

Review Article
Role of MicroRNA in Endometrial Carcinoma
Zeeshan Javed, Mukhtar Ullah*, Hafiz Ahsan Ashfaq, Afzaal Hussain Shah, Muhammad Shahzad, Muhammad Bilal, Aleena Sumrin, Hamid Bashir, Muhammad Hassan Siddiqi, Haleema Sadia
Adv. life sci., vol. 4, no. 1, pp. 08-13, November 2016
*- Corresponding Author: Mukhtar Ullah (Email: mukhtar.camb@gmail.com)
Authors' Affiliations
Abstract![]()
Introduction
Methods
Discussion
References
Abstract
Endometrial carcinoma (EC) is a hall mark of gynecological malignancies that usually affects women above the age 50. It is one of the major causes of mortality in females with ever increasing prevalence and the mortality rate is 1.7 to 2.4 per 100000 and each year 10000 death occur due to ECs. MiRNAs regulate the expression of different proto-oncogenes and signaling pathways that are directly or indirectly involved in the development of cancer. Different miRNAs i.e. (miRNA-449, miRNA 370, miRNA-424, and miRNA-152) which gets up or down regulated during endometrial cancer are the potential biomarkers for early diagnosis of EC. Targeting this relationship between the miRNA and signaling pathways may help in the development of new treatment in endometrial cancer. In current study, we reviewed literature from PubMed using miRNA and endometrial cancer as keywords and outlined the synthesis of potent miRNA and role of different miRNAs involved in ECs. The study revealed different sub types of miRNA played crucial role in the development of cancer by up and down regulation of different metabolic pathways. Many evidences have supported that miRNAs play role in control and regulation of different pathways leading to cancer and targeting these pathways may bring changes in the diagnosis as well as in treatment of EC.
Keywords: miRNA Biogenesis, Endometrium, Cancer
Introduction
Endometrial cancer (EC) affects 75% of women with post-menopausal [1,2]. It is highly prevalent and nearly found in 287,000 people worldwide [3]. Endometrial carcinoma can be classified in to two subtypes i.e. Endometriod endometrial carcinoma (Type-I) and non-Endometriod endometrial carcinoma (Type-II) [4]. Type-I EC arises from eminent increase in the amount of circulating estrogen which is common in women after menopause. Broadly, type-I ECs can be distinguished on the basis of cytological examinations in to three major grades (G1 ≤ 6%, G2 is 5-50 and G3 > 50%) each with its own diagnostic features that are referred to as bench mark in diagnosis of Type-I ECs [5].
An elevated estrogen level down regulates the expression of phosphatase and tensin homolog PTEN that in turn initiate a plethora of molecular signaling pathways which facilitates transcription of oncogenic Kirsten rat sarcoma (K-ras). K-ras in turn accelerates the phosphorylation of estrogen receptor via activation of beta-catenin through increased level of Mitogen-activated protein kinases (MAPK). This altered signaling drastically enhances cellular proliferation of endometrial tissue that leads to cancer. On the other hand, Type-II ECs are far more different from the Type-I. As they are generally characterized after passing several stages of metastasis and are independent of estrogen levels [6]. Type-II ECs can be classified in to serous, clear cell differentiation or undifferentiated tumors [7]. Serous carcinoma falls under the category of Type-II ECs and are characterized by dyshesive polygonal cells that have high ratio of granules in cytoplasm. They are generally referred to as papillary tumors that affect females with age more or less five years greater than those suffering from Type-I ECs [8]. Clear cell differentiated endometrial carcinoma is a rare pathology that has characteristics similar to papillary tumors but differ by the presence of clear cytoplasm. Undifferentiated tumors are also termed as Carcinosarcoma that are quite prevalent among postmenopausal women [9]. They are diagnosed on the basis of mesenchymal components. Major therapy against this massive malignancy includes surgery which primarily involves excision of ovaries and other reproductive organs and other forms of treatment include adjuvant therapies that involve chemotherapy, radio therapy and hormonal therapies. These therapies have promising benefits yet the rate of recovery is quite often very slow and rate of relapse and recurrences is far greater then preventive cure [10].
The size of miRNAs are usually 10-25 base pair and have been confirmed to play an extensive role in cancer initiation, progression and metastasis [11]. Several of evidences have revealed the bifacial aspect of miRNAs in tumor environment. On the basis of their involvement in B-cell lymphoma, He et al demonstrated first time the oncogenic role of miRNAs. Their findings threw limelight on involvement of miRNA 17 polycistronic cluster in tumor progression, accelerated levels of miRNAs resulted in the hyper-expression of c-Myc causing tumor in induced B-cell Lymphoma mouse model. Furthermore, they set the foundation of classification of microRNAs in to onco-miRNAs and tumor suppressor miRNAs [12,13]. On these grounds it can be established that several miRNAs can act as oncogenic or tumor suppressor. However, numerous publications have implicated miRNAs as being suppressed in majority of cancer related pathologies [14-16]. Owing to this global perception, scientists worldwide tried to investigate the role of miRNAs processing machinery (RNase III drosha and dicer in development of cancer. The RNase III drosha initiate the processes of miRNA synthesis where the dicer cleave the double strand RNA and miRNA in to short fragments [17-19]. Kumar et al conducted studies on mouse model of lung cancer, suggesting that miRNAs processing machinery has a pivotal role in tumor differentiation and metastasis, absence of any one of these can lead to preferential tumor growth in mice lacking either Drosha or Dicer1 [16].
Methods
Search Strategy and Literature Selection Criteria
In this study we searched data in PubMed, and PubMed Central by using keywords microRNA and endometrial cancer. This review primarily focused on synthesis of miRNAs and their involvement in EC.
Discussion
MiRNAs Biogenesis
MiRNA biogenesis is regulated by TGF-β signaling pathway. In TGF- β mediated pathway the ligand bind with the TGF-β receptor which activate the R-Smads complex with common smads (Co-Smad) and transloccated in to nucleus and alter the miRNA transcription [20]. Most of the miRNA genes transcribed as single or cluster from the intronic region of DNA. The newly synthesized pre-miRNA then go through further processing by micro processing complex that contain RNAse III enzyme Drosha and a RNA binding protein DGCR8 ( DiGeorge Syndrome critical region 8) that makes this molecules in to double stranded pre-miRNA, symbolized by its hairpin loop structure [21]. DROSHA contain two domain for RNAse III that consciously remove the long structure of pre-miRNA [22]. Then the Exportin protein (Xpo5) transport the pre-miRNA from the nucleus. These proteins carry two NLS (Nuclear Localizing Sequences) that helps in the transportation of pre-miRNA. The miRNA entering to the cytoplasm is further processed by another RNAse III enzyme called DICER1. The activity of Dicer1 is to transform pre-miRNA in to mature miRNA consisting of leading strand [23]. The mature miRNA is then brimmed into the RNA induced complex that already carries Transactivation Responsive RNA Binding Protein (TRBP) and Arguanate proteins (AGO) [24]. The exact mechanism of TRBP is not well known but there are different views about the mechanism of the action of TRBP and AGO, the most appraisal view suggests that the TRBPs orchestrate the physiological binding of various types of AGO proteins with Dicer. The AGO mediate the cleave of the hairpin to yield ~23 nucleotide mature miRNA and the whole activity in turns in promote guiding strand to bind to the target [25,26].
MiRNA silence the expression of target mRNA by degrading mRNA poly-A tail in the presence of GW-182 and AGO-associating protein. Perturbed processing can lead to tumor progression and eventually cancer. There are certain steps in the microRNA processing which are considered to be pinnacle in maintaining normal gene silencing carried out by miRNAs. There are a number of factors which must be taken into consideration like genomic alterations, oncogenic factors and epigenetic modifications of histone proteins that alter normal processing of miRNAs and leads to aberrant expression of downstream targets of miRNAs to produce tumor [27]. Piling evidences have suggested the involvement of aberrant miRNA signaling in diverse gynecological malignancies strengthening the fact that miRNAs are produced as pre miRNAs molecules which are trimmed, cleaved and transformed into primary miRNA which are exported out of the cell and mature in to mature miRNA that with the aid of TRBP and AGO undermines the process of gene silencing. miRNAs serve as oncogenic or cancer suppressor depending upon their level of expression and concentration [28-30].
Subtypes of miRNA and their Potential Role in EC
MiRNAs have substantial role as therapeutic target in ECs. Extensive research has been conducted to understand the multidimensional benefits of miRNAs in different cancer related pathologies which can aid in their diagnosis, prognosis and treatment with the recent advancements in the field of bioinformatics and molecular biology, myriads of opportunities have emerged to delineate therapeutic and structural prose of miRNAs.
Jurcevic et al. studied the expression of miRNA in endometrial cancer and normal endometrial samples and found that there are 138 different miRNAs which are differential expressed in EC. This analysis of differential expression of miRNA will help in classification and prognosis of the ECs [31]. A list of several types of miRNA that are differential expressed in ECs and their associated pathway are given at the end. miRNA-101 and its potential role in ECs was investigated by Konno et al and reported that this inhibits the proliferation and induces apoptosis in EC cells. It also initiate the migration, and invasion of epithelial mesenchymal transition associated cancer cell migration and invasion, renege the sphere forming capacity and increase the chemosensitivity toward paclitaxel drug. MiRNA-101 potentially target the expression of EZH2, MCL-1 and FOS and thus inhibit the cell proliferation, invasion and self-renewal in endometrial cancer by silencing of multiple oncogenes. The studies suggested that miRNA-101 and EZH2/MCL-1/FOS are the potential target for therapeutic development [32].
A study carried by Kaur et al using disease type model investigated the effects of epigenetic modifications on miRNAs expression profile. Investigation establish the crucial involvement of methylation in prognosis of early tumor differentiation, moreover methylation of miRNA-129b is inevitable for early endometrial tumorgenesis [33].
In some studies it is reported that there are imbalance in the endometrial cell proliferation by unimpeded estrogen and mismatch repair (MMR) system. Banno et al showed that the both types of EC are due to methylation and mutation in different genes. In Type-I EC, hypermethylation of MMR gene hMLH1 and mutation in PTEN, β-catenin, K-ras gene while in Type-II EC HER-2/neu and p53 are mutated and hypermethylation of SPRY2, RASSFIA, RSK4, CHFR and CDH1 and also methylation of some tumor silencing microRNA including miRNA-124, miRNA-126, miRNA-137, miRNA-129-2, miRNA-152 and miRNA-491. The study suggested that both genetic and epigenetic mechanisms are involved in endometrial cancer. Mutation and methylation of MMR gene lead to activate different proto-oncogenes and mutation and methylation of MMR gene in germ cell which might lead to inherited and familial cancer. The study also suggested that targeting the MMR gene which might help in development of new treatment and diagnostic strategies [34]. Zhang et al, using EC Ishikawa cells, demonstrated that miRNA-205 down regulates the PTEN expression by binding to the 3ꞌ UTR region of PTEN, which leads to the inhibition of cellular apoptosis in endometrial cancer cells. The studies suggested that targeting this relationship between miRNA-205 and PTEN might be helpful in therapeutic strategies of EC as well [35]. miRNA-424, a tumor suppressor miRNA has been studied extensively for its diagnostic benefits. Owing to its high expression in ECs it has been estimated to have a critical role in the process of tumor suppression. miRNA-424 directly mediates tumor suppression in ECs by directly binding to G-protein couple estrogen receptor (GPER) resulting in activation Akt/PI3K rendering it passive to bind to the PTEN domain. This could aid in designing targeted therapy against ECs [36].
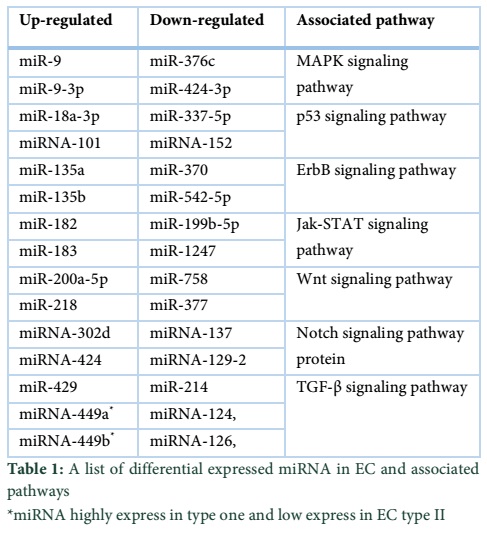
The endogenous miRNA-234 also plays an important role in the EC suppression. miRNA-234 inhibits the proliferation and migration of EC stem cells by down regulation of protein O-glucosyltransferase expression and Notch signaling pathway protein. Gao et al also reported that the expression level of miRNA-234 genomic imprinted miRNA cluster between EC stem cell (HuECSC) and human endometrial cancer cell were significantly different from each other. Their findings have clearly delineate the fact that miRNA-234 presence is mandatory for the development and maturation of endometrial cells and thus can be used as diagnostic marker while estimating the extent of ECs [37].
Jang et al, using miRNA microarray to check the microRNA expression level in clear cell carcinomas, serous carcinomas and germ cell tumor, shed light on the differential expression of six miRNAs. They found that all three types of cancer had a varied amount of miRNA expression. However, miRNA-449 was under-expressed in ovarian and uterine clear cell carcinomas, while the miRNA-302d was highly expressed in germ cell tumor suggesting the fact miRNA-449 has a suppressive role in all of classified tumors while miRNA-302 aggravates the intensity of tumor [38]. In similar context Chen et al study the role of miRNA-370 in endometrial ovarian cancer. Their findings have suggested that miRNA-370 in IGROVI and TOV112D endometrial ovarian cancer cells inhibit the cellular viability and colony formation and increase the chemo sensitivity of endometrial ovarian cancer cell toward cDDP. Furthermore, they suggested that miRNA-370 was also involved in the negative regulation of Endoglin (ENG). Owing to these ground breaking realities they concluded that the hypermethylation was the potential mechanism of miRNA epigenetic silencing [39,40].
In conclusion of the discussion, EC is one of the major causes of death in women due to limited diagnosis and treatment facility. Majority of women suffering from this disease die each year due to poor prognosis and disease diagnosis. MiRNAs have shown to be a leading regulator of oncogenes as well as their expression. Tones of evidence that peep deep in to miRNAs functioning in cancer have revealed that miRNAs have a momentous role in tumor progression and inhibition. Some of the miRNA epigenetically silence the mismatch repair gene (MMR gene) that lead to inherited cancer. So role of miRNA is very important for the development of EC. Different sub types of miRNA were studied and each has played an important role in suppression and induction of tumor. Upgrading the accessibility of tumor silencer miRNAs is likewise a noteworthy hindrance that should be overcome and in accordance with this methodology, different Nano innovative systems must be utilized to enhance bioavailability and uptake of miRNAs at the objective destinations. The distributed date on miRNA in malignancy has extended at a momentous rate and their connections with infection firstly portrayed in 2002 and soon get their significance as biomarker for forecast, conclusion and anticipation of various ailments. Point by point and exhaustive pre-clinical studies will enhance our comprehension of the still understudied parts of miRNAs in various diseases and subtypes. A further insight into miRNA profile can unfold and expand views about EC and can set new foundations in the disease diagnosis and cure.
Reference
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. A Cancer Journal for Clinicians, (2016); 66(1): 7-30.
- Tsikouras P, Bouchlariotou S, Vrachnis N, Dafopoulos A, Galazios G, et al. Endometrial cancer: molecular and therapeutic aspects. European Journal of Obstetrics & Gynecology and Reproductive Biology, (2013); 169(1): 1-9.
- Jemal A, Bray F, Center MM, Ferlay J, Ward E, et al. Global cancer statistics. A Cancer Journal for Clinicians, (2011); 61(2): 69-90.
- Rose PG. Endometrial carcinoma. New England Journal of Medicine, (1996); 335(9): 640-649.
- Mutter GL, Lin MC, Fitzgerald JT, Kum JB, Baak JP, et al. Altered PTEN expression as a diagnostic marker for the earliest endometrial precancers. Journal of the National Cancer Institute, (2000); 92(11): 924-930.
- Leslie KK, Thiel KW, Goodheart MJ, De Geest K, Jia Y, et al. Endometrial cancer. Obstetrics & Gynecology Clinics of North America, (2012); 39(2): 255-268.
- Cavanagh D, Fiorica JV, Hoffman MS, Durfee J, Nicosia SV. Adenocarcinoma of the Endometrium: An Institutional Review. Cancer Control, (1999); 6(4): 354-360.
- Ryan AJ, Susil B, Jobling TW, Oehler MK. Endometrial cancer. Cell Tissue Research, (2005); 322(1): 53-61.
- Zagouri F, Bozas G, Kafantari E, Tsiatas M, Nikitas N, et al. Endometrial cancer: what is new in adjuvant and molecularly targeted therapy? Obstetrics and Gynecology International, (2010); 749579.
- Secord AA, Havrilesky LJ, O'Malley DM, Bae-Jump V, Fleming ND, et al. A multicenter evaluation of sequential multimodality therapy and clinical outcome for the treatment of advanced endometrial cancer. Gynecologic Oncology (2009); 114(3): 442-447.
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell, (1993); 75(5): 843-854.
- He L, Thomson JM, Hemann MT, Hernando-Monge E, Mu D, et al. A microRNA polycistron as a potential human oncogene. Nature, (2005); 435(7043): 828-833.
- Calin GA, Croce CM. MicroRNA signatures in human cancers. Nature Reviews Cancer, (2006); 6(11): 857-866.
- Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, et al. MicroRNA expression profiles classify human cancers. Nature, (2005); 435(7043): 834-838.
- Thomson JM, Newman M, Parker JS, Morin-Kensicki EM, Wright T, et al. Extensive post-transcriptional regulation of microRNAs and its implications for cancer. Genes & Development, (2006); 20(16): 2202-2207.
- Kumar MS, Erkeland SJ, Pester RE, Chen CY, Ebert MS, et al. Suppression of non-small cell lung tumor development by the let-7 microRNA family. Proceedings of National Academy of Sciences USA, (2008); 105(10): 3903-3908.
- Karube Y, Tanaka H, Osada H, Tomida S, Tatematsu Y, et al. Reduced expression of Dicer associated with poor prognosis in lung cancer patients. Cancer Science, (2005); 96(2): 111-115.
- Lin RJ, Lin YC, Chen J, Kuo HH, Chen YY, et al. microRNA signature and expression of Dicer and Drosha can predict prognosis and delineate risk groups in neuroblastoma. Cancer Research, (2010); 70(20): 7841-7850.
- Merritt WM, Lin YG, Han LY, Kamat AA, Spannuth WA, et al. Dicer, Drosha, and outcomes in patients with ovarian cancer. New England Journal of Medicine, (2008); 359(25): 2641-2650.
- Guo L, Zhang Y, Zhang L, Huang F, Li J, et al. MicroRNAs, TGF-beta signaling, and the inflammatory microenvironment in cancer. Tumor Biology, (2016); 37(1): 115-125.
- Rakheja D, Chen KS, Liu Y, Shukla AA, Schmid V, et al. Somatic mutations in DROSHA and DICER1 impair microRNA biogenesis through distinct mechanisms in Wilms tumours. Nature Communications, (2014); 24802.
- egert J, Ishaque N, Vardapour R, Georg C, Gu Z, et al. Mutations in the SIX1/2 pathway and the DROSHA/DGCR8 miRNA microprocessor complex underlie high-risk blastemal type Wilms tumors. Cancer Cell, (2015); 27(2): 298-311.
- Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nature Reviews Genetics, (2008); 9(2): 102-114.
- Yi R, Qin Y, Macara IG, Cullen BR. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes & Development, (2003); 17(24): 3011-3016.
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature, (2001); 409(6818): 363-366.
- Chendrimada TP, Gregory RI, Kumaraswamy E, Norman J, Cooch N, et al. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature, (2005); 436(7051): 740-744.
- Lee HY, Doudna JA. TRBP alters human precursor microRNA processing in vitro. RNA, (2012); 18(11): 2012-2019.
- Zhang L, Huang J, Yang N, Greshock J, Megraw MS, et al. microRNAs exhibit high frequency genomic alterations in human cancer. Proceedings of National Academy of Sciences USA, (2006); 103(24): 9136-9141.
- Cimmino A, Calin GA, Fabbri M, Iorio MV, Ferracin M, et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proceedings of National Academy of Sciences USA, (2005); 102(39): 13944-13949.
- Raver-Shapira N, Marciano E, Meiri E, Spector Y, Rosenfeld N, et al. Transcriptional activation of miR-34a contributes to p53-mediated apoptosis. Molecular Cell, (2007); 26(5): 731-743.
- Jurcevic S, Olsson B, Klinga-Levan K. MicroRNA expression in human endometrial adenocarcinoma. Cancer Cell International, (2014); 14(1): 88.
- Konno Y, Dong P, Xiong Y, Suzuki F, Lu J, et al. MicroRNA-101 targets EZH2, MCL-1 and FOS to suppress proliferation, invasion and stem cell-like phenotype of aggressive endometrial cancer cells. Oncotarget, (2014); 5(15): 6049-6062.
- Kaur S, Lotsari JE, Al-Sohaily S, Warusavitarne J, Kohonen-Corish MR, et al. Identification of subgroup-specific miRNA patterns by epigenetic profiling of sporadic and Lynch syndrome-associated colorectal and endometrial carcinoma. Clinical Epigenetics, (2015); 7(1): 20.
- Banno K, Yanokura M, Iida M, Masuda K, Aoki D. Carcinogenic mechanisms of endometrial cancer: involvement of genetics and epigenetics. Journal of Obstetrics and Gynaecology Research, (2014); 40(8): 1957-1967.
- Zhang G, Hou X, Li Y, Zhao M. MiR-205 inhibits cell apoptosis by targeting phosphatase and tensin homolog deleted on chromosome ten in endometrial cancer Ishikawa cells. BMC Cancer, (2014); 14: 440.
- Zhang H, Wang X, Chen Z, Wang W. MicroRNA-424 suppresses estradiol-induced cell proliferation via targeting GPER in endometrial cancer cells. Cellular and Molecular Biology (Noisy-le-grand), (2015); 61(7): 96-101.
- Gao Y, Liu T, Huang Y. MicroRNA-134 suppresses endometrial cancer stem cells by targeting POGLUT1 and Notch pathway proteins. FEBS Letters, (2015); 589(2): 207-214.
- Jang SG, Yoo CW, Park SY, Kang S, Kim HK. Low expression of miR-449 in gynecologic clear cell carcinoma. International Journal of Gynecological Cancer, (2014); 24(9): 1558-1563.
- Chen XP, Chen YG, Lan JY, Shen ZJ. MicroRNA-370 suppresses proliferation and promotes endometrioid ovarian cancer chemosensitivity to cDDP by negatively regulating ENG. Cancer Letters, (2014); 353(2): 201-210
- Ye W, Xue J, Zhang Q, Li F, Zhang W, et al. MiR-449a functions as a tumor suppressor in endometrial cancer by targeting CDC25A. Oncology Reports, (2014); 32(3): 1193-1199.

