

Review Article
CRISPR/Cas9 system: Current applications and future potential in rice breeding
Anh Phu Nam Bui
Adv. life sci., vol. 7, no. 4, pp. 225-231, August 2020
*- Corresponding Authors: Anh Phu Nam Bui (Email: buiphunamanh@yahoo.com)
Authors' Affiliations
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
Rice (Oryza sativa L.) plays a key role in human social and economic life. In order to meet the increasing needs of human food consumption, there is a constant requirement to develop rice cultivars with enhanced agricultural traits. The emerge of clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated proteins (CRISPR/Cas9) system provides unprecedented opportunities in studying gene functions and creating new rice varieties with better characteristics, including improved tolerance to biotic and abiotic stresses, and increasing yield and quality. This review aims to provide details about the latest results of CRISPR/Cas9 system application on rice to obtain better adapted to environmental and commercial demands.
Keywords: CRISPR/Cas9; Biotic stress; Abiotic stress; Yield; Quality
Introduction![]()
Rice (Oryza sativa L.) is one of the most important stable food and have contended a pivotal role for human life. Because of its main contribution to the world’s energy supply as one of the main daily food for more than half of the human population [1] and its versatile characteristics to multiple environmental conditions [2], rice has been regarded as a strategic crop plant by Food and Agriculture Organization (FAO) [3]. However, rice production is currently facing multiple demands such as climate change, biotic and abiotic stress, and shortage of arable land. As a result, the rice yield is continuously decreasing in recent years. Additionally, rapid human population growth is estimated to expand to 9 billion in 2050, increasing by 34%. Therefore, global rice consumption is projected to rise 45% in the next 30 years, which is equivalent to 450 million tons in 2020 to 650 million ton by 2050 [4,5]. To meet these challenges, enhancing rice resistance to various stress factors and improving rice productivity is a matter of great importance.
In the last few decades, progresses in breeding approaches, especially forward genetic approaches, have played vital roles in elucidating the molecular mechanism that influence agricultural traits of interest in rice. Traditionally, in forward genetic approaches, rice mutations can be mutagenized by chemical (ethyl methane sulfonate-EMS), physical irradiation, (heavy-ion beams or gamma rays) and bacterial genetic insertion (T-DNAs and transposons) [6,7]. However, these methods have limitations. For examples, since EMS and irradiation-generated mutations often lead to mosaicism at the first generation (M1), it is impossible for forward genetic screening to detect any desirable phenotypes at the M1 generation. Similarly, because of the heterozygosity in T0 generation, screening the mutant phenotype caused by T-DNA/transposon has to be conducted at T1 generation. However, with the advent of RNA interference-based screens, only one generation is needed for the identification of causative mutations for mutant phenotypes. Nevertheless, off-targets effect and instability are the main disadvantages of RNA interference-based mutagenesis [8]
Other restrictions of forward genetic approaches also create obstacles in deciphering important crop traits in rice. For instance, constructing a mapping population is often required to narrow down the causal mutations in an EMS mutants or irradiation mutants. For the untagged T-DNA/transposon mutants, map-based cloning is sometimes involved, while for the tagged T-DNA/transposon mutants, identifying the T-DNA-flanked sequences is sometimes difficult due to the truncation or tandem repeats of the T-DNA fragments [8]. Taken together, since rice plants usually take 3 to 6 months from germination to maturity, establishing the relationship between their phenotypes and the genotypes is often laborious and time-consuming.
The newly developed technologies in genome-editing have provided more tools to address the limitations of traditional breeding methods in elaborating functional genomics and crop improvement in rice. These genetic innovations provide more accurate, time-saving, efficient targeted genomic modifications, including whole-gene insertion or deletion, stacking or pyramiding of genes, in a transgene-free manner [9,10]. The principles of genome-editing approaches are the production of double- stranded breaks (DSBs) of targeted DNA and the introduction of cellular repair of DNA damages [11]. The repair mechanism of DSB is predominantly processed through two pathways, i.e. Non-Homologous End-Joining (NHEJ) and Homology-Directed Repair (HDR) [8]. The NHEJ is the error-prone pathway which mainly causes insertions or deletions, thus results in frameshift mutations, or gene knockouts. On the other hand, the HDR pathway is much more precise in repairing by relying on the exchange of the homologous sequences surrounding the DSBs which contributes to the gene replacement. Regardless of the pathways used, DSBs are initiated by site-specific nucleases, including zinc-finger nuclease (ZFNs), transcription activator-like effector nucleases (TALENs) and the recently discovered clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated proteins (CRISPR/Cas9) [12,13]. Although ZFNs and TALENs approach have been successfully applied in molecular breeding in rice, they also have distinctive limitations in plasmid construction and are expensive.
Methods![]()
Literature search strategy and selection criteria
Google Web, Google scholar, NCBI Databases and OMIC Tools were used to obtain data for this review paper. Different key words were used to retrieve the required research articles and bioinformatics-based information, such as “CRISPR/Cas9’’ and “CRISPR/Cas9 in rice’’. Research papers consulted for this review were those published over last 10 to 15 years and information regarding CRISPR/Cas9 application in rice was considered for current review.
Discussion![]()
1. CRISPR/Cas9 – MEDIATED GENOME EDITING SYSTEM
The latest CRISPR/Cas9 is extensively employed in genome editing research thanks to its reliability, efficiency and simplicity [14]. Basically, CRISPR/Cas9 is a RNA- mediated adaptive immune system that can be found in bacteria, and archaea [15]. This immune protection provides resistance against genetic attacks and later stores infection histories in a form of spacer sequences for future safety. These spacers function in concert with Cas9 endonuclease proteins to monitor, recognize and degrade exogenous DNA. This process can be divided in three stages: spacer acquisition, biogenesis and immunity. In the spacer acquisition stage, the foreign DNA is identified, captured and embedded into the CRISPR locus in a form of spacer. Subsequently, the expression of the CRISPR/Cas9 system will be initiated in the biogenesis stage, in which the primary CRISPR-RNAs (crRNAs) is synthesized from the CRISPR locus and subsequently undergone many processes to become crRNAs. Finally, in the immunity stage, the crRNAs, together with the trans-activating RNAs (tracrRNAs), will associate with Cas9 endonuclease, forming a ribonucleotide complex. This complex will initiate interference and consequent degradation of the targeted foreign DNA by base pairing recognition mechanism and endonucleases, respectively.
It was not until the work of Jinek et al. (2012), the significant contribution of CRISPR/Cas9 technology to genome editing begins to emerge by the establishment of the programmable version of CRISPR/Cas9 [16]. This modified version of CRISPR/Cas9 is made up of the customizable single strand RNA (sgRNA), which is the fusion product of crRNA and tracrRNA, the recombinant Cas9 protein and. This combination will result in Cas9/sgRNA complex that targets and initiates DSB at specific DNA sequences. Once DSBs are introduced, NHEJ or HDR strategy is activated to repair the DNA damages, leading to gene knockout, or gene knock-in, respectively [8,17]. CRISPR/Cas9 system has been widely employed in various research model research, including Prokaryotes (Escherichia coli) [18] and Eukaryotes (Saccharomyces cerevisiae, Drosophila melanogaster, Caenorhabditis elegans, Arabidopsis thaliana, etc.) [19-22].
2. CRISPR/Cas9 AND ITS APPLICATION IN RICE IMPROVEMENT
Besides its contribution to human food consumption, rice has been widely employed as excellent model research plant because of its availability of small genome, richness of genetic resources, and genomic synteny with other major crops. As such, there are more than 80 different publications involved in CRISPR/Cas9-mediated genome editing in rice [23]. The results from these papers have shed more lights in understanding the functions of important genes and their potentials. This knowledge has been realized in improving productivity traits, including abiotic stress, herbicide tolerance, disease resistance, yield increase and nutrition enhancement in rice and other agriculture plants.
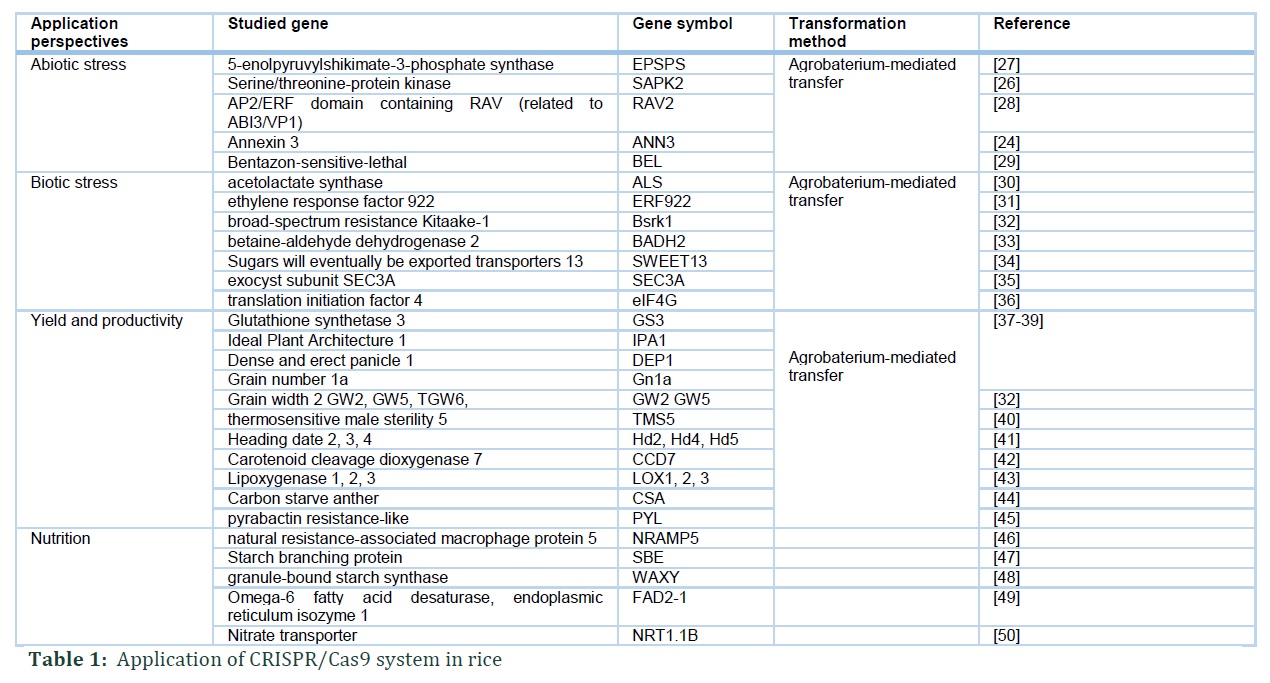
2.1 Abiotic Stress
CRISPR/Cas9 system has been utilized in some reports to elucidate the functions of genes in response to cold, salinity and drought stresses.
2.1.1 Cold Stress
Annexins (OsANN3) are calcium-dependent phospholipid-binding proteins that participate in plant development, various cellular responses including the negative regulation the cold tolerance. The engineered sgRNA targeted and degraded second exon of OsANN3 gene, which subsequently was repaired by NHEJ strategy [24]. Homozygous mutant lines harboring the induced mutations were tested and presented more susceptibility to cold stress than the wild type, illustrating OsANN2 is the negative regulator of cold tolerance-involved mechanism. Similarly, another transcription factor TIFYb participated in cold stress was mutated in CRISPR/Cas9 experiments performed by Huang et al. (2017). Homologous mutant lines indicated the role of TIFYb protein of rice adaptation in low temperature environment [25].
2.1.2 Drought Stress
The phytohormone ABA is known to mediate the responsiveness to many stress processes in plant, including drought. By using CRISPR/Cas9 system to create mutations in third exon of OsSAPK2, the function of OsSAPK2 in the ABA-related drought stress pathway was successfully elaborated. After subjecting the homozygous mutant lines to drought experiments, the results revealed that sapk2 mutants were ABA-sensitive, and exhibited more sensitivity to drought stress compared to the wild type. Taken together, this reveals OsSAPK2 negatively mediates the drought stress in rice [26].
2.1.3 Salinity Stress
The function of transcription factor OsRAV2 protein in saline stress is well-documented. In one study by Li et al. (2020), the authors have stressed on the importance of the promoter region of this gene, GT-1, known for the salt induction. By designing a sgRNA to target this GT-1 element by CRISPR/Cas9 system, after Agrobacterium-mediated transformation, homozygous mutant lines were not able to express OsRAV2 under environment of high salinity. This illustrates the importance of this promoter GT-1 region in saline stress [28].
2.2 Biotic Stress
Disease caused by biotic stress agents, including bacteria, can have harmful influence on rice production. Several genes have been recently mutated by CRISPR/Cas9 system to increase the disease resistance in rice.
2.2.1 Bacterial leaf blight disease
Bacterial leaf blight disease, one of the main threats in rice production, is triggered by the bacteria Xanthomonas oryzae pv. Oryzae (Xoo). When entering the host cells, the pathogen will hijack the cellular resources, including a group of type II TAL (transcription activator-alike), to induce the host’s gene expression to satisfy the pathogen needs. Some of the targets of these TALs are sugars will eventually be exported transporters (SWEET) genes, which encode sugar transporter proteins. In order to test the role of OsSWEET13 in Xoo resistance in rice, sgRNA was engineered to target the first exon of the gene. The mutant lines depicted an increase in Xoo resistance, and less severe symptoms compared to the wild type. This concludes OsSWEET is a promising candidate gene in engineering rice cultivars with low susceptibility to bacterial leaf blight disease [34].
2.2.2 Rice Blast
The most detrimental rice disease is rice blast caused by ascomycetes fungus Magnaporthe oryzae. Rice blast is documented to damage from 10% to 30% loss in rice production and is found widespread in all rice-growing countries. Mountains of evidence, including know-down RNAi expression, portrayed that ethylene responsive factors (ERFs) negatively regulate M. oryzae resistance. Through CRISPR/Cas9 system, one sgRNA was customized to disrupt the first exon of ERF gene. The mutant lines exhibited enhanced resistance to M. oryzae compared to the wild type [31].
In another paper, to explore the function of OsSEC3A in rice blast defense, Ma et al. (2018) engineered two sgRNAs to aim for the third and tenth exon of the gene. The mutant lines demonstrated declined or impaired agricultural traits, including smaller seedlings, shorter main roots, decrease in plant height, panicle length, tiller number, 1000-grain weight and spikelet fertility. However, significant enhancement of M. oryzae resistance was observed. Due to this augmented defense against rice blast, OsSAC3A could be also prominent candidate gene in future research [35].
2.2.3 Rice Tungro Disease
Rice tungro disease is another important rice disease that creates destructive rice production loss in many Asian countries. The interaction of two different viruses, i.e. rice tungro spherical virus (RTSV) and rice tungro bacilliform virus (RTBV), is thought to cause this disease. Traditional breeding approach has illustrated that the translation initiation factor 4 gamma (eIF4G) known to be involved in RTSV resistance. Data from 2018 generated by Macovei et al. (2018) indicated that IR64 mutant line has been generated as an attempt to create new source of RTSV resistance. As a result, these novel eIF4G allele-containing mutant lines can be further studied and developed more diverse RTSV-resistant cultivars [36].
2.2.4 Herbicide Tolerance
Herbicide resistance is another important agricultural trait that has been recently studied and introduced into rice. Acetolactate synthase 1 (ALS1) and 5-enolpyrylshikimate-3-phosphate synthase (EPSPS) are the main target of important herbicides such as chlorsulfuron, bispyribac sodium and glyphosate. While ALS1 is an essential enzyme responsible for biosynthesis branched amino acids, EPSPS participates in the bio-production of auromatic amino acids. CRISPR/Cas9 system has been utilized to mutate ASL1 and EPSPS by customizable gRNAs. Phenotypic screening showed that asl1, and epsps mutant lines were resistant to herbicide and grew normally [27]. Thus, CRISPR/Cas9 system could efficiently generate herbicide resistant rice plants within one generation.
2.3 Yield Improvement
Grain yield is a complex agricultural trait governed by many different genes and environmental factors. Six genes including Gn1a, DEP1, GS3, GS5, GW2, IPA1, and TGW6 are documented to function in grain number, panicle architecture, grain size and plant architecture, respectively. These genes were independently targeted by CRISPR/Cas9 system and the results showed that individual mutant lines harboring their respective edited genes demonstrated increases in grain number, dense erect panicles, and larger grain size [37].
Another line of evidence also reconfirmed the efficiency of CRISPR/Cas system on mutating Gn1a and GS3 [38]. However, only three out of ten novel genotypes lines displayed higher grain number and grain size, suggesting that genetic background could also play a crucial role in masking the expressed mutated genes. Similarly, inducing mutations in three genes GS3, GW2 and Gn1a resulted in higher yields per panicle in two of out three studied varieties, indicating that different cultivars could affect the expression of the CRISPR-Cas9 mediated target genes [39].
Rice heading date (Hd) is another trait controlled by multiple genetic and environmental factors. Three genes Hd2, Hd4 and Hd5 has been revealed as negative regulators of the heading date of rice. Three different sgRNAs were designed to simultaneously disrupt these genes in a single plasmid. After Agrobacterium transformation, homozygous triple mutant lines showed significantly shortened heading date from 5-30 days compared to the wild type [41].
CRISPR/Cas9 system also can be used to facilitate the development of heterosis in hybrid rice. Hybrid rice is a type of rice that has been bred from two very different parents. It can significantly gain better yield other rice varieties. Hybrid rice contributes from 10% to 20% increase in yield over conventional rice; therefore it has a crucial role in global rice production. Thermo-sensitive 5 (TM5) gene was targeted by CRISPR/Cas9 to develop new thermos-sensitive genic male-sterile (TGMS) lines within one year as maintainer lines under permissive conditions. Similarly, carbon starved anther (CAS) gene, which exhibits male sterility and male fertility, depending on short days or long days conditions, respectively. CAS were also mutated to create reverse photoperiod-sensitive genic male-sterile (PGMS) [40]. Taken together, acceleration of TGMS lines breeding can be achieved by CRISPR/Cas9 system and further developed toward large scale application in two-line hybrid rice breeding.
In rice, yield shares overlapping crucial phytohormones and signaling networks with growth. Previous works have revealed the crucial role of the abscisic acid and its receptors in growth control. In order to provide an alternative genetic strategy to improve rice yields, Miao et al. (2018) utilized CRISPR/Cas9 system to target two groups of pyrabactin resistance 1 (PYR1)/PYR1-like (PYL)/regulatory components of (ABA) receptor (RCAR) family proteins, including group I (PYL1-PYL5, and PYL12) and group II (PYL7-PYL11 and PYL13). Although mutants in group II displayed wild-type phenotypes, group I mutants exhibited increased growth and productivity, while maintaining other important traits [45].
2.4 Nutrition Improvement
Rice provides a significant source of daily nutrition for more than half of the world human population. As a result, there is an urgent need to create new rice cultivars enhance the nutrition. Amylose content (AC) and resistant starch (RS) are two of the interest nutrition traits that researchers focus on. AC is arguably the most important quality indicator in rice, and the rice classification based on AC is determined based on the AC. Depend of the consumer demands, the AC and the RS level can be manipulated by the application of CRISPR/Cas9. In the case of reducing AC, AC negative regulator waxy gene were targeted to develop loss of function mutant by CRISPR/Cas9. The homozygous mutant lines waxy showed decreased AC while demonstrated no differences in other agricultural traits compared to wild type, including plant height, grain number per panicle, panicle number per plant, yield per plot, grain width, grain length and 1000-grain weight [48].
In another study, CRISPR/Cas9 has been successfully employed to increase AC and RS by mutating two starch branching enzymes (SBEs), including SBEI and SBEII. While sbeI mutants were not distinguishable from the wild type, sbeII showed significantly higher content in AC and RS, indicating SBEII is a key gene in mediating the nutritional aspect of rice regarding AC and RS [47].
Rice brain oil (RBO) is a commercial derivative product from rice that is highly appreciated in Asian countries as cooking oil. One of the main components in RBO is oleic acid, which will be converted to linoleic acid by fatty acid desaturase 2 (FAD2) gene. Of the four FAD2 gene in rice, FAD2-1 is found highly expressed in rice seed. As a result, the knock-out of FAD2-1 gene by CRIPR/Cas9 system is hypothesized to augment the oleic acid content in rice. A work by Abe et al. (2018) demonstrated that fad2-1 mutants exhibited the two-fold increase of oleic acid content compared to the wild type [49].
Beside improving nutrition by enhancing beneficial factors, decreasing toxic components, such as Cadmium (Cd), is also another aim in rice breeding. Since Cd is highly toxic heavy metal element that poses a serious threat to human consumption, its excessive presence and accumulation in rice need to be resolved. One of the strategy is to mutate the metal transporter gene NRAMP5 using CRISPR/Cas9 system. NRAMP5 regulates the root uptake of Cd, thus knock-outing this gene can reduce the Cd content. Two sgRNAs were engineered to target the ninth exon of NRAMP5. Homozygous nramp5 mutants displayed less Cd quantity and retained wild-type-like traits when grown in Cd-contaminated fields, indicating NRMAP5 is a good candidate gene in development rice cultivars with low Cd content [46].
CRISPR/Cas9 system has emerged as the most effective tool for crop improvement thanks to its accuracy in creating mutations at genes of interest. By generating a genome-wide mutant library, functional characterization of more than 12000 unknown genes can be achieved. Therefore, incorporating beneficial genes in developing elite cultivars for the breeding process not only in rice, but also in other important crops, can be significantly accelerated. Another major advantage of this process is the precision in eliminating the transgene at the first generation through genetic segregation. Consequently, the transgene-free mutant plants, for instance waxy corn, flavored tomato, can be exempted from the biosafety regulation rules by the USDA.
However, there are still some challenges of CRISPR/Cas9 application in rice that researchers need to mitigate. The first challenge is the requirement of tissue culture in introduction of foreign DNA. This can pose difficulties in some commercial varieties, especially indica. Moreover, the complications during tissue culture including labor-insensitivity, tedium, can create somaclonal variations which can compromise the overall of regenerated plants [9]. Second challenge is the disadvantages of the delivery system for CRISPR/Cas9, i.e. Agrobacterium and particle bombardment- mediated transformation. While Agrobacterium-mediate gene transformation is only efficient to limited genotypes within species, particle bombardment- mediated transfer can be productive in a broader range of genotypes. Nevertheless, plant regeneration following bombardment is restricted [9]. The third challenge is the occurrence of undesired off-target events. Although the precision of targeting genes of interest has been enhanced by the introduction of higher specificity endonuclease than Cas9, such as Cpf1 [9,14], gene replacement editing is still difficult because of the low efficiency of HR in plants. The fourth challenge is the field adaptation of the CRISPR/Cas9-mediated genetic modified plants. A majority of the CRISPR/Cas9- related works in rice are only conducted in confined lab conditions. There can be complications once these edited plants are tested on field trials [14]. Therefore, more studies of edited plants on the field can help answer this challenge.
The author would like to thank Ho Chi Minh city Open University for their support during writing this manuscript.
The author declare that there is no competing interests.
References ![]()
- Birla DS, Malik K, Sainger M, Chaudhary D, Jaiwal R, et al. Progress and challenges in improving the nutritional quality of rice (Oryza sativa L.). Critical Reviews in Food Science and Nutrition, (2017); 57(11): 2455-2481.
- Breviario D, Genga A. Stress Response in Rice. J Rice Res, (2013); 2.
- Pérez-Montaño F, Alías-Villegas C, Bellogín RA, del Cerro P, Espuny MR, et al. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiological Research, (2014); 169(5): 325-336.
- Rejesus RM, Mutuc MEM, Yasar M, Lapitan AV, Palis FG, et al. Sending Vietnamese Rice Farmers Back to School: Further Evidence on the Impacts of Farmer Field Schools. Canadian Journal of Agricultural Economics/Revue canadienne d'agroeconomie, (2012); 60(3): 407-426.
- Milovanovic V, Smutka L. Asian Countries in the Global Rice Market. Acta Universitatis Agriculturae et Silviculturae Mendelianae Brunensis, (2017); 65(2): 679-688.
- Wei F-J, Droc G, Guiderdoni E, Hsing Y-IC. International Consortium of Rice Mutagenesis: resources and beyond. Rice (New York, NY), (2013); 6(1): 39-39.
- Li G, Chern M, Jain R, Martin Joel A, Schackwitz Wendy S, et al. Genome-Wide Sequencing of 41 Rice (Oryza sativa L.) Mutated Lines Reveals Diverse Mutations Induced by Fast-Neutron Irradiation. Molecular Plant, (2016); 9(7): 1078-1081.
- Yang N, Wang R, Zhao Y. Revolutionize Genetic Studies and Crop Improvement with High-Throughput and Genome-Scale CRISPR/Cas9 Gene Editing Technology. Molecular plant, (2017); 10(9): 1141-1143.
- Romero FM, Gatica-Arias A. CRISPR/Cas9: Development and Application in Rice Breeding. Rice Science, (2019); 26(5): 265-281.
- Jang G, Lee S, Um TY, Chang SH, Lee HY, et al. Genetic chimerism of CRISPR/Cas9-mediated rice mutants. Plant Biotechnology Reports, (2016); 10(6): 425-435.
- Ran FA, Hsu PD, Wright J, Agarwala V, Scott DA, et al. Genome engineering using the CRISPR-Cas9 system. Nature Protocols, (2013); 8(11): 2281-2308.
- Bi H, Yang B (2017) Chapter Five – Gene Editing With TALEN and CRISPR/Cas in Rice. In: Weeks DP, Yang B, editors. Progress in Molecular Biology and Translational Science: Academic Press. pp. 81-98.
- Char SN, Li R, Yang B (2019) CRISPR/Cas9 for Mutagenesis in Rice. In: Kumar S, Barone P, Smith M, editors. Transgenic Plants: Methods and Protocols. New York, NY: Springer New York. pp. 279-293.
- Mishra R, Joshi RK, Zhao K. Genome Editing in Rice: Recent Advances, Challenges, and Future Implications. Frontiers in Plant Science, (2018); 9(1361).
- Xie K, Yang Y. RNA-Guided Genome Editing in Plants Using a CRISPR–Cas System. Molecular Plant, (2013); 6(6): 1975-1983.
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science (New York, NY), (2012); 337(6096): 816-821.
- Xu R, Wei P, Yang J (2017) Use of CRISPR/Cas Genome Editing Technology for Targeted Mutagenesis in Rice. In: Reeves A, editor. In Vitro Mutagenesis: Methods and Protocols. New York, NY: Springer New York. pp. 33-40.
- Jiang Y, Chen B, Duan C, Sun B, Yang J, et al. Multigene Editing in the Escherichia coli Genome via the CRISPR-Cas9 System. Applied and Environmental Microbiology, (2015); 81(7): 2506-2514.
- Laughery MF, Wyrick JJ. Simple CRISPR-Cas9 Genome Editing in Saccharomyces cerevisiae. Current Protocols in Molecular Biology, (2019); 129(1): e110.
- Gratz SJ, Rubinstein CD, Harrison MM, Wildonger J, O'Connor-Giles KM. CRISPR-Cas9 Genome Editing in Drosophila. Current protocols in molecular biology, (2015); 11131.32.31-31.32.20.
- Dickinson DJ, Goldstein B. CRISPR-Based Methods for Caenorhabditis elegans Genome Engineering. Genetics, (2016); 202(3): 885-901.
- Miki D, Zhang W, Zeng W, Feng Z, Zhu J-K. CRISPR/Cas9-mediated gene targeting in Arabidopsis using sequential transformation. Nature Communications, (2018); 9(1): 1967.
- Hussain A, Imran QM, Yun B-W. CRISPR/Cas9-Mediated Gene Editing in Grain Crops. (2019); Recent Advances in Grain Crops Research.
- Shen C, Que Z, Xia Y, Tang N, Li D, et al. Knock out of the annexin gene OsAnn3 via CRISPR/Cas9-mediated genome editing decreased cold tolerance in rice. Journal of Plant Biology, (2017); 60(6): 539-547.
- Huang QN, Y.F S, X.B Z, L.X S, B.H F, et al. Construction and analysis of tifya and tifyb mutants in rice (Oryza sativa) based on CRISPR/Cas9 technology. J Agric Biotechnol, (2016); 251003-1012.
- Lou D, Wang H, Liang G, Yu D. OsSAPK2 Confers Abscisic Acid Sensitivity and Tolerance to Drought Stress in Rice. Frontiers in plant science, (2017); 8993-993.
- Li J, Meng X, Zong Y, Chen K, Zhang H, et al. Gene replacements and insertions in rice by intron targeting using CRISPR–Cas9. Nature Plants, (2016); 2(10): 16139.
- Li Q, Sapkota M, van der Knaap E. Perspectives of CRISPR/Cas-mediated cis-engineering in horticulture: unlocking the neglected potential for crop improvement. Horticulture Research, (2020); 7(1): 36.
- Xu R, Qin R, Li H, Li D, Li L, et al. Generation of targeted mutant rice using a CRISPR-Cpf1 system. Plant biotechnology journal, (2017); 15(6): 713-717.
- Sun Y, Zhang X, Wu C, He Y, Ma Y, et al. Engineering Herbicide-Resistant Rice Plants through CRISPR/Cas9-Mediated Homologous Recombination of Acetolactate Synthase. Molecular Plant, (2016); 9(4): 628-631.
- Wang F, Wang C, Liu P, Lei C, Hao W, et al. Enhanced Rice Blast Resistance by CRISPR/Cas9-Targeted Mutagenesis of the ERF Transcription Factor Gene OsERF922. PLOS ONE, (2016); 11(4): e0154027.
- Fiaz S, Ahmad S, Noor MA, Wang X, Younas A, et al. Applications of the CRISPR/Cas9 System for Rice Grain Quality Improvement: Perspectives and Opportunities. International journal of molecular sciences, (2019); 20(4): 888.
- Gaoneng S, Lihong X, Guiai J, Xiangjin W, Zhonghua S, et al. CRISPR/CAS9-mediated Editing of the Fragrant Gene Badh2 in Rice. Experimental Techniques, (2017); 31(2): 216.
- Oliva R, Ji C, Atienza-Grande G, Huguet-Tapia JC, Perez-Quintero A, et al. Broad-spectrum resistance to bacterial blight in rice using genome editing. Nature Biotechnology, (2019); 37(11): 1344-1350.
- Ma J, Chen J, Wang M, Ren Y, Wang S, et al. Disruption of OsSEC3A increases the content of salicylic acid and induces plant defense responses in rice. Journal of Experimental Botany, (2017); 69(5): 1051-1064.
- Macovei A, Sevilla NR, Cantos C, Jonson GB, Slamet-Loedin I, et al. Novel alleles of rice eIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to Rice tungro spherical virus. Plant biotechnology journal, (2018); 16(11): 1918-1927.
- Li M, Li X, Zhou Z, Wu P, Fang M, et al. Reassessment of the Four Yield-related Genes Gn1a, DEP1, GS3, and IPA1 in Rice Using a CRISPR/Cas9 System. Frontiers in Plant Science, (2016); 7(377).
- Huang L, Zhang R, Huang G, Li Y, Melaku G, et al. Developing superior alleles of yield genes in rice by artificial mutagenesis using the CRISPR/Cas9 system. The Crop Journal, (2018); 6(5): 475-481.
- Wang Y, Geng L, Yuan M, Wei J, Jin C, et al. Deletion of a target gene in Indica rice via CRISPR/Cas9. Plant Cell Reports, (2017); 36(8): 1333-1343.
- Barman HN, Sheng Z, Fiaz S, Zhong M, Wu Y, et al. Generation of a new thermo-sensitive genic male sterile rice line by targeted mutagenesis of TMS5 gene through CRISPR/Cas9 system. BMC Plant Biology, (2019); 19(1): 109.
- Cui Y, Zhu M, Xu Z, Xu Q. Assessment of the effect of ten heading time genes on reproductive transition and yield components in rice using a CRISPR/Cas9 system. Theoretical and Applied Genetics, (2019); 132(6): 1887-1896.
- Butt H, Jamil M, Wang JY, Al-Babili S, Mahfouz M. Engineering plant architecture via CRISPR/Cas9-mediated alteration of strigolactone biosynthesis. BMC Plant Biology, (2018); 18(1): 174.
- Ma L, Zhu F, Li Z, Zhang J, Li X, et al. TALEN-Based Mutagenesis of Lipoxygenase LOX3 Enhances the Storage Tolerance of Rice (Oryza sativa) Seeds. PLOS ONE, (2015); 10(12): e0143877.
- Gu W, Zhang D, Qi Y, Yuan Z (2019) Generating Photoperiod-Sensitive Genic Male Sterile Rice Lines with CRISPR/Cas9. In: Qi Y, editor. Plant Genome Editing with CRISPR Systems: Methods and Protocols. New York, NY: Springer New York. pp. 97-107.
- Miao C, Xiao L, Hua K, Zou C, Zhao Y, et al. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proceedings of the National Academy of Sciences, (2018); 115(23): 6058-6063.
- Tang L, Mao B, Li Y, Lv Q, Zhang L, et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Scientific Reports, (2017); 7(1): 14438.
- Sun Y, Jiao G, Liu Z, Zhang X, Li J, et al. Generation of High-Amylose Rice through CRISPR/Cas9-Mediated Targeted Mutagenesis of Starch Branching Enzymes. Frontiers in plant science, (2017); 8298-298.
- Zhang J, Zhang H, Botella JR, Zhu J-K. Generation of new glutinous rice by CRISPR/Cas9-targeted mutagenesis of the Waxy gene in elite rice varieties. Journal of integrative plant biology, (2018); 60(5): 369-375.
- Abe K, Araki E, Suzuki Y, Toki S, Saika H. Production of high oleic/low linoleic rice by genome editing. Plant Physiology and Biochemistry, (2018); 13158-62.
- Li J, Zhang X, Sun Y, Zhang J, Du W, et al. Efficient allelic replacement in rice by gene editing: A case study of the NRT1.1B gene. Journal of Integrative Plant Biology, (2018); 60(7): 536-540.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


