

Full Length Research Article
In silico advancement of Aspergillus niger gluconic acid hyperproduction recommends plans for switching acid productivity
Aiman Habib1*, Sher Muhammad2*, Khalid Naveed3, Tayyaba Huma1*, Asad Ali Shah1, Kamran Rashid1,4, Khadim Hussain1,5*, Temoor Ahmed5,6,7, Muhammad Ismail8, Irfan Manzoor1, Muhamamd Faisal9, Mohammed Aladhadh10
Adv. life sci., vol. 12, no. 1, pp. 62-70, February 2025
*- Corresponding Author: Sher Muhammad (Email: sher.muhammad@superior.edu.pk)
Authors' Affiliations
2. Faculty of Agriculture and Veterinary Sciences, Superior University Lahore – 56000 – Pakistan
3. Department of Plant Pathology, University of Agriculture, Faisalabad, Sub-campus Depalpur, Okara – 56300 – Pakistan
4. Department of Entomology, University of Georgia, 1109 Experiment Street, Griffin – GA 30223 – USA
5. Plant Protection Department, College of Food and Agriculture Sciences, King Saud University Riyad – Saudi Arabia
5. Department of Life Sciences, Western Caspian University, Baku – Azerbaijan
6. Department of Plant Biotechnology, College of Life Science, Korea University, 145 Anam-ro, Seongbuk-gu, Seoul – 02841 – Republic of Korea
7. Advanced Research Centre, European University of Lefke, Lefke, Northern Cyprus, TR-10 Mersin –Turkey
8. Department of Biotechnology, Akhuwat-Faisalabad Institute of Research Science and Technology, Faisalabad – 38000 – Pakistan
9. Food Department, Government of Punjab, Lahore – Pakistan
10. Department of Food Science and Human Nutrition, College of Agriculture and Food, Qassim University, Buraydah – 51452 – Saudi Arabia
[Date Received: 28/04/2023; Date Revised: 09/09/2024; Date Published: 31/12/2024]
Editorial Expression of Concern:
Editorial Note:
28 May 2025: You are viewing the latest version of this article having minor corrections in use of English language. Readers may please see figures with higher DPI given at the end of Results section. Expression of concern is hereby revoked.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Non-toxic and non-volatile organic acid, gluconic acid (pentahydroxycaproic acid) is found in abundance in honey and plants and in wines. Gluconic acid and its salts are used widely in food, feed, pharmaceuticals, textiles, and many other industries.
Methods: In the current investigation, Aspergillus niger was utilized for Gluconic acid production in submerged fermentation. Furthermore, for Gluconic acid hyperproduction, the fungus Aspergillus niger was exposed to MNNG (100 µg/ml) for 15 minutes. Aspergillus niger strain (parent and mutant) was tested and identified for its capability of producing gluconic acid. We finished the molecular identification, in which we extracted the DNA from the A. niger and amplified its ITS region using specific primers. We further sequenced the amplified product and created a phylogenetic tree employing the MEGA6 software.
Results: The tree results showed 100% identity with our target sequence with Aspergillus niger, which proved that the isolated strain belongs to the Aspergillus genera. In addition, factors related to the production of gluconic acid were confirmed by cultivating the mutant strain A. niger (MG1) and the parent strain under several conditions. These included varying the pH (4-7), temperature (30-40°C), and the concentration of the carbon source (40-100 g/l). The outcomes displayed that the best pH was 5. The most favorable temperature was 30°C, and maximum production occurred at a carbon source concentration of 100 g/l, for both mutant and parent strains.
Conclusion: This study shows how microbial strains and substrates could be used to attain cost-effective hyperproduction of gluconic acid to achieve objectives of industrial significance.
Keywords: Gluconic Acid; Aspergillus niger; Hyper-production; Mutation; Molecular identification; Sequencing
Introduction![]()
Gluconic acid (GA, C6H12O7, 2,3,4,5,6-pentahydroxycaproic acid) is a non-toxic, non-volatile, bio-based minor organic acid that is in abundant supply [1]. It is commonly found in honey, various kinds of plants, and most fruit. It is also present in various unexpected sources, too, including wine and meat. Though, GA is a relatively simple structure, it can be prepared commercially only by the oxidation of glucose [1]. Due to its unique characteristics, Gluconic acid has wide applications in pharmaceutical, food and feedstuff, cleansing agent, textile, beverage, leather, graphic as well and construction activities [2–4]. The demand for Gluconic acid increased exponentially in the preceding years, especially in biological and pharmaceutical procedures. The international market extended 800,000 tons annually [5]. Four different methods are being adopted for Gluconic acid production at an industrial scale [6]. The initial technique of Gluconic acid production is a chemical method where glucose is oxidized in the presence of the hypochlorite solution [7]. The following gluconic acid production process involves fermentation, using a suitable culture containing glucose and other components [8]. Another famous procedure of Gluconic acid production is the electrolytic oxidation of glucose in bromide presence [9]. Preceding but not least, the photocatalytic process is used for Gluconic acid production [10].
Gluconic acid production by fermentation is a remarkable topic for investigators these days [3]. The fermentation procedure is considered a controlling and effective method for Gluconic acid production [11]. The variation in the fermentation or microbial process for producing gluconic acid reduces practical challenges in the biochemical process, increases the production rate and yield of gluconic acid, and enhances its quality [12]. For fermentation of Gluconic acid diverse species are under concern like Aspergillus, Pseudomonas flurorescens, Penicillium, Pseudomonas plecoglossicida, Arthrobacter globiformis, Gluconobacter, Zymomonas [13]. On the other hand, Aspergillus is considered a revolution in the fermentation of Gluconic acid. Aspergillus is a genus that comprises of 300 mold species [14]. Aspergillus is broadly spread in the atmosphere due to its spores being plentiful in the air. Aspergillus can bear exciting conditions if all other conditions are practicable [15]. Aspergillus need water and temperature accessibility for their development [16]. Aspergillus is considered appropriate for gluconic acid production due to its ability to produce enzymes. It yields all the enzymes consumed in the translation of glucose to gluconic acid [6]. These enzymes are lactonase, mutarotase, catalase, and glucose oxidase [6]. Usually, the crystalline glucose monohydrate comes in alpha arrangement and converts into beta arrangement during the gluconic acid fermentation [6]. One more circumstance is that it helps Aspergillus in the mutarotase enzyme that accelerates the production of Gluconic acid [17,18].
Multiple investigations are made on Aspergillus genera especially on Aspergillus niger due to its high gluconic acid productivity. A. niger is considered a breakthrough in fermentation for the production of organic composites, for example gluconic acid, proteins, and citric acid [19–21]. To attain the gluconic acid hyper-production, a strain of Aspergillus niger was mutated and identified by molecular methods, and sequenced and different parameters like carbon source, pH, temperature, and inoculum size were adjusted for the gluconic acid production and kinetic parameters determined for growth and gluconic acid formation (Yp/x, µ, qp) by A. niger and its mutant strains in diverse growth environments.
Methods![]()
Microorganism
The Bioprocess Engineering Lab at the Government College University Faisalabad (GCUF), Pakistan, provided the fungus for our study which was identified morphologically by our research group and was sequenced and assigned an accession number SUB14433379 Aspergillus PP767952. The Aspergillus niger was sustained on the PDA plates by incubating for 48 hrs at 30 °C. Petri plates were preserved at 4 °C after sporulation and subcultured each month.
Mutagenesis
Repeated subculturing on potato dextrose agar (PDA) slants was used to produce the monospores of A. niger at 30 °C for 4-5 days. The obtained colonies were used to produce the monospores. Subsequently, sterilized 0.001 % (v/v) Tween-80 was mixed and the slant was shaken for five minutes to yield the monospores. The spores were splashed with phosphate buffer 0.5M (pH 7.0). The spores were consecutively diluted by using the phosphate buffer and spread on the PDA plates. Then the standardized spore suspension (1×10-7 spores/ ml) was used to monitor the gluconic acid production. The highest-yielding spore suspension was nominated and transformed with n-methyl n-nitro-n-nitrosoguanidine (MNNG). 1ml of MNNG (1.0 mg/ml) was mixed in the 5 ml gluconic acid-producing monospore suspension in the 50 ml falcon. The mix was protected at 30°C for different time intervals (20, 30, 40, and 50 minutes) in a rotating shaker agitated at 150 rpm. Following incubation, it was rinsed three times with 0.2M phosphate buffer to eliminate MNNG. Following this, the culture was consecutively diluted by the similar buffer and sub-cultured on PDA agar dishes. These dishes were then protected at 30°C intended for 4-5 days. Numerous colonies emerged; from these, the well-developed colony MG1 was chosen based on the zone of clearance observed on the plate. Afterward, consecutively diluted culture with the similar buffer was sub-cultured on the PDA agar dishes. Subsequent dilutions of the original culture, using the same buffer, were plated onto PDA agar dishes. These dishes were protected at 30 °C for 4 to 5 days. Several colonies appeared, among them, one well-established MG1 colony, which produced a zone of clearance that was evident on the plate [22].
Identification
Molecular approaches proposed vast benefits over old methods for fungal identification. Although conventional methods are still in use, they have low diagnostic sensitivity, high turnaround time, and less species-specific sensitivity. Molecular fungal identification includes polymerase chain reaction (PCR) and sequence-based assay.
DNA Extraction
A number of methods are available for DNA isolation for fungal strains. Typically, all methods involve lysing cells in the starting material and then eliminating proteins and other contaminants to end up with purified DNA. The A. niger (parent and mutant) strain’s DNA was extracted through the fungal DNA extraction technique [23].
The fungal cell mass was cultivated primarily. For this purpose, the A. niger (parent and mutant) was grown in Vogel’s media. The mycelia were separated from the 50 ml culture by using the filter paper and air-dried it followed by the addition of 10 ml extraction buffers (200mM Tris Hcl, 25 mm EDTA, 250 mm NaCl, plus 0.5% SDS) according to the (1-2g) mycelia weight and 5 ml sodium acetate buffer (3M sodium acetate) of 5.2 pH. The mixture was protected at -20˚c for ten minutes. The mix was centrifuged at 10,000 rpm for five minutes or until the compact pellet was made. The resultant pellet was discarded and the isopropanol (v/v) ice cold was added in the supernatant at room temperature for five minutes. Afterwards, the mixture was centrifuged again at 10,000 rpm for 5 min, and kept the supernatant at this time. The ice-cold 70% ethanol was added volume by volume and centrifuged it. The resulting pellet was dehydrated in the incubator at 37˚C and dissolved in 500µl d3H2O. The extracted DNA was analyzed on 1% agarose gel.
Amplification of DNA
The pair of primers was used to amplify the ITS region (internal transcribed spacer region) that is situated among the conserved genes encoding 28S, 5.8S, and 18S ribosomal subunits. The ITS region could be amplified from an inclusive variety of fungi using the ITS1 and ITS4 primers. The PCR reaction was performed in 25µl volume containing 2mM dNTPS 2.5µl, 10X Taq buffer 2.5µl, 25mM MgCl2 2µl, forward primer (ITS1 5’- TCCGTAGGTGAACCTGCGG -3') 3µl, reverse primer (ITS4 5’ TCCTCCGCTTATTGATATGC 3’) 3µl, Taq polymerase (5U/µl) 0.3µl, and double distilled deionized water 9.2µl. The PCR consisted of first denaturation at 95˚c for five min followed by 94˚c for 30sec, 55˚c for 30sec, 72˚c for 45 sec, and 72˚c for five minutes. The PCR product was run on the 1% agarose gel containing 0.5X TAE buffer visualized by Ethidium bromide under UV light. The DNA fragment's size was evaluated in comparison to the Gene Ruler TM 1Kb ladder. The PCR product was refined by the PCR clean-up Mini Kit Favor Prep (FAPCK001-1).
Sequencing of purified PCR product and in-silico analysis of purified PCR products
Using gene-specific primers, the Macrogen Company in Korea sequenced the PCR product in more depth. We took the sequences that we obtained and aligned them against the GenBank database using NCBI's BLASTN. The fungal identification was finished based on extreme uniqueness ≥98% and query coverage ≥90%.
ITS sequence was aligned by Multiple sequence alignment and through this alignment, the unrooted phylogenetic tree was created by employing MEGA6 software by the help of Aspergillus niger strain/isolates used in phylogenetic tree construction. The evolutionary history was concluded by Neighbor-Joining method. The association's confidence level and stability were evaluated using bootstrap methods. This involved resampling 1,000 times from the original data set. The results were assessed to give a bootstrap value associated with each branch point in the tree. The tree diagram represents the bootstrap results.
Fermentation of Gluconic acid
The submerged fermentation of gluconic acid was completed in 250 ml volume cotton wool plugged Erlenmeyer flask with 50 ml media. However Vogel’s media was used of the following composition (g/ml) Tri sodium citrate (0.5), Dipotassium phosphate (0.5), Ammonium sulphate (4), Ammonium nitrate (0.2), Magnesium sulphate heptahydrate (0.2), Yeast extract (0.1), Glucose (1) and Agar (3). A 50 µl sample of the antibiotic was added to sterilized medium and mixed. A loopful of the wild-type and mutant strains of A. niger was then inoculated into the medium. The inoculated cultures were protected from light and protected for 48 hrs at 30°C in an orbital shaker on 150 rpm. To perform various assays, aliquots were collected at 4-hour intervals.
Gluconic acid assay
Detection of gluconic acid was carried out with ferric chloride. The supernatant was placed in test tubes, and a drop of ferric chloride was added in separate tubes. The color changed to deepened yellow at an absorbance of 560 nm [24].
Protein Estimation
Measurement of total protein was done by using standard laboratory protocol. This was then used to estimate the protein content in an unknown sample by taking 1 ml of Bradford reagent and added with 100 µl of diluted enzyme, respectively at OD595 nm.
Total Reducing Sugars
Total reducing sugar contents were estimated by using dinitro salicylic acid (DNS) method.
Growth optimization of parent and MG1 of A. niger
Quadruplicate flasks of wild and MG1 were subjected to time course study by collecting an aliquot from each flask at 4-hour intervals during the experimental period. For the gluconic acid assay, tested it at 10,000 rpm for 5 minutes using a centrifuge. The gluconic acid test was performed using the cell mass-free supernatant, whereas spectrophotometrically diluted cell mass was used for taking a reading of them.
Effect of different temperatures and pH on Gluconic Acid production
We investigated the temperature effect on the production of gluconic acid. To do this, we optimized our experimental conditions and then incubated two different strains (MG1 and a wild-type strain) in shaking flasks (250 ml) at temperatures of 30-40 ± 1°C. We also studied the influence of the initial pH on gluconic acid production. We adjusted the pH in the carbon and nitrogen source medium to a range of 4-7 ± 0.1. We used the two strains (MG1 and wild-type) in these experiments as well.
Determination of kinetic parameters
Growth kinetics was measured as described in [25]. Kinetics of substrate utilization (Gluconic acid production) and determination wild type by cell mass & Mutant (MG1), in triplicate. Specific growth rate (m) was calculated based on the flasks picked in triplicate after each 4 h. We plotted in (X) against time to find the value of μ, where X is the cell mass (g) in the medium during the exponential stage. We calculated separate curves for cell mass (g/l), substrate (g/l s), and the volumetric production rate of gluconic acid (g/l) in time (h) of the fermentation to find out the maximum volumetric rates, otherwise known as extremes (Qs, Qx, and Qp). A number of growth kinetic coefficients were noted by employing the formulae given as:
The cell yield coefficient (YX/S) indicates how many grams of cell mass are produced from each gram of substrate consumed. The specific substrate consumption rate (qs) represents the biomass production rate in grams of substrate required to generate one gram of cell mass in one hour.
Product yield (YP/S), IU per g of substrate used = PM/SM
Precise product produce (YP/X), IU/g dry cell mass = PM/XM
Specific rate of product development (qP), IU/g cell mass per h = m . YP/X
In this context, XM, SM, and PM refer to the largest quantities of substrate, cells, and enzymes, respectively.
Results![]()
Mutagenesis
MNNG works in preference at the DNA replication fork, and causes a mass of mutations which will occur very early. MNNG alkylates the pyrimidine and purines to induce mutation. This mutation causes transition mutation which is more common for the G: C to A: T transition than that of the A: T to G: C, this type of changes in gluconic acid producing bacteria would make at transcription & translation levels that leads a incrementation in Gluconic Acid production and stabilizes enzyme active site towards substrate tie up conversion into Gluconic Acid. Later on, the culture was 10 times serially diluted then sub-cultured onto PDA agar plates using similar buffer. Also, protected these plates at 30 °C for 4-5 days. The multiple colonies were obtained and selected from the plates, Single well-developed colony MG1 was selected from some of the zones clearance appeared on plate. Around 3–4% of cells were surviving. Multiple colonies grew on the plate and three to four large single colonies from both plates were picked as mutants onto medium plates that was incubated at 30°C on a static incubator there was growth seen on the plate after 48h.
Identification of Fungus
This study utilized both the parent and mutant soil isolate of A. niger for molecular characterization. To achieve this, we cultured the A. niger isolates on Vogel's medium. The resultant mycelia were then used for the DNA extraction. The 1% agarose gel showed that the DNA from both parent and mutant of A. niger prepared equally, using ladder of a one kilobase (Kb) (Fig 1A and 1B). Two pair of primers (ITS & ITS4, and ITS1 &ITS4) were used for amplification of the genomic DNA A. niger isolate for this study from the respective region called the ITS domain. The bands of approximately 600bp were confirmed by results. Both primers effectively amplified the ITS domain (Figure 1C & 1D). The amplified DNA was subsequently purified using the Favor Prep PCR Clean-up mini kit to remove residual components from the original polymerase chain reaction (PCR) mixture (Fig 1E and F).
Sequencing of the PCR product of ITS region of Aspergillus niger
We shipped the refined product of 600 bp out for sequencing to Macrogen, Korea. We used the sequence-specific primers to obtain the necessary data. Once we received the results, we compared them to nucleotide sequences in the database (GenBank) using the BLAST (Basic Local Alignment Search Tool). We found that our query sequence had 100% identity with Eurotiales sp. We then took those data, along with the information from the homologous sequences, and produced the unrooted Neighbor Joining tree employing the MEGA6 software (see Figure 2). The sequences descending from the mutual ancestor were gathered together similar the AH1ITS1.ab 11292 group together with Eurotiales sp. NC1578, Eurotiales sp.1428, Aspergillus awamori strain SRRC332, and Aspergillus sp. XN-3-2-3 and have 100% identity with a very negligible nucleotide substitution rate, indicating that the ITS sequences have evolved but the substitution rate was very negligible at 0.1%. It depicted that our isolate belongs to the Aspergillus genera.
Growth kinetics
To obtain a substantial concentration of gluconic acid requires the choice of the right media. This study utilized Vogel's medium and achieved a great yield of gluconic acid, with 1.05 g/l from the parent strain and 1.95 g/l from the mutant strain (MG1). These results underscore the media's crucial role and choice in the production of gluconic acid, which can serves as an important platform chemical in various industrial sectors, such as food processing and pharmaceuticals. Importantly, the results also hint at the potential of the selected mutant strain to outpace its parent strain (and likely, laps in genetic path) in the biotechnological production of said acid.
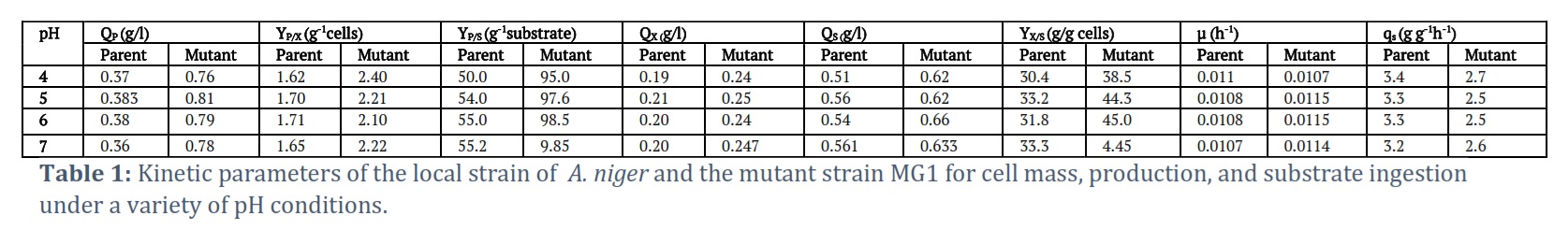
Effect of pH
The present research explored how different pH levels, from 4 to 7, affected the production of gluconic acid. The optimal production of gluconic acid occurred at pH 5, with the parent strain generating 0.383 g/l and the mutant strain producing 0.81 g/l after 48 hours of incubation. Higher or lower pH levels resulted in less production. Consequently, the maximum production occurred in the well-type plate at pH 5. The study underscores the significance of the investigated conditions: temperature, substrate, and pH. It highlights the necessity for maintaining conditions that are optimally favorable and also reveals the effects of unfavorable conditions on the growth of both the parent and mutant strains, as shown in Table 1. Parent strain benchmarked a maximum production of gluconic acid of 0.383 g/l, while the mutant strain produced 0.813 g/l, at 48 h of incubation at pH 5. It seemed that production rate and cell mass were reduced above and below this pH. The specific value of growth and product yield coefficients Yx/s, Yp/s for mutant strain were also greater as compared to parent strain at different pH. (Table 1) which indicates that gluconic acid production rate with respect to substrate utilization was greater for mutant (MG1). The values of definite rate constants (Qp, Qx and Qs in g/l/h) were also extra significant in case of MG1 mutant strain of Aspergillus niger as depicted in Table 1. In the current research, interestingly the precise growth rate (µ = 0.01h-1) remained almost same at different pH for both parent and mutant strain.
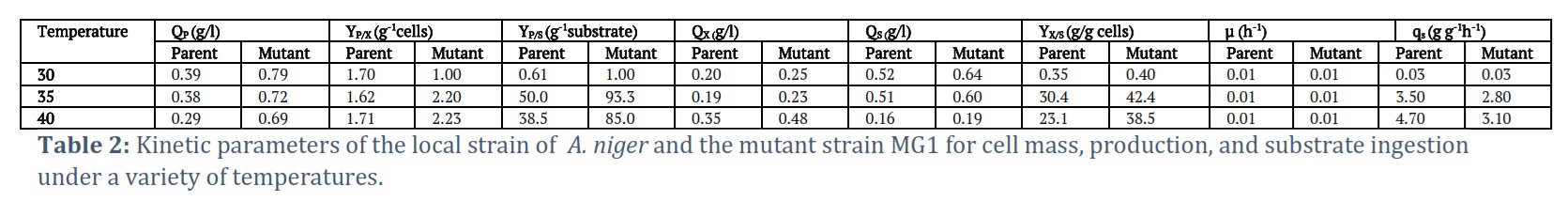
Effect of temperature
In order to improve the temperature settings, both the parent and the mutant strains (MG1) of A. niger were cultivated at several temperatures between 30 and 40 °C. The maximum gluconic acid yields for both strains occurred at the lowest temperature in the range, 30 °C; the parent strain produced gluconic acid at a yield of 0.39 g/l while the mutant strain yielded 0.79 g/l. On the other hand, substantial decreases in yield occurred at temperatures above 30 °C, and the highest temperature (40 °C) yielded 0.29 g/l and 0.68 g/l for parent and mutant strains, respectively (Table 2). These results suggest that gluconic acid productivity is thermodynamically favorable at 30 °C and that the metabolic pathways are more efficient and/or better regulated at that temperature when both strains work.
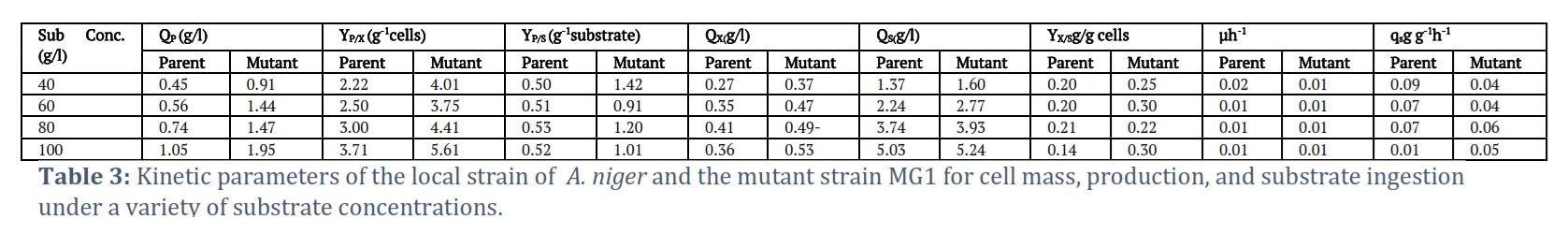
Effect of substrate concentration
To achieve optimum enzyme activity and obtain the maximum amount of product, the substrate concentration must be appropriate. The study examined several different concentrations of glucose, from 4% to 10%, and discovered that the highest production levels occurred at 10%. The parent strain yielded 1.05 g/L and the mutant strain (MG1) 1.95 g/L. The details on how much substrate was consumed, how much cell mass was obtained, and the production amounts for both the parent and mutant strains appear in Table 3.
Figures & Tables


Multiple investigations are made on Aspergillus genera especially on Aspergillus niger due to its high gluconic acid productivity. A. niger is acknowledged as a breakthrough in fermentation for the production of organic composites, such as gluconic acid, proteins and citric acid, [19–21]. To attain the gluconic acid hyper-production, a strain of Aspergillus niger was mutated and identified by molecular methods, and sequenced and different parameters like carbon source, pH, temperature, and inoculum size were improved for the gluconic acid production and kinetic parameters determined for growth and gluconic acid development (µ, qp, Yp/x) through A. niger and its mutant products under different growth conditions. In our study, we found that the compound MNNG preferentially acts on the DNA replication fork and induces a large number of mutations at the origin of replication. MNNG stimulates the mutation process by alkylating the bases of DNA, which primarily results in G:C to A:T transitions, while the A:T to G:C transition is far less common [26]. This mechanism of action clearly has impacts at the genomic level; however, because our primary interest is in the effects of these mutations at the functional level, we focused our analysis on the enzymes that are part of the process leading to gluconic acid formation [27]. After identifying several colonies, we sub-cultured a single well-developed colony (MG1) on PDA plates, which appeared to have a zone of clearance [22].
Molecular identification of the Aspergillus niger soil isolate (both parent and mutant) was performed in the study. To amplify the internal transcribed spacer (ITS) region, two pairs of primers were used (ITS1 & ITS4, and ITS & ITS4). Both primer pairs produced a positive result. The purification product (600 bp) was sequenced. The sequencing result showed 100% identity with Eurotiales sp. The homologous sequences found in the database were then used to draw an unrooted Neighbor Joining tree. MEGA6 software was used for this part of the work. The lines of descent from the common ancestor were gathered together, one might say "bunched together," like AH1ITS1.ab 11292, which is found in Eurotiales sp. NC1578, Eurotiales sp. 1428, Aspergillus awamori strain SRRC 332, and Aspergillus sp. XN-3-2-3. These lineages have 100% identity, with an exceedingly low substitution rate among the nucleotides. It was depicted in our research that our isolate belongs to the Aspergillus genera.
The phylogenetic relationships of Greek isolates of Aspergillus species have been ascertained using sequence data from the ITS and the IGS region [28]. These components of the AFLP and ITS rDNA have also been used for species delineation of the the A. niger collection [29]. The current study builds on this work by determining the phylogenetic relationships of some of the Aspergillus species found in the Greco-Roman world; it also examines the good-yielding organic acid (gluconic acid) production potential of two isolates (one parent and one mutant) in the A. niger phylogenetic group. For the current research, the same media used for the itaconic acid yield studies, as well as for acid phosphatase studies, was used to grow both A. niger isolates to determine the activity levels of glucose oxidase, an important therapeutic enzyme [30]. The A. niger parent isolate produced 1.05 g/l of gluconic acid, while the amount produced by the mutant was approximately twice that of the parent at 1.95 g/l. In adding, [31] used Vogel’s media for the production of acid phosphatase from strain N402A of A. niger. Likewise, [32] researched the glucose oxidase action by growing A. niger on vogel’s media broadcasting an action of 2.32 U mg-1.
To ferment successfully, it is essential to maintain pH because each enzyme operates best at its optimal pH level. The enzyme responsible for the production of gluconic acid, glucose oxidase, achieves its maximum production at pH 5 [33]. Both below this pH and above pH 7, production levels are significantly lower. For this study, the effect of five different pH levels from 4 to 7 on gluconic acid production was tested. The results found that the parent and mutant strains produced gluconic acid in the following quantities: parent strain, 0.383 g/L; mutant strain, 0.81 g/L; both at 48 hours post-inoculation and at pH 5. Reductions in both cell mass and production levels were observed for both strains on either side of pH 5. Despite the study’s testing of five different pH levels, it is important to note that previous studies by [33–36] also indicated high accumulation of gluconic acid at pH levels not exceeding pH 5. To achieve the ideal activity level for the enzyme and to yield the maximum quantity of product, the proper concentration of substrate is necessary. Thus, in our research, we examined various concentrations of glucose, ranging from 4% to 10%. We found that the greatest production level was reached at the 10% substrate concentration, yielding 1.95 g per L for the mutant (MG1) and 1.05 g per L for the parent. This work (not ours but that of others) corresponds to findings reported in [35,37]. In optimizing the temperature conditions for our research, we cultivated both the parent and mutant MG1 strains of A. niger at different temperatures, ranging from 30 to 40 °C. The maximum gluconic acid production for both strains occurred at 30 °C, with the parent strain producing 0.39 g/l and the mutant strain yielding 0.79 g/l. This temperature also matched the optimum found by [34], who identified that the highest gluconic acid production (58.18 g/l) occurred at 30 °C. In a similar vein, [35] recorded maximum gluconic acid production (58.46 g/l) at this same temperature. To achieve the best enzyme activity and synthesis of the maximum product, the substrate must be present at an ideal concentration. We varied glucose concentration at levels ranging from 4% to 10% in our investigation. The outcome was that we achieved the highest production at 10%, yielding 1.05 g/l for the parent strain and 1.95 g/l for the mutant MG1. This work corresponds with that of [35,37].
Gluconic acid was studied extensively from last century for quality enhancement and production. The purpose of the study is to find out the economical approaches to reduce the acid expenses. However, the present research assessed the local isolate of A. niger based on total production of gluconic acid. For this, the strain was subjected to mutagenesis with MNNG (100 µg/ml) to derive a hyper-producing variant. Identification of the A. niger strain, which was found to produce the highest amount of gluconic acid, was done using the ITS region of the rRNA gene. For this, a purified PCR product was sequenced by the commercial service provider Macrogen, located in South Korea. Subsequent to this, the search for homologous sequences was performed in the NCBI and Uniprot databases, and the results were aligned and used to construct a phylogenetic tree. Additional production conditions were assessed by culturing both the parent and the mutant (MG1) strain of A. niger at pH values ranging from 4 to 7, temperatures from 30 to 40 °C, and varying concentrations of a carbon source (40 g/l–100 g/l). Maximum yields of gluconic acid were obtained at pH 5 for both the parent and the mutant strain, with 30 °C being the best temperature for both strains. Additionally, the mutant and parent strains of A. niger produced the maximum amount of gluconic acid when cultured with a 100 g/l concentration of the carbon source.
Acknowledgement
The Researchers would like to thank the Deanship of Graduate Studies and Scientific Research at Qassim University for financial support.
Author Contributions
Aiman Habib, Sher Muhammad: These authors have contributed equally to this work therefore share the first authorship. The rest of the authors contributed in the research design, experimentation, analyses and manuscript writing in their individual capacity.
The author declare that there is no conflict of interest regarding the publication of this paper.
![]() References
References
- Cañete-Rodríguez AM, Santos-Dueñas IM, Jiménez-Hornero JE, Ehrenreich A, Liebl W, García-García I. Gluconic acid: Properties, production methods and applications—An excellent opportunity for agro-industrial by-products and waste bio-valorization. Process Biochemistry, (2016); 51(12): 1891-903.
- Ma Y, Li B, Zhang X, Wang C, Chen W. Production of gluconic acid and its derivatives by microbial fermentation: Process improvement based on integrated routes. Frontiers in Bioengineering and Biotechnology, (2022); 10: 864787.
- Pal P, Kumar R, Banerjee S. Purification and concentration of gluconic acid from an integrated fermentation and membrane process using response surface optimized conditions. Frontiers of Chemical Science and Engineering, (2019);13(1): 152-63.
- Anastassiadis S, Morgunov I. Gluconic acid production. Recent Patents on Biotechnology, (2007); 1(2): 167-80.
- Abtew E, Ezra AF, Basu A, Domb AJ. Biodegradable poly (acetonide gluconic acid) for controlled drug delivery. Biomacromolecules, (2019); 20(8): 2934-41.
- Yadav P, Chauhan AK, Singh RB, Khan S, Halabi G. Organic acids: microbial sources, production, and applications. In Functional Foods and Nutraceuticals in Metabolic and Non-Communicable Diseases, (2022); p.325-337, Elsevier.
- Varničić M, Zasheva IN, Haak E, Sundmacher K, Vidaković-Koch T. Selectivity and Sustainability of Electroenzymatic Process for Glucose Conversion to Gluconic Acid. Catalysts, (2020); 10(3): 269.
- Singh V, Haque S, Niwas R, Srivastava A, Pasupuleti M, Tripathi CKM. Strategies for fermentation medium optimization: An in-depth review. Frontiers in Microbiology, (2017); 7: 2087.
- Moggia G, Kenis T, Daems N, Breugelmans T. Electrochemical oxidation of d‐glucose in alkaline medium: Impact of oxidation potential and chemical side reactions on the selectivity to d‐gluconic and d‐glucaric acid. ChemElectroChem, (2020); 7(1): 86-95.
- Yin J, Zhang Q, Yang C, Zhang B, Deng K. Highly selective oxidation of glucose to gluconic acid and glucaric acid in water catalyzed by an efficient synergistic photocatalytic system. Catalysis Science & Technology. Royal Society of Chemistry, (2020); 10(7): 2231-2241.
- Zhou X, Zhao J, Zhang X, Xu Y. An eco-friendly biorefinery strategy for xylooligosaccharides production from sugarcane bagasse using cellulosic derived gluconic acid as efficient catalyst. Bioresource Technology, (2019); 289: 121755.
- D’Este M, Alvarado-Morales M, Angelidaki I. Amino acids production focusing on fermentation technologies–A review. Biotechnology Advances, (2018); 36(1): 14-25.
- Abd Alsaheb RA, Abdullah JK, Lafi ASA. Systematic comparison of gluconic acid optimization produced by Aspergillus using raw bioresources cultivation medium. Journal of Engineering Science and Technology, (2022); 17(1): 0673-88.
- Razzaghi-Abyaneh M, Rai M. Introductory Chapter: The Genus Aspergillus-Pathogenicity, Mycotoxin Production and Industrial Applications. In: Razzaghi-Abyaneh M, Rai M, editors. The Genus Aspergillus – Pathogenicity, Mycotoxin Production and Industrial Applications (2022); IntechOpen.
- Son YE, Cho HJ, Chen W, Son SH, Lee MK, et al. The role of the VosA-repressed dnjA gene in development and metabolism in Aspergillus species. Current Genetics, (2020); 66(3): 621-633.
- Lahouar A, Marin S, Crespo-Sempere A, Saïd S, Sanchis V. Effects of temperature, water activity and incubation time on fungal growth and aflatoxin B1 production by toxinogenic Aspergillus flavus isolates on sorghum seeds. Revista Argentina de Microbiología, (2016); 48(1): 78-85.
- Perrone G, Gallo A. Aspergillus Species and Their Associated Mycotoxins. In: Moretti A, Susca A, editors. Mycotoxigenic Fungi. (2017); p.33-49. Springer, New York.
- Mafra ACO, Ulrich LG, Kornecki JF, Fernandez-Lafuente R, Tardioli PW, Ribeiro MP de A. Combi-CLEAs of glucose oxidase and catalase for conversion of glucose to gluconic acid eliminating the hydrogen peroxide to maintain enzyme activity in a bubble column reactor. Catalysts, (2019); 9(8): 657.
- Pel HJ, de Winde JH, Archer DB, Dyer PS, Hofmann G, Schaap PJ, et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nature Biotechnology, (2007); 25(2): 221-231.
- Li C, Zhou J, Du G, Chen J, Takahashi S, Liu S. Developing Aspergillus niger as a cell factory for food enzyme production. Biotechnology Advances, (2020); 44: 107630.
- Meyer V, Wu B, Ram AFJ. Aspergillus as a multi-purpose cell factory: current status and perspectives. Biotechnology Letters, (2011); 33(3): 469-76.
- Raksha S, Srinivasan S, Prasant G, Prabu R. Over-expression of gluconic acid in Aspergillus oryzae RP-21 mutants generated by a random mutagenesis approach. 3 Biotech, (2012); 2(3): 219-23.
- Moťková P, Vytřasová J. Comparison of methods for isolating fungal DNA. Czech Journal of Food Sciences, (2012); 29: S76-S85.
- Talalay P, Fishman WH, Huggins C. Chromogenic substrates: II. Phenolphthalein glucuronic acid as substrate for the assay of glucuronidase activity. Journal of Biological Chemistry, (1946); 166(2): 757-772.
- Pirt JS. Principles of Microbe and Cell Cultivation. 4-44. Blackwell Scientific Publications, Cornwall.
- Prabu R, Rosfarizan M, Shah UM, Ariff AB. Improvement of Aspergillus flavus Link S44-1 using random mutational method for kojic acid production. Minerva Biotecnologica, (2011); 23(4): 83.
- Gunka K, Newman JA, Commichau FM, Herzberg C, Rodrigues C, Hewitt L, et al. Functional Dissection of a Trigger Enzyme: Mutations of the Bacillus subtilis Glutamate Dehydrogenase RocG That Affect Differentially Its Catalytic Activity and Regulatory Properties. Journal of Molecular Biology, (2010); 400(4): 815-27.
- Krimitzas A, Pyrri I, Kouvelis VN, Kapsanaki-Gotsi E, Typas MA. A Phylogenetic Analysis of Greek Isolates of Aspergillus Species Based on Morphology and Nuclear and Mitochondrial Gene Sequences. BioMed Research International, (2013); 2013: 1-18.
- Giraud F, Dupont J, Haon M, Bouzid O, Alibeu O, Navarro D, et al. Phylogenetic analysis of the Aspergillus niger aggregate in relation to feruloyl esterase activity. Research in Microbiology, (2007); 158(5): 413-419.
- Blumhoff ML, Steiger MG, Mattanovich D, Sauer M. Targeting enzymes to the right compartment: Metabolic engineering for itaconic acid production by Aspergillus niger. Metabolic Engineering, (2013); 19: 26-32.
- Hidayat BJ, Eriksen NT, Wiebe MG. Acid phosphatase production by Aspergillus niger N402A in continuous flow culture. FEMS Microbiology Letters, (2006); 254(2): 324-31.
- Zia MA, Riaz A, Rasul S, Abbas RZ. Evaluation of antimicrobial activity of glucose oxidase from Aspergillus niger EBL-A and Penicillium notatum. Brazilian Archives of Biology and Technology, (2013); 56(6): 956-61.
- Anastassiadis S, Aivasidis A, Wandrey C. Continuous gluconic acid production by isolated yeast-like mould strains of Aureobasidium pullulans. Applied Microbiology and Biotechnology, (2003); 61(2): 110-7.
- Znad H, Markoš J, Baleš V. Production of gluconic acid from glucose by Aspergillus niger: growth and non-growth conditions. Process Biochemistry, (2004); 39(11): 1341-1345.
- Shindia AA, El-Sherbeny GA, El-Esawy AE, Sheriff YMMM. Production of Gluconic Acid by Some Local Fungi. Mycobiology, (2006); 34(1): 22-29.
- Ahmed AS, Farag SS, Hassan IA, Botros HW. Production of gluconic acid by using some irradiated microorganisms. Journal of Radiation Research and Applied Sciences, (2015); 8(3): 374-80.
- Sankpal NV, Kulkarni BD. Optimization of fermentation conditions for gluconic acid production using Aspergillus niger immobilized on cellulose microfibrils. Process Biochemistry, (2002); 37(12): 1343-1350.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()

