmad SD, Sabir SM, Shah AH. Detection of drought tolerant sugarcane genotypes (Saccharum officinarum) using lipid peroxidation, antioxidant activity, glycine-betaine and proline contents. Journal of soil science and plant nutrition, (2014); 14: 233-243.
- Abbas SR, Ahmad SD, Sabir SM, Shah AH. Detection of drought tolerant sugarcane genotypes (Saccharum officinarum) using lipid peroxidation, antioxidant activity, glycine-betaine and proline contents. J. soil scenic. Plant. (2014); 49: 211-219.
- Abbas SR, Ahmad SD, Sabir SM, Shah AH. Detection of drought tolerant sugarcane genotypes (Saccharum officinarum) using lipid peroxidation, antioxidant activity, glycine-betaine and proline contents. Journal of soil science and plant nutrition, (2014); 14(1), 233-243.
- Anjum S, Tanveer M, Ashraf U, Hussain S, Shahzad B, Khan I, Wang L. Effect of progressive drought stress on growth, leaf gas exchange, and antioxidant production in two maize cultivars. Environmental Science and Pollution Research, (2016); 23: 17132-17141.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


