

![]()
Assessment of salinity tolerance in rice using seedling based morpho-physiological indices
Syed Adeel Zafar1*, Sajid Shokat2, Hafiz Ghulam Muhu-Din Ahmed1, Adeel Khan1, Muhammad Zeshan Ali3, Rana Muhammad Atif1¥
Adv. life sci., vol. 2, no. 4, pp. 142-149, August 2015
*- Corresponding Author: Syed Adeel Zafar (Email: adeelzafarpbg@gmail.com)
Author Affiliations[Date Received: 25/05/2015; Date Revised: 10/08/2015; Date Published Online: 25/08/2015]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Salinity is among the most damaging abiotic stresses for rice production which limits its growing area. The present research was conducted to evaluate five rice varieties for salinity tolerance at seedling stage.
Methods: Experiment was conducted in triplicate and in two sets. One set was grown as a control (non-stress) and other as salt stressed. Salt stress of 15 dS/m was applied to one set of rice seedlings under controlled conditions. Data for different growth related morpho-physiological traits, i.e. germination percentage, root and shoot length, seedling fresh and dry weight, Na+ and K+ uptake were recorded after 15 days of seedling emergence under control as well as salinity condition.
Results: Significant differences were observed among the genotypes under both the treatments and interaction of the evaluated traits suggested a significant variability among the rice genotypes under salt stress. NIAB-IRRI-9, Basmati-198 and KSK-133 were proved to be relatively salt tolerant varieties as they showed good performance for the recorded parameters. However, Basmati-385 was observed a salt sensitive variety due to highest reduction in seedling fresh and dry weight along with the maximum Na+ uptake.
Conclusion: Based on obtained results, it was concluded that the evaluated morpho-physiological traits were useful to screen rice cultivars for salinity stress. In addition, NIAB-IRRI-9, Basmati-198 and KSK-133 can be used in breeding programs as tolerant check and Basmati-385 can be used as sensitive check.
Keywords: Rice, salinity stress, Basmati, morphological traits
Introduction
Rice is an imperative cereal crop of Pakistan and cultivated on an area of 2.891 million hectares with a production of 7.005 million tonnes and average yield of 2,423 Kg/Ha [1]. In Pakistan, rice is the second largest cereal food crop and a major export commodity. Its exports earned foreign exchange of US$ 1.53 billion during July-March 2014-15 [2].
Salinity is amongst the foremost challenges faced by agriculture [3]. More than 30% of arable land is affected by salts stress that limits the distribution and productivity of crops [4,5]. High salinity injects Na+ or Cl- in plants and causes necrosis and burning of leaf tips [6]. Salinity limits the germination of seed, survival of seedlings and giving rise to uneconomical crop stand [7]. Higher level of salinity reduces the shoot weight, number of leaves, plant height, root length and surface area of roots [8,9].
Rice is a salinity sensitive crop, but the level of salinity tolerance varies in different varieties of rice [10]. The extent of salinity varies depending upon the plant growth stage. In most of the cultivars, more damage is done at germination and early growth stages [11], indicating that salinity has negative effect on plant early growth stages. It also affects the germination time, root index, root and shoot length [12,13].
In salt stress, sodium ions enter the plant cell and causes ionic stress to plant [14,15]. Since plants cannot move from one place to another to mitigate salt stress so nature has gifted them with different mechanisms of salinity tolerance. Na+ to K+ ratio confers salt tolerance in rice [16]. Under saline conditions when there is a higher level of Na+ present in the root zone, plant alters its ion uptake and tolerant genotypes maintain a lower ratio of Na+ to Ca+, Mg+ and high level of K+ while susceptible genotypes could not maintain it [17]. Screening on the basis of these parameters can provide plant material that has the ability to tolerate stress and give better yield under saline conditions.
Primary objective of this study was to assess the response of different growth parameters in five rice varieties against salt stress. Secondary objective of this study was to identify salt tolerant rice varieties on the basis of seedling based morpho-physiological characters.
Methods
Five genotypes of Basmati rice were sown in the greenhouse of Department of Plant Breeding & Genetics, University of Agriculture, Faisalabad, Pakistan during the growing season of 2013-2014 to find out the effect of salinity stress on various growth related morphological parameters of rice.
Mean minimum and maximum temperature in the green house was maintained by electric heaters during day and night time at 25-32°C respectively. Seeds of two sets were grown in plastic pots filled with sand in the completely randomized design (CRD) in triplicate (15 seeds per replication). Salt stress of 15 dS/m was applied to one set of pots. The other set was treated as control.
The genetic material, i.e. NIAB-IRRI-9, KSK-133, Super Basmati, Basmati-185 and Basmati-198 used in research was collected from Department of Plant Breeding & Genetics, University of Agriculture Faisalabad, Pakistan. Seedling growth response of 15 days old seedlings was assessed on the basis of different growth related traits, including germination percentage, root and shoot lengths, seedling fresh and dry weight, dry matter percentage, Na+ and K+ uptake. Root length (cm) was measured from the root-shoot joint to the end of root tip with the help of measuring tape. The average data of different parameters were calculated. Shoot length (cm) was measured from the root-shoot joint to the upper tip of leaf with the help of measuring tape. Root-Shoot length ratio was computed by dividing the root length by shoot length. Seedling fresh weight (SFW) was recorded immediately after harvesting to avoid evaporation. For dry weight (g) estimations, pre-weighted seedlings were kept at 90°C in brown paper till complete drying. Seedling dry weight (SDW) was measured after complete drying. Na+ and K+ concentration (mg g-1 dry weight) of shoot was determined from a 0.5 g dried digested sample using a flame photometer (Jenway PFP-7). The experiment was conducted under completely randomized fashion with three repeats. The differences between genotypes for the recorded characters were tested for significance using standard techniques of Analysis of variance (ANOVA) by using Statistix 8.1 program. Tuckey’s test at 5% level of probability was used to test the differences among mean values [18].
Results
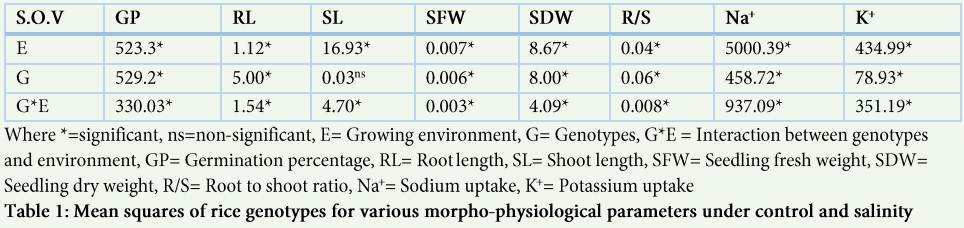
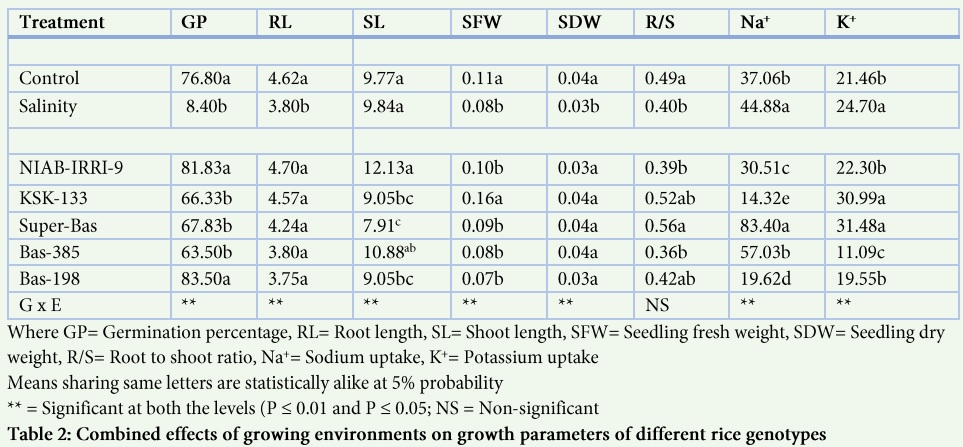
Mean squares extracted from Analysis of variance at seedling depicted considerable range and variability for all the studied traits among the five rice varieties grown under control and saline conditions (Table 1). The statistical analysis also showed significant differences for genotype x environment interactions for different recorded parameters (Table 1). Germination percentage was decreased under salt stress in the genotypes NIAB-IRRI-9, KSK-133 and Basmati-385 while it was high in Super Basmati and Basmati-198 (Fig. 1A). Highest reduction in germination percentage was observed in the genotype KSK-133 (34%) followed by Basmati-385 (27.67%) under salt stress while a little reduction in germination percentage was observed in NIAB-IRRI-9 (5%).
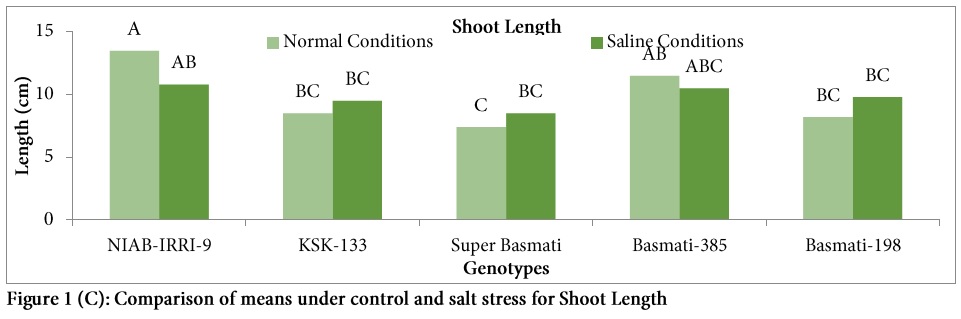
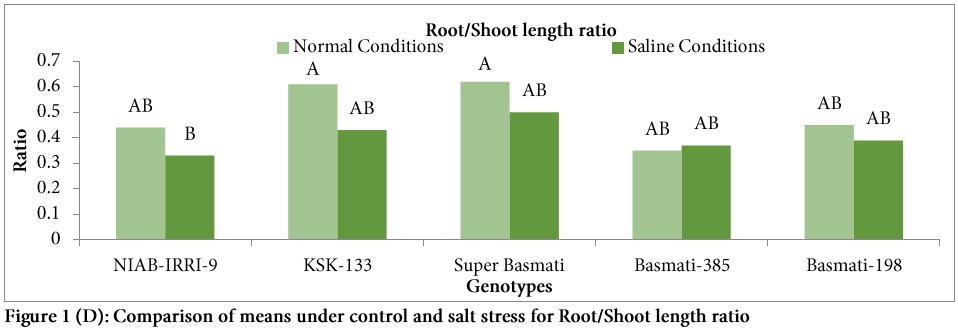
Root length was generally reduced under salinity stress in all genotypes except Basmati-198 (Fig. 1B). Highest percent reduction in root length was observed in genotype NIAB-IRRI-9 (40.66%) under salt stress followed by KSK-133 (25%) and highest root length (5.9 cm) was also observed in the same genotype NIAB-IRRI-9 under control (Fig. 1B). Similarly, significant differences were observed among the genotypes for shoot length (Fig. 1). Highest shoot length was observed in the genotype NIAB-IRRI-9 under control (13.5 cm) and saline conditions (10.8 cm) while lowest shoot length was observed in Super Basmati under control (7.4 cm) and saline conditions (10.5 cm). In the present study, significant variation was observed among the genotypes for root/shoot ratio under salt stress as compared to control (Table 2). Root/shoot ratio was decreased under salt stress in all genotypes except Basmati-385 (sensitive variety). Highest root/shoot ratio under control condition was observed in Super Basmati (0.62) followed by KSK-133 (0.61).
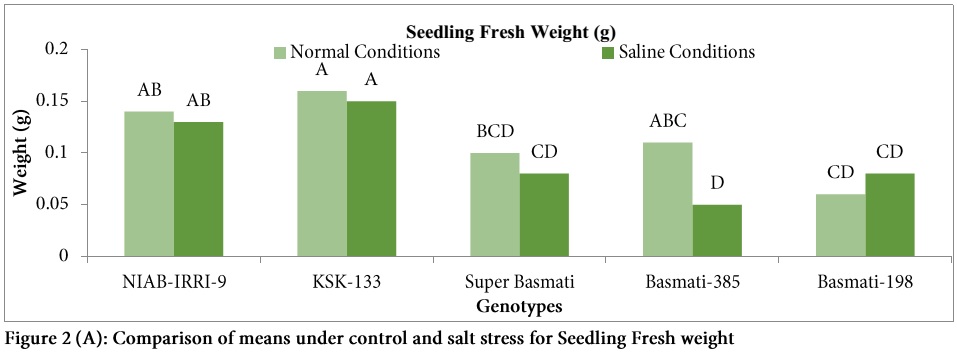
SFW was reduced in all genotypes under salt stress except Basmati-198 (Fig. 2A) and statistically significant differences were observed for fresh weight under salt stress as compared to control (Table 1).
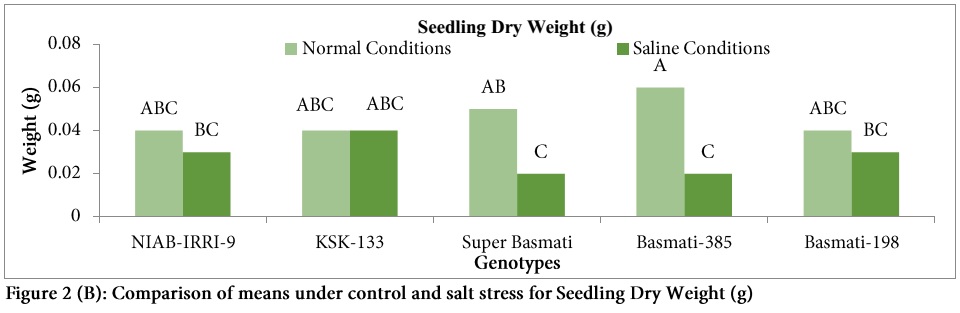
Highest reduction in SFW under salt stress was observed in genotype Basmati-385 (54.5%) followed by Super Basmati (20%). Seedling dry weight was reduced under salt stress in all genotypes except KSK-133 (Figure 2) and significant variation was observed among all the genotypes (Table: 1). Highest SDW was observed in Basmati-385 (0.06 g) under control conditions. Highest reduction (66.66%) in SDW was also observed in the genotype Basmati-385followed by Super Basmati (60%) under salt stress.
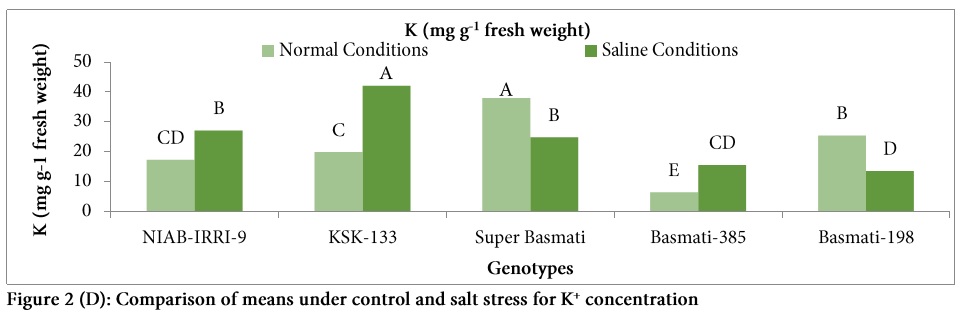
Significant differences were observed among the genotypes for Na+ uptake under salt stress (Table: 1, Fig. 2). Highest Na+ concentration was observed in the genotype Basmati-385 and it was 142% higher under salt stress as compared to control while lowest Na+ uptake was observed in the genotype KSK-133 under saline conditions. In the current study, significant variation for K+ uptake was observed in all genotypes (Table: 1). K+ uptake was increased under salt stress in the genotypes NIAB-IRRI-9, KSK-133 and Basmati-385 while it decreased in Super Basmati and Basmati-198 (Fig. 2).
Tables and Figures



Discussion
Agriculture in Pakistan is facing multiple problems regarding yield and other factors [19-22]. Different strategies are also showing up against those problems [23-25]. Salinity has been one of the major abiotic stress in agriculture and limiting crop growing area and production worldwide [26]. Rice crop is sensitive to salinity particularly during the seedling growth stage [27]. Therefore, screening of rice for salinity tolerance at seedling stage is important for the improvement of rice production under saline environment [28]. Effect of salt stress on different seedling based growth parameters was reported in different crops [29-31].
In the present study, GP was decreased under salt stress in the genotypes NIAB-IRRI-9, KSK-133 and Basmati-385. The results of this study were in agreement with previous studies [7,12,13] that germination percentage was seriously affected by salt stress especially in salt sensitive genotypes. However, it was reported that GP is not a single good criteria for assessing salinity tolerance because germination depends upon several other factors like storing conditions and temperature of seeds and time of harvesting, etc. [31].
Root is the organ which has direct contact with salt solution and thus it can be a good selection criteria for salinity tolerance [32]. In present study, RL was generally reduced under salinity stress in all genotypes except Basmati-198.Previously, the decrease in RL was reported under salt stress in rice [13] and wheat [32]. In the current study, significant differences were observed among the genotypes for SL. Various studies [12,13] have reported the salt stress induced decrease in shoot length in rice particularly in salt sensitive cultivars.
In the present investigation, significant variation was observed among the genotypes for root/shoot ratio under salt stress as compared to control. Root/shoot ratio was decreased under salt stress in all genotypes except Basmati-385. Thus, it can be suggested that root/shoot length ratio has negative relation with salt tolerance in rice.
High salinity level causes reduced plant growth [33,34] which ultimately leads to reduced SFW. A similar response was observed in the current investigation that SFW was reduced under salt stress in all genotypes except Basmati-198 and statistically significant differences were observed for SFW under salt stress as compared to control. The current findings agreed with previous studies [35,36] that salinity reduces the fresh weight of rice seedlings. Since Basmati-198 was observed to be a salt tolerant variety and it also maintains high SFW under salinity. Thus it can be logically inferred that SFW is a good indicator of salt tolerance.
NaCl stress leads to the reduction in SDW [32]. In the present study, SDW was reduced under salt stress in all genotypes except KSK-133 and significant variation was observed among the genotypes. However, SDW was least affected in NIAB-IRRI-9 and Basmati-198.Similar response of rice genotypes was also observed for SDW under salt stress by other researchers [13] and it was mentioned that salt stress induced decrease in SDW in several other crops [29,37,38].
High salinity injects Na+ or Cl– in plants which mark them with necrosis and leaf tip burn [6]. There was a variable response to Na+ uptake by different genotypes and tolerant genotypes showed less Na+ uptake than sensitive ones [17]. In the present study, significant differences were observed among the genotypes for Na+ uptake under salt stress (Table: 2, Fig. 2). Highest Na+ concentration was observed in the genotype Basmati-385 while lowest Na+ uptake was observed in the genotype KSK-133 under saline conditions. Thus, it can be inferred that high Na+ uptake is related to salt sensitivity. While K+ uptake was increased under salt stress in the genotypes NIAB-IRRI-9, KSK-133 and Basmati-385 and it was decreased in Super Basmati and Basmati-198 (Fig. 2). It was reported that high K+ uptake compensate the deleterious effects of Na+ ions in rice [16].
As limited information is available on the screening of Basmati rice germplasm against salinity stress at seedling stage, so this new information may be helpful to devise a breeding strategy against this menace. Based on the performance of recorded parameters under saline conditions the genotypes NIAB-IRRI-9, Basmati-198 and KSK-133 were proved to be relatively salt tolerant and these can further be evaluated in saline areas. Also the highest reduction in seedling fresh and dry weight, maximum Na+ uptake and increased ratio of root/shoot length indicates the salt sensitivity of Basmati-385. It is suggested that these seedling based morpho-physiological parameters were proved to be effective and this methodology can be used to screen large rice germplasm for salt tolerance at an early growth stage in a cost effective manner.
Acknowledgement
Authors thank Department of Plant Breeding and Genetics, University of Agriculture Faisalabad Pakistan, for supplying the seed of tested rice genotypes and space for experimentation. We also thank to Department of crop physiology for providing lab facility, including instruments and chemicals for physiological analysis.
References
- Masood SA, Jabeen S, Anum M, Naseem Z, Jamshaid A, et al. Genetic Association of transcriptional factors (OsAP2 gene family) to incorporate drought tolerance in rice. Life Science Journal, (2015); 12(3s): 71-76.
- Masood SA, Naseem Z, Anum M, Arshad S, Jamshaid A, et al. An overview of genetic improvement for drought tolerance in rice (Oryza sativa L.). Life Science Journal, (2015); 12(3s): 63-70.
- Abid M, Qayyum A, Dasti A, Wajid R. Effect of salinity and sar of irrigation water on yield, physiological growth parameters of maize (Zea mays L.) and properties of the Soil. J Res Sci, (2001); 12(1): 26-33.
- Wang Z-f, Wang J-f, Bao Y-m, Wu Y-y, Xuan S, et al. Inheritance of rice seed germination ability under salt stress. Rice Science, (2010); 17(2): 105-110.
- Munns R. Genes and salt tolerance: bringing them together. New Phytologist, (2005); 167(3): 645-663.
- Wahome P, Jesch H, Grittner I. Mechanisms of salt stress tolerance in two rose rootstocks: Rosa chinensis ‘Major’and R. rubiginosa. Scientia Horticulturae, (2001); 87(3): 207-216.
- Pushpam R, Rangasamy SS. In vitro response of rice genotypes to salt stress. Madras Agricultural Journal, (2000); 87(10/12): 694-697.
- Mohammad M, Shibli R, Ajlouni M, Nimri L. Tomato root and shoot responses to salt stress under different levels of phosphorus nutrition. Journal of Plant Nutrition, (1998); 21(8): 1667-1680.
- Meloni DA, Oliva MA, Ruiz HA, Martinez CA. Contribution of proline and inorganic solutes to osmotic adjustment in cotton under salt stress. Journal of Plant Nutrition, (2001); 24(3): 599-612.
- Khan M, Hamid A, Karim M. Effect of sodium chloride on germination and seedling characters of different types of rice (Oryza sativa L.). Journal of Agronomy and Crop Science, (1997); 179(3): 163-169.
- Lutts S, Kinet J, Bouharmont J. Changes in plant response to NaCl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. Journal of Experimental Botany, (1995); 46(12): 1843-1852.
- Akbari G, Sanavy S, Yousefzadeh S. Effect of auxin and salt stress (NaCl) on seed germination of wheat cultivars (Triticum aestivum L.). Pakistan Journal of Biological Sciences, (2007); 10(15): 2557-2561.
- Abbas A, Khan S, Hussain N, Hanjra MA, Akbar S. Characterizing soil salinity in irrigated agriculture using a remote sensing approach. Physics and Chemistry of the Earth, Parts A/B/C, (2013); 5543-52.
- Lee S, Lee EJ, Yang EJ, Lee JE, Park AR, et al. Proteomic identification of annexins, calcium-dependent membrane binding proteins that mediate osmotic stress and abscisic acid signal transduction in Arabidopsis. The Plant Cell, (2004); 16(6): 1378-1391.
- Vinocur B, Altman A. Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations. Current Opinion in Biotechnology, (2005); 16(2): 123-132.
- Deepa S, Subbaraman P, Narayanan SL. Evaluation of temperature sensitive genic male sterility based rice hybrids for adaptation in salt affected environments by AMMI analysis. Journal of Ecobiology, (2008); 23263-274.
- Zhang Z-H, Qiang L, Song H-X, Rong X-M, Ismail AM. Responses of contrasting rice (Oryza sativa L.) genotypes to salt stress as affected by nutrient concentrations. Agricultural Sciences in China, (2011); 10(2): 195-206.
- Steel RG, Torrie JH, Dickey DA Principles and procedures of statistics: A biological approach. Chapter: Book Name. 1997 of publication; 352-356. McGraw-Hill.
- Q Rao A, Bajwa KS, Puspito AN, Khan M, Abbas MA, et al. Variation in Expression of Phytochrome B Gene in Cotton (Gossypium hirsutum L.). Journal of Agricultural Science and Technology, (2013); 15(5): 1033-1042.
- Muzaffar A, Kiani S, Khan MAU, Rao AQ, Ali A, et al. Chloroplast localization of Cry1Ac and Cry2A protein-an alternative way of insect control in cotton. Biological research, (2015); 481-11.
- Ashraf MA, Shahid AA, Rao AQ, Bajwa KS, Husnain T. Functional characterization of a bidirectional plant promoter from cotton leaf curl Burewala virus using an Agrobacterium-mediated transient assay. Viruses, (2014); 6(1): 223-242.
- Abid M, Qayyum A, Dasti A, Wajid R. Effect of salinity and sar of irrigation water on yield, physiological growth parameters of maize (Zea mays L.) and properties of the Soil. Journal of Research (Science), (2001); 12(1): 26-33.
- Nasir IA, Tabassum B, Qamar Z, Javed MA, Tariq M, et al. Herbicide-tolerant sugarcane (Saccharum officinarum L.) plants: an unconventional method of weed removal. Turkish Journal of Biology, (2014); 38439-449.
- Awan M, Abass M, Muzaffar A, Ali A, Tabassum B, et al. Transformation of Insect and Herbicide Resistance Genes in Cotton (Gossypium hirsutum L.). Journal of Agricultural Science and Technology, (2015); 17(2): 287-298.
- Rao AQ, Bakhsh A, Nasir IA, Riazuddin S, Husnain T. Phytochrome B mRNA expression enhances biomass yield and physiology of cotton plants. African Journal of Biotechnology, (2013); 10(10): 1818-1826.
- Kronzucker HJ, Szczerba MW, Schulze LM, Britto DT. Non-reciprocal interactions between K+ and Na+ ions in barley (Hordeum vulgare L.). Journal of Experimental Botany, (2008); 59(10): 2793-2801.
- Flowers T, Yeo A. Variability in the resistance of sodium chloride salinity within rice (Oryza sativa L.) varieties. New Phytologist, (1981); 88(2): 363-373.
- Zeng L, Shannon M, Grieve C. Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters. Euphytica, (2002); 127(2): 235-245.
- Ashraf MY, Akhtar K, Sarwar G, Ashraf M. Evaluation of arid and semi-arid ecotypes of guar (Cyamopsis tetragonoloba L.) for salinity (NaCl) tolerance. Journal of Arid Environments, (2002); 52(4): 473-482.
- Ashraf MY, Akhtar K, Hussain F, Iqbal J. Screening of different accessions of three potential grass species from Cholistan desert for salt tolerance. Pakistan Journal of Botany, (2006); 38(5): 1589-1597.
- Ashraf MY, Akhtar K, Hussain F, Iqbal J. Screening of different accessions of three potential grass species from Cholistan desert for salt tolerance. Pakistan Journal of Botany, (2006); 38(5): 1589-1597.
- Khan M, Yasmin S, Ansari R, Shirazi M, Ashraf M. Screening for salt tolerance in wheat genotypes at an early seedling stage. Pakistan Journal of Botany, (2007); 39(7): 2501-2509.
- Cherian S, Reddy M, Pandya J. Studies on salt tolerance in Avicennia marina (Forstk.) Vierh.: effect of NaCl salinity on growth, ion accumulation and enzyme activity. Indian Journal of Plant Physiology, (1999); 4(4): 266-270.
- Takemura T, Hanagata N, Dubinsky Z, Karube I. Molecular characterization and response to salt stress of mRNAs encoding cytosolic Cu/Zn superoxide dismutase and catalase from Bruguiera gymnorrhiza. Trees, (2002); 16(2-3): 94-99.
- Ashraf MY, Akhtar K, Sarwar G, Ashraf M. Role of the rooting system in salt tolerance potential of different guar accessions. Agronomy for sustainable development, (2005); 25(2): 243-249.
- Hakim M, Juraimi AS, Begum M, Hanafi M, Ismail MR, et al. Effect of salt stress on germination and early seedling growth of rice (Oryza sativa L.). African Journal of Biotechnology, (2010); 9(13): 1911-1918.
- Pessarakli M, Huber J. Biomass production and protein synthesis by alfalfa under salt stress. Journal of Plant Nutrition, (1991); 14(3): 283-293.
- Al‐Rawahy S, Stroehlein J, Pessarakli M. Dry‐matter yield and nitrogen‐15, Na+, Cl‐, and K+ content of tomatoes under sodium chloride stress 1. Journal of plant nutrition, (1992); 15(3): 341-358.