

Full Length Research Article
Batch fermentation and GC-MS analysis of biocontrol agent, Bacillus amyloliquefaciens strain KSAS6 and its impact on soilborne fungus, Sclerotium bataticola
Abdulaziz A. Al-Askar1, Fatimah O. Al-Otibi1, Mohamed T. Yassin1, Gaber A. Abo-Zaid2, Ahmed Abdelkhalek3
Adv. life sci., vol. 12, no. 1, pp. 260-270, February 2025
*– Corresponding Author: Abdulaziz A. Al-Askar (Email: aalaskara@ksu.edu.sa)
Authors' Affiliations
2. Bioprocess Development Department, Genetic Engineering and Biotechnology Research Institute, City of Scientific Research and Technological Applications, Alexandria – 21934 – Egypt
3. Plant Protection and Biomolecular Diagnosis Department, Arid Land Cultivation Research Institute, City of Scientific Research and Technological Applications, Alexandria – 21934 – Egypt
[Date Received: 08/09/2024; Date Revised: 07/11/2024; Date Published: 31/12/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Sclerotium bataticola, a soil-born fungus, is responsible for charcoal rot in a variety of plants. It is also responsible for causing substantial damage to a wide range of horticultural crops around the world.
Methods: Fifteen different Bacillus isolates were isolated and evaluated for their ability to inhibit S. batatacola's growth. The promising bacterial isolate was molecularly identified using NCBI-Blast and phylogenetic tree analysis of the 16S rRNA gene. Batch fermentation was performed in a stirred tank bioreactor to maximize culture biomass and secondary metabolite synthesis. Gas chromatography-mass spectrometry was used to discover secondary metabolite compounds.
Results: The KSAS6 isolate was the most effective for inhibiting the fungal growth of mycelial cells, with a 48.2% inhibition percentage. The probable biocontrol agent, B. amyloliquefaciens strain KSAS6, was identified and recorded in GenBank under the accession number PQ271636. The culture biomass and secondary metabolites were maximized by the batch fermentation technique, reaching the highest achievable level of 2.1 g L-1 at 11 hours. This was accomplished while maintaining a steady specific growth rate (µ) of 0.13 h-1. Based on the observations, the biomass yield coefficient was found to be 0.37 g cells/g glucose. Among the 21 secondary metabolite compounds identified in GC-MS analysis, diisooctyl phthalate was the highest compound (43.31%).
Conclusion: The strain of rhizobacterium B. amyloliquefaciens known as KSAS6 can inhibit the growth of S. bataticola, which makes it a promising candidate for the biocontrol of fungal infections in plants.
Keywords: Sclerotium bataticola; Bacillus amyloliquefaciens; Batch fermentation; Secondary metabolites; antifungal
Introduction![]()
Milk Fungi-caused diseases are among the most significant contributors to crop failure [1]. Over 19,000 different kinds of fungi have been identified as being responsible for infecting crop plants all over the world. Pathogenic fungi are responsible for approximately thirty percent of all agricultural diseases [2]. There is a wide variety of phytopathogens that can interact with plants, and a significant number of these pathogens are found in the soil. These disease complexes that are transmitted through soil are particularly challenging to control. Once established, these disease complexes can drastically limit the diversity of microorganisms, which in turn impacts the rhizosphere and endosphere of plants, thereby increasing the phytosanitary risks for crops [3]. Sclerotium bataticola is one of the soil-borne fungi. This fungus is transmitted through the soil and causes charcoal rot in a variety of plants, including potatoes, sweet potatoes, corn, sunflowers, and soybeans [4].
Over the past few decades, plant growth-promoting rhizobacteria (PGPR) have emerged as significant and promising tools for sustainable agriculture practices to manage plant diseases and increase crop yield [5]. This is because the productivity of crops is heavily dependent on the microbial communities that are present in the rhizosphere soil [6]. PGPR are microorganisms that are beneficial to soil-dwelling plants. These microorganisms can stimulate plant growth and make it easier for plants to absorb and use mineral nutrients [7,8]. PGPR has the potential to both boost crop productivity and decrease the incidence of disease. As a result, they are regarded as the most promising agents for the development of cash crops [9]. Bacillus species are rod-shaped, motile, Gram-positive bacteria commonly found in diverse environments, particularly in soil. They are a prominent group of rhizobacteria known for their ability to form spores, enabling them to survive in the soil for extended periods, even in harsh environmental conditions. This characteristic facilitates their germination in response to various environmental stimuli, prolongs the storage of the biocontrol agent, and simplifies the formulation process [10]. Consequently, beneficial strains of Bacillus that are considered major candidates for PGPR are well-regarded for their application in biological control and enhancing plant tolerance to environmental challenges [11]. Bacillus species have been recognized as biological agents capable of managing plant diseases using various approaches. These techniques involve employing hydrolytic enzymes to restrict pathogen growth as well as the secretion of antifungal compounds or lipopeptides [12]. Approximately fifty percent of the bacterial biocontrol agents that are now accessible for commercial use are Bacillus-based products [13]. Numerous Bacillus species have been utilized as applications for microbial biopesticides [14].
The development of the final product is influenced by various criteria that affect both the quality and quantity of goods. Amanullah et al. [15] and Bareither et al. [16] recommend researching these characteristics to determine the most appropriate microbial manufacturing line. The shake flask scale stage is a valuable screening stage that can be used to begin the examination of the variability of the raw materials, growth systems, inoculum size, pH, temperature ranges, and microbial responses to high and low agitation conditions. These data hold the potential to provide additional support for bioreactor evaluation [17–20]. In order to facilitate the development of microorganisms that will ultimately result in the production of a particular end product, the bioreactor offers a controlled and optimized environment that includes aeration rates, pH, improved mixing, and heat transfer. The bioreactor ought to be able to function in an aseptic manner for several days; it ought to be dependable when it is used for an extended time; and it ought to comply with the legislation governing containment [21,22]. In each fermentation process, mixing in the stirred tank bioreactor is of utmost significance for a significant number of different components of fermentation. The process will be sped up, the oxygen concentration will be increased, mass transfer will be improved, and the amount of time required for hydraulic retention will be decreased through efficient mixing [23]. Liu [24] cites several reasons why mixing is crucial: it prevents temperature stratification; maintains ideal pH levels; improves the interaction between the microbial culture and the substrate; prevents foaming; and provides a uniform distribution of the microorganisms and substrate. Furthermore, excellent mixing may prevent the buildup of harmful metabolites in the bioreactor sections, which are characterized by lower mixing [25]. In order to accomplish profitable production of the fermentation process, sustainability, and high cell density are critically crucial. To find the appropriate operating parameters, a comprehensive analysis is required [26,27].
In this study, the primary objective is to evaluate the inhibitory effect of fifteen Bacillus isolates as biocontrol agents against S. bataticola. Following this, 16S rRNA sequencing will be employed to identify the most promising isolate, KSAS6. To facilitate large-scale production of KSAS6 biomass and its secondary metabolites, batch fermentation will be conducted in a stirred-tank bioreactor. Finally, the secondary metabolites extracted from the culture filtrate will be analyzed using gas chromatography-mass spectrometry (GC-MS).
Methods![]()
Experimental isolation procedures
Isolation tests were conducted in the Kingdom of Saudi Arabia (KSA) using soil dilutions from the rhizospheres of tomato, potato, and eggplant [28]. A single gram of soil was agitated in a sterile saline solution containing 0.85% NaCl for fourteen minutes. 200 µL of the 10-5, 10-6, and 10-7 dilutions were added to the glycerol nutrient agar media and the peptone nutrient agar medium. Following a 48-hour incubation period at a temperature of 30 °C, many bacterial colonies became apparent. The detected colonies were isolated and purified using the process of isolating a single colony. The bacterial cultures were stored in a 50% glycerol broth at a temperature of -80 °C for future investigations.
Bacillus isolates antagonism test
An in vitro antagonist test was conducted to investigate the antagonistic effect of Bacillus isolates as biocontrol agents against S. bataticola, a plant pathogenic fungus. The antagonistic bacterial isolates were lined onto potato dextrose agar (PDA) plates after being cultivated for two days. This was completed 48 hours before any fungi were introduced for testing. Located in the middle of the Petri plate was a circular area of mycelium of 5 mm in length. This area contained a vigorously growing culture of the fungus being studied. The mycelial circle was consistently positioned at a fixed distance from the border of the plate. The plate was then incubated at a temperature of 30 °C for a period of three to seven days. The percentage of inhibition was calculated according to the procedure described by Maurhofer et al. [29]. The experiment was conducted with three repetitions. The data obtained were subjected to statistical analysis using analysis of variance (ANOVA) with the use of CoStat software. Tukey's HSD test was employed to distinguish the means at a significance level of p≤0.05.
Molecular characterization of the hopeful Bacillus isolate
After inoculating 5 milliliters of Luria-Bertani broth medium with the pure colony, the mixture was stirred at a temperature of 30°C for an entire night. Using the Wizard Genomic DNA Purification Kit (Promega, USA), bacterial genomic DNA was extracted from a 1 mL bacterial pellet. The procedure was carried out following the instructions provided by the manufacturer. A polymerase chain reaction (PCR) experiment was carried out by employing certain primers of 16s rRNA (F: AGAGTGATCCTGGCTCAG, R: GGTTACCTTGTTACGACTT), following the approach that was described earlier [30]. Following that, the PCR fragments were subjected to purification and sequencing protocols. Following the uploading of the annotated sequences to GenBank, a comparison study was carried out with the isolates that had been published in the past.
Experiment on Fermentation
Bioreactor (fermentor)
The B. amyloliquefaciens isolate KSAS6, which showed great potential, was grown and fermented in a ten-liter bench-top bioreactor (Cleaver, Saratoga, USA) equipped with four baffles and turbine impellers that had six-bladed discs. A computerized control system was employed to supervise the batch fermentation process with a working capacity of four liters. An automated method was controlled using a device equipped with a 10.4-inch color touch-screen interface. This device can store up to 59,994 distinct programs for various scenarios. The batch fermentation process was conducted at a consistent temperature of 30°C and a pH level of 7, which was carefully maintained. For pH regulation, a solution of 2 mol L–1 NaOH and a solution of 2 mol L–1 HCl were automatically supplied. Once the air passed through a sterile filter, it was compressed and adjusted to a flow rate of 0.5 VVM (air volume per broth volume per minute). The agitation speed was initially set at 200 rpm and then manually increased to maintain the dissolved oxygen (DO) level above 20%. The DO content and pH were measured using METTLER TOLEDO electrodes [31].
Batch fermentation process
The batch fermentation process is a method of producing a desired product by allowing microorganisms to grow and metabolize in a controlled environment for a specific time. The highly potential B. amyloliquefaciens isolate KSAS6 was inoculated as a single colony in a 500 mL-1 Erlenmeyer flask containing 100 mL-1 of Number 3 medium [32]. The cell broth was thereafter incubated overnight at a temperature of 30°C with continuous agitation at a speed of two hundred revolutions per minute. The batch fermentation process was initiated in a stirred tank bioreactor with an optical density (OD550) of 0.5. The bioreactor had a working volume of 4 liters and was filled with Number 3 medium. During the fermentation process, many culture samples were collected and the quantities of biomass and glucose were measured in each sample.
Analytical methods
Assessment of cell biomass
After the culture's optical density (OD) at 550 nm was measured using a spectrophotometer to track the cell count. After subjecting a sample of 10 mL-1 to centrifugation with a force of 894 g for 10 minutes, the mass of the cells was restored, purified, and then subjected to centrifugation again to determine the weight of the cells after they had been dried. Subsequently, the pellet was subjected to a drying process for the entire night at a temperature of 80°C in a dry-air oven, as described by Van Dam-Mieras et al. [33].
Measurement of glucose concentration
To determine the concentration of glucose, an enzymatic colorimetric kit was utilized. This methodology relies on the activities of peroxidase and glucose oxidase. The quinoneimine dye, which is used as a glucose concentration marker, transforms into a reddish-violet color as the final product.
Gas chromatography-mass spectroscopy (GC–MS) analysis
Through the use of GC–MS analysis, the bioactive components that were found in the cell-free supernatant of the promising B. amyloliquefaciens isolate KSAS6 were identified and described. To accomplish this, the culture filtrate was combined with ethyl acetate in a ratio of 1:1 by volume. According to Hirpara et al. [34], after the mixture had been thoroughly agitated for ten minutes and then left to sit for five minutes, two distinct layers that were not able to mix were formed. Through the utilization of a separating funnel, the top layer of the solvent, which included the biomolecules that were extracted, was separated separately. According to Sharma et al. [35], the ethyl acetate extract was further concentrated to a tiny volume by evaporating the solvent in a rotary evaporator while the vacuum was maintained. It was then that brown gum was shown after the ethyl acetate had evaporated. Following that, the crude extract was stored at a temperature of 4 degrees Celsius. An Agilent 7000D instrument (Santa Clara, California, United States) was used to perform GC–MS analysis on the residues that were produced, and the program conditions were programmed following Khamis et al. [36].
Results![]()
Fungal and bacterial isolates
Bacillus isolates were extracted from the rhizospheres of tomato, potato, and eggplant by the use of an isolation experiment that was carried out in the Kingdom of Saudi Arabia (KSA). In this particular investigation, fifteen different Bacillus isolates were Identified. Bacillus isolates BS1, BS2, BS3, and BS4 were initially a part of group 1, which was isolated from the rhizosphere of the tomato plant. Separated from the potato rhizosphere was group 2, which included BS5, BS6, BS7, BS8, BS9 and BS10. This group was referred to as the "second group. Finally, group 3 was responsible for the extraction of BS11, BS14, BS16, BS18, and BS20 from the rhizosphere of the eggplant.
Antagonism test of Bacillus isolates
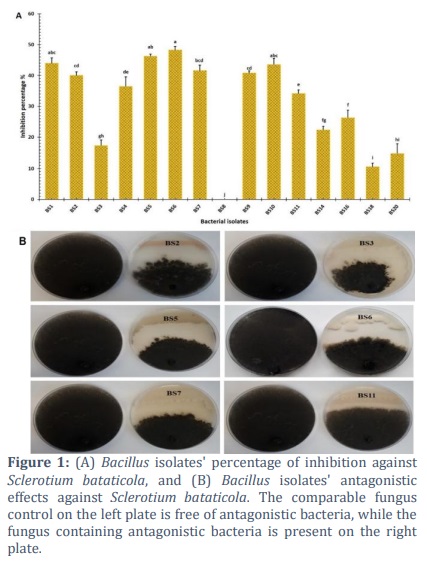
To determine whether or not they could be used as biological control agents, fifteen different Bacillus isolates were examined for their ability to inhibit the growth of S. bataticola in vitro. Based on their antagonistic activity against S. bataticola, it was demonstrated that there were significant differences between the Bacillus isolates. Isolate BS6 was more successful than other isolates in suppressing the growth of fungal mycelia caused by S. bataticola. It also had the highest inhibition percentage against the pathogen, which reached 48.2%. The next highest percentage of inhibition against the pathogen was in bacterial isolates BS5, which reached 46.3%. Both Bacillus isolates BS1 and BS10, which did not exhibit any significant differences amongst themselves, demonstrated antagonistic activity against S. bataticola that reached 43.9% and 43.5%, respectively. On the other hand, isolate BS8 did not exhibit any antagonistic action against the fungal pathogen (Figure 1).
Molecular characterization of the hopeful Bacillus isolate
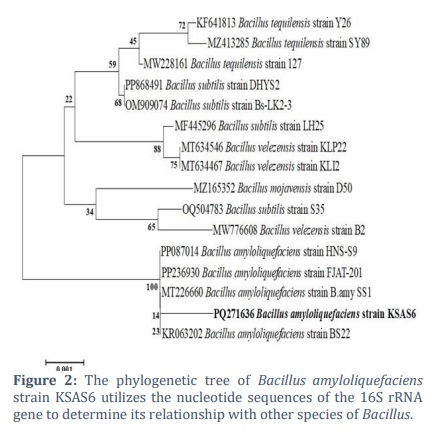
The phylogenetic tree analysis and NCBI-BLAST alignment identified the BS6 isolate as B. amyloliquefaciens. With the accession number PQ271636, the 1448 bp annotated nucleotide sequence has been uploaded to the GenBank database as B. amyloliquefaciens strain KSAS6. The nucleotide sequence exhibited a 100% sequence coverage with other GenBank-deposited B. amyloliquefaciens. Using the 16S rRNA nucleotide sequences, a phylogenetic tree was created for the B. amyloliquefaciens strain KSAS6 that was obtained in this work as well as other Bacillus species that were deposited in GenBank. Two main clusters were identified by the phylogenetic tree. The B. amyloliquefaciens strain KSAS6 (PQ271636), which was obtained for this investigation, and other strains of the same species that were deposited in GenBank with a 100% similarity percentage were included in Cluster 1. Bacillus species deposited in the GenBank database were included in cluster two. This cluster was further separated into two sub-clusters. Bacillus species strains B. velezensis, B. subtilis, and B. mojavensis were included in the first sub-cluster. Two groups were created from the second sub-cluster: the first group contained two strains of B. velezensis and one strain of B. subtilis, with an 88% similarity percentage. Two sub-groups were formed out of the second group. Two B. subtilis strains were found in the first sub-group, while three B. tequilensis strains were found in the second sub-group (Figure 2).
Fermentation experiments
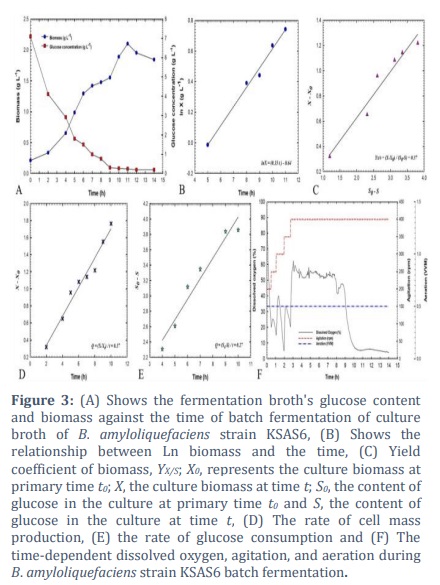
The initiation of batch fermentation of isolate KSAS6 of B. amyloliquefaciens was carried out with the assistance of a stirred engine bioreactor. It was observed that this particular strain exhibited a greater proportion of inhibition against the plant pathogenic fungus known as S. bataticola. For the batch fermentation of B. amyloliquefaciens culture broth, a plot was created that showed the concentration of biomass and glucose versus the time that had passed (Figure 3A). The maximum amount of biomass was observed at 2.1 g L-1 after 11 hours had passed. During the exponential stage, the logarithmic relationship was applied to obtain the constant specific growth rate (µ) of 0.13 h-1. This resulted in an exponential increase in cell mass over time, as depicted in Figure 3B. The natural logarithm (ln) of the biomass, which was calculated during the exponential phase, is depicted in the graph that originated before it as a linear path on a semi-logarithm that illustrates the relationship between time and biomass. The glucose concentration declined rapidly, reaching 1.6 g L-1 after 6 hours and 0.2 g L-1 after 14 hours (Figure 3A). The yield coefficient, also known as YX/S, is an important metric that is assessed during the exponential stage of bacterial cell growth. This coefficient compares the amount of biomass that is achieved with the amount of glucose that is consumed as a carbon source. 0.37 grams of cells per gram of glucose was the biomass yield coefficient that was achieved by this batch fermentation process (Figure 3C). During the exponential phase, additional factors were also measured to ascertain the effectiveness of the bacterial culture development process that was carried out by the batch fermentation method. Several aspects were taken into consideration, including the rate of cell mass creation and the rate of carbon supply consumption (glucose). As shown in Figure 3D, the rate of biomass synthesis (Qx) was 0.17 g L-1 h-1, while the rate of glucose consumption (Qs) was 0.27 g L-1 h-1 (Figure 3E).
The levels of dissolved oxygen (DO) influence the growth of bacterial cells. This concentration is controlled by the rate of aeration and the speed of agitation. Initiating the batch fermentation of B. amyloliquefaciens isolate KSAS6 with an agitation speed of 200 revolutions per minute and a rate of aeration equal to 0.5 VVM was done with a higher value of DO equal to 88.4%. During the first hour of the batch fermentation process, there was a significant decrease in the proportion of dissolved oxygen. As a result of the growing culture's increased need for oxygen, there was a fast decrease in the amount of dissolved oxygen, which reached 20% after half an hour. It is believed that a decrease in the amount of dissolved oxygen is a sign that the bacterial culture is developing and utilizing the glucose that is present in the culture broth throughout the growth process. It was possible to keep the oxygen level at or above 20% by adjusting the speed of the agitation, which ranged from 200 to 400 revolutions per minute. Following an increase of 2.7 hours, the dissolved oxygen level reached 62.4% at 3.05 hours, and then it rapidly decreased until it reached 20.3% at 9.35 hours (Figure 3F).
Identification of Bioactive Metabolites of Bacillus amyloliquefaciens
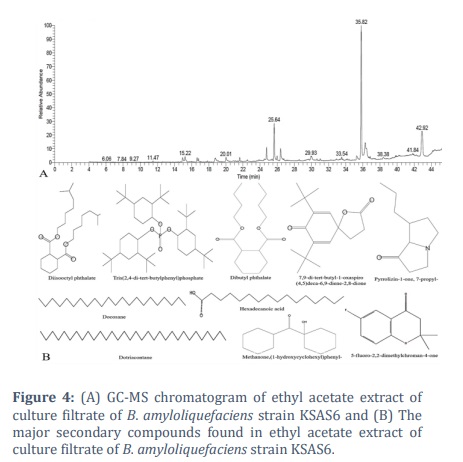
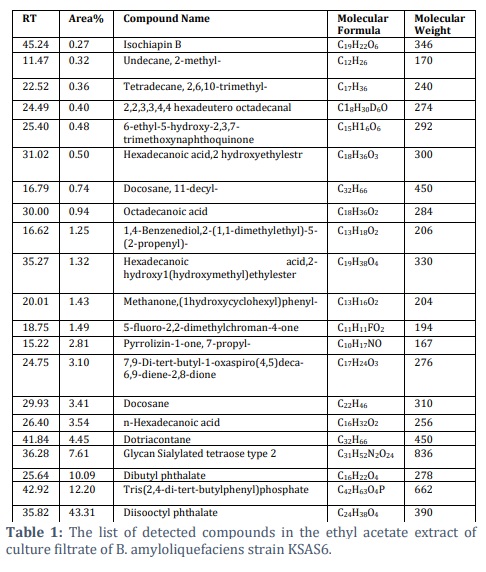
Extract of KSAS6 culture filtrate, which was achieved by ethyl acetate, was analyzed using GC-MS analysis to identify the specific compounds that could be responsible for its antagonistic effect against S. bataticola (Figure 4). The GC-MS profile showed the existence of twenty-one different compounds in the culture filtrate of KSAS6 (Table 1). These compounds including Isochiapin B (0.27%), Undecane, 2-methyl- (0.32%), Tetradecane, 2,6,10-trimethyl- (0.36%), 2,2,3,3,4,4 hexadeutero octadecanal (0.40%), 6-ethyl-5-hydroxy-2,3,7 trimethoxynaphthoquinone (0.48%), Hexadecanoic acid, 2-hydroxyethylester (0.50%), Docosane, 11-decyl- (0.74%), Octadecanoic acid (0.94%), 1,4-Benzenediol,2-(1,1-dimethylethyl)-5-(2-propenyl)- (1.25%), Hexadecanoic acid,2-hydroxy-1 (hydroxymethyl)ethylester (1.23%), Methanone,(1-hydroxycyclohexyl)phenyl- (1.43%), 5-fluoro-2,2-dimethylchroman-4-one (1.49%), Pyrrolizin-1-one, 7-propyl- (2.81%), 7,9-Di-tert-butyl-1-oxaspiro(4,5)deca-6,9-diene-2,8-dione (3.10%), Docosane (3.41%), n-Hexadecanoic acid (3.51%), Dotriacontane (4.45%), Glycan Sialylated tetraose type 2 (7.61%), Dibutyl phthalate (10.09%), Tris(2,4-di-tert-butylphenyl)phosphate (12.2%) and Diisooctyl phthalate (43.31%).
Figures & Tables
The ecological balance of soil-dwelling microbes has been upset by the widespread use of chemicals (pesticides) to manage plant diseases [37]. This has resulted in the emergence of resistant pathogen strains, contaminate groundwater, and pose clear health risks to humans [38]. The discovery of environmentally acceptable substitutes for the chemical pesticides now in use to address a variety of crop diseases is one of the major ecological concerns facing plant pathologists and microbiologists in the future [39]. Utilizing microorganisms for biological control can be a highly efficient method to reduce the negative impacts of synthetic chemical usage on the environment and decrease pollution and disturbances [40–42].
Bacillus sp. uses between 5 and 8% of its genome to synthesize bioactive secondary metabolites, which are antagonistic compounds meant to inhibit infections. Bacillus species use a broad range of antagonistic chemicals, such as polyketide compounds, bacteriocins, siderophores, and non-ribosomally produced peptides and lipopeptides, to inhibit pathogens in the root environment [43]. The most researched Bacillus species for their capacity to produce volatile organic compounds (VOCs) include B. amyloliquefaciens, B. velezensis, B. subtilis, and B. altitudinis [28,44]. B. subtilis and B. amyloliquefaciens are the producers of volatile organic compounds (VOCs), which include 2,3-butanediol. According to Saraf et al. [45], these are linked to the biocontrol of diseases and cause plants to develop induced systemic resistance (ISR). Extracellular enzymes including chitinase, β-1,3-and β-1,4-glucanases, laminarinase, oxalate oxidase, cellulases, proteases, phytases, and lipases are produced by Bacillus species. These enzymes have the ability to break down the components of fungi's cell walls, which may be a defense against fungus-related infections [46]. In this investigation, we evaluated fifteen Bacillus isolates as potential biocontrol agents against the in vitro mycelial growth of S. bataticola. When it came to S. bataticola mycelia inhibition, KSAS6 fared better than the other Bacillus isolates, with an inhibition percentage of 48.2%. The bioagent KSAS6's antagonistic effects can be attributed to its excretion of compounds and enzymes with both direct and indirect antifungal effects. Three Bacillus strains that exhibited antifungal activity against S. bataticola with an inhibition percentage of 64% were identified by Sicuia et al. [47]. This activity relates to their capacity to synthesize lipopeptides. According to Boiu-Sicuia and Cornea [48], some Bacillus strains isolated from various vegetables suppressed the fungal mycelial of S. bataticola in vitro with an inhibitory percentage equivalent to 56.7%. Additionally, the microscopic examination of S. bataticola's impeded mycelial growth demonstrated cytoplasmic content leaks, fungal perforations, and ulcerations in the cells.
The optimal circumstances for microbe development, including agitation, aeration, pH, and temperature, can be achieved in a stirred tank bioreactor. These factors are critical for optimizing the release of secondary metabolites and the proliferation of bacterial cells. In the current study and during the exponential phase, the cell mass of B. amyloliquefaciens isolate KSAS6 expanded exponentially at a steady specific growth rate of 0.13 h–1, which reached its maximum value of 2.1 g L-1 at 11 hours, with a production rate of 0.17 g L-1 h-1. The cell mass of B. subtilis isolate G-GANA7 increased exponentially with a constant specific growth rate of 0.3 h-1 [31]. On the other hand, the cell mass of B. velezensis strain GB1 was cultivated in a stirred tank bioreactor with a specific growth rate of 0.1 h-1 during a batch fermentation approach [49]. Glucose is one of the substrates that is most commonly used as a carbon source in various growing techniques. Since substrate inhibition is known to happen at specific doses. Thus, it is essential to ascertain the proper medium composition to facilitate the growth of a particular type of bacteria [50]. The glucose concentration was employed at its optimal value by the authors of this article for these reasons. After 14 hours, the glucose concentration reached 0.2 g. In this study, the biomass yield coefficient of the B. amyloliquefaciens isolate KSAS6 was 0.37 g cell/g glucose. The exponential phase of the bacterial growth curve for the B. amyloliquefaciens isolate KSAS6 revealed that the bacteria were expanding swiftly and consuming a significant amount of oxygen and carbon sources to do so, with a rate of glucose intake of 0.27 g L-1 h-1. Our findings indicate that during the first hour of the culture's growth, DO progressively dropped. Then, once the exponential phase began, the DO rapidly decreased, necessitating an increase in agitation speed to keep it above 20%. Stojanović et al. [51] showed that in B. subtilis cultivations, the generation of biomass and spores is significantly influenced by the agitation and aeration of the cultures, which have a direct impact on the DO concentration. B. subtilis did not sporulate when exposed to low oxygen levels, although it did continue to produce bioactive chemicals [52]. Numerous studies have found that increasing agitation and aeration rates or regulating the DO content over 20–30% can result in quicker cell development [53–55]. Nevertheless, higher agitation levels cause shear stress on cells, which can have detrimental consequences such as sporulation mechanism deactivation, uneven cell populations, smaller cells, and even cell death [56].
Microbial volatile organic compounds, often known as MVOCs, are a class of molecules that are generated as a result of metabolic activities performed by fungi and bacteria. There are many different chemical forms that volatile organic compounds (VOCs) can take, including ketones, alcohols, terpenoids, sulfur compounds, alkenes, and many others. Certain chemicals are shared by all of the microbes in the group, while other molecules are unique to specific strains. According to Sidorova et al. [57], a single bacterium is capable of producing up to one hundred different volatile organic compounds. According to, Bacillus species are believed to be factories of plant protective VOCs and play a variety of roles in the protection of plants against bacterial and fungal diseases [58]. Antimicrobial VOCs have been reported to be produced by B. amyloliquefaciens strains in several different studies. According to Yu et al. [59], VOCs produced by the B. amyloliquefaciens strain have the potential to impede the mycelial development of Penicillium digitatum and P. italicum. It is possible to assert that all strains of B. amyloliquefaciens are capable of producing VOCs. However, the volume, type, chemical composition, relative abundance, and spectrum of inhibition differ among the strains. Furthermore, the spectrum of inhibition was influenced by the concentration and type of nutrients present in the medium [60]. In the current study, the identification of bioactive constituents in the ethyl acetate extract of B. amyloliquefaciens KSAS6 was conducted through the application of GC–MS analysis. GC-MS chromatogram of KSAS6 revealed that twenty-one VOCs compounds, which varied from aromatic and aliphatic compounds, were produced by B. amyloliquefaciens. The major compound was the aromatic compound, diisooctyl phthalate, which constitutes 43.31% of the total area followed by the aromatic compounds, tris(2,4-di-tert-butylphenyl)phosphate and dibutyl phthalate which contributed 12.20% and 10.09%, respectively. Additionally, the aromatic compounds, 7,9-Di-tert-butyl-1-oxaspiro(4,5)deca-6,9-diene-2,8-dione, Pyrrolizin-1-one, 7-propyl-, 5-fluoro-2,2-dimethylchroman-4-one and Methanone,(1-hydroxycyclohexyl)phenyl- were detected in varying proportion. On the other hand, Dotriacontane, n-hexadecanoic acid, and Docosane as aliphatic compounds were detected, constituting 4.45%, 3.54%, and 3.41% of the total area, respectively. The primary component of Penicillium sp.'s ethyl acetate extract, diisooctyl phthalate, was shown by Lykholat et al. [61] to be the cause of the extract's antifungal action against Alternaria alternata. It can prevent the formation of germs and possesses qualities that aid in wound healing [62,63]. Diisooctyl phthalate has recently been suggested by El-Enain et al. [64] as a possible medicinal agent for the treatment of severe bacterial infections. According to Gheda and Ismail [65], cyanobacteria can produce diisooctyl phthalate, which has antibacterial properties. Dibutyl phthalate suppressed the growth of the yeast Candida albicans and the gram-positive bacterium Staphylococcus aureus, as shown by Blazević et al. [66]. Additionally, it was demonstrated that dibutyl phthalate, which is produced by B. amyloliquefaciens and B. velezensis, has antifungal properties [67]. Meloidogyne incognita J2 was inhibited by dibutyl phthalate and diisobutyl phthalate [68]; Colletotrichum fragariae spore germination and hyphal growth were significantly inhibited by dibutyl phthalate [69]; and Verticillium dahliae Kleb proliferation was significantly suppressed by 5 mmol.L-1 dibutyl phthalate in the eggplant rhizosphere [70]. Tris (2,4-di-terbutylphenyl) phosphate is an antibiotic that has biological effects. It is a member of the phenol group. According to Valinluck et al. [71], the phenolic compounds can suppress the growth of P. chrysogenum, Fusarium oxysporum, and Aspergillus niger. Furthermore, according to Abdullah et al. [72], they have the ability to suppress the growth of E. coli, S. aureus, and Pseudomonas aeruginosa. B. tequilensis generated tris (2,4-di-terbutylphenyl) phosphate, which exhibited antibacterial action [73]. According to Amborabé et al. [74], phenolic chemicals have the ability to harm the cytoplasmic membrane, raise its permeability, and cause the discharge of intracellular fluids such proteins, nucleic acids, and inorganic ions. The antifungal action of B. amyloliquefaciens KSAS6 against S. bataticola may be attributed to these volatile organic molecules.
Finally, in terms of suppressing S. bataticola's mycelial development, the B. amyloliquefaciens stain KSAS6 was more effective than other isolates. During the batch fermentation of KSAS6 in the stirred tank bioreactor, the production of secondary metabolites and culture biomass was maximized to a satisfactory degree. The highest achievable level of biomass was 2.1 g L-1, and the yield coefficient was established at 0.37 g cells/g glucose. The GC-MS chromatogram of KSAS6 culture filtrate contained 21 distinct compounds. These compounds may be responsible for the antagonizing activity of B. amyloliquefaciens strain KSAS6 against S. bataticola. In conclusion, the B. amyloliquefaciens strain KSAS6 demonstrates significant antifungal effectiveness against S. bataticola. Consequently, it could serve as a potent biocontrol agent for managing plant fungal infections. However, additional studies and field trials are necessary to confirm results and identify potential limitations in practical agricultural applications, as well as to assess the effectiveness of this treatment on a larger scale.
Acknowledgement
The authors extend their appreciation to the Deputyship for Research & Innovation, “Ministry of Education” in Saudi Arabia for funding this research work through the project number (IFKSUDR_ F121).
Author Contributions
Conceptualization: Abdulaziz Al-Askar, Gaber Abo-Zaid, Ahmed Abdelkhalek; Data Curation: Fatimah Al-Otibi; Formal Analysis: Gaber Abo-Zaid, Ahmed Abdelkhalek; Methodology: Gaber Abo-Zaid, Ahmed Abdelkhalek; Abdulaziz Al-Askar; Project Administration: Abdulaziz Al-Askar, Fatimah Al-Otibi; Software: Mohamed Yassin; Supervision: Abdulaziz Al-Askar; Validation Abdulaziz Al-Askar, Fatimah Al-Otibi; Writing – Original Draft Preparation: Abdulaziz Al-Askar, Fatimah Al-Otibi, Mohamed Yassin, Gaber Abo-Zaid, Ahmed Abdelkhalek; ; Writing – Review & Editing: Gaber Abo-Zaid, Ahmed Abdelkhalek. All authors reviewed the manuscript.
The author declares no conflicts of interest.
![]() References
References
- El-Kazzaz MK, Ghoneim KE, Agha MKM, Helmy A, Behiry SI, Abdelkhalek A, et al. Suppression of Pepper Root Rot and Wilt Diseases Caused by Rhizoctonia solani and Fusarium oxysporum. Life, (2022); 12(4): 587.
- Jain A, Sarsaiya S, Wu Q, Lu Y, Shi J. A review of plant leaf fungal diseases and its environment speciation. Bioengineered, (2019); 10(1): 409–424.
- Wolfgang A, Taffner J, Guimarães RA, Coyne D, Berg G. Novel strategies for soil-borne diseases: Exploiting the microbiome and volatile-based mechanisms toward controlling Meloidogyne-based disease complexes. Frontiers in Microbiology, (2019); 10(JUN): 1296.
- Lodha S, Mawar R. Population dynamics of Macrophomina phaseolina in relation to disease management: A review. Journal of Phytopathology, (2020); 168(1): 1–17.
- Abdelkhalek A, Aseel DG, Király L, Künstler A, Moawad H, Al-Askar AA. Induction of Systemic Resistance to Tobacco mosaic virus in Tomato through Foliar Application of Bacillus amyloliquefaciens Strain TBorg1 Culture Filtrate. Viruses, (2022); 14(8): 1830.
- Yadav RC, Sharma SK, Varma A, Singh UB, Kumar A, Bhupenchandra I, et al. Zinc-solubilizing Bacillus spp. in conjunction with chemical fertilizers enhance growth, yield, nutrient content, and zinc biofortification in wheat crop. Frontiers in Microbiology, (2023); 14: 1210938.
- Kloepper JW, Lifshitz R, Zablotowicz RM. Free-living bacterial inocula for enhancing crop productivity. Trends in biotechnology, (1989); 7(2): 39–44.
- Shao J, Xu Z, Zhang N, Shen Q, Zhang R. Contribution of indole-3-acetic acid in the plant growth promotion by the rhizospheric strain Bacillus amyloliquefaciens SQR9. Biology and Fertility of Soils, (2015); 51: 321–330.
- Raguchander T, Jayashree K, Samiyappan R. Management of Fusarium Wilt of Banana using Antagonistic Microorganisms. Journal of Biological Control, (1997); 11: 101–105.
- Foysal MJ, Lisa AK. Isolation and characterization of Bacillus sp. strain BC01 from soil displaying potent antagonistic activity against plant and fish pathogenic fungi and bacteria. Journal of Genetic Engineering and Biotechnology, (2018); 16(2): 387–392.
- Fendrihan S, Constantinescu F, Sicuia OA, Dinu S. Beneficial Bacillus strains improve plant resistance to phytopathogens: a review. International Journal of Environment, Agriculture and Biotechnology, (2016); 1(2): 238512.
- Khan N, Martínez-Hidalgo P, Ice TA, Maymon M, Humm EA, Nejat N, et al. Antifungal activity of Bacillus species against Fusarium and analysis of the potential mechanisms used in biocontrol. Frontiers in microbiology, (2018); 9: 2363.
- Arguelles-Arias A, Ongena M, Halimi B, Lara Y, Brans A, Joris B, et al. Bacillus amyloliquefaciens GA1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens. Microbial cell factories, (2009); 8: 1–12.
- Kiesewalter HT, Lozano-Andrade CN, Wibowo M, Strube ML, Maróti G, Snyder D, et al. Genomic and chemical diversity of Bacillus subtilis secondary metabolites against plant pathogenic fungi. Msystems, (2021); 6(1): 10–1128.
- Amanullah A, Otero JM, Mikola M, Hsu A, Zhang J, Aunins J, et al. Novel micro‐bioreactor high throughput technology for cell culture process development: Reproducibility and scalability assessment of fed‐batch CHO cultures. Biotechnology and bioengineering, (2010); 106(1): 57–67.
- Bareither R, Bargh N, Oakeshott R, Watts K, Pollard D. Automated disposable small scale reactor for high throughput bioprocess development: a proof of concept study. Biotechnology and bioengineering, (2013); 110(12): 3126–3138.
- Krishna C, Nokes SE. Predicting vegetative inoculum performance to maximize phytase production in solid-state fermentation using response surface methodology. Journal of Industrial Microbiology and Biotechnology, (2001); 26(3): 161–170.
- Gangadharan D, Sivaramakrishnan S, Nampoothiri KM, Pandey A. Solid culturing of bacillus amyloliquefaciens for alpha amylase production. Food Technology and Biotechnology, (2006); 44(2): 269–274.
- Hewitt CJ, Nienow AW. The scale‐up of microbial batch and fed‐batch fermentation processes. Advances in applied microbiology, (2007); 62: 105–135.
- Ruiz‐Sanchez J, Flores‐Bustamante ZR, Dendooven L, Favela‐Torres E, Soca‐Chafre G, Galindez‐Mayer J, et al. A comparative study of taxol production in liquid and solid‐state fermentation with Nigrospora sp. a fungus isolated from Taxus globosa. Journal of applied microbiology, (2010); 109(6): 2144–2150.
- Kumar S, Sharma N, Pathania S. Optimization of process parameters and scale-up of xylanase production from Bacillus Amyloliquifaciens Sh8 in a stirred tank bioreactor. Cellulose Chemistry and Technology, (2017); 51(5–6): 403–415.
- Allman T. Bioreactors: design, operation, and applications. In: Fermentation Microbiology and Biotechnology, Fourth Edition, CRC Press; 2018. p. 283–308.
- Kiebus-Rpaa A, Karcz J. Mass Transfer in Multiphase Mechanically Agitated Systems. In: El-Amin M (ed.) Mass Transfer in Multiphase Systems and its Applications, London, UK: InTech; 2011. p. 93–116.
- Liu S. Bioprocess engineering: kinetics, sustainability, and reactor design. Amsterdam, The Netherlands: Elsevier; 2020.
- Glick BR. Some Techniques to Elaborate Plant–Microbe Interactions: Beneficial Plant-Bacterial Interactions, Springer, Cham 2015, p 97-122.
- Hathi Z, Mettu S, Priya A, Athukoralalage S, Lam TN, Choudhury NR, et al. Methodological advances and challenges in probiotic bacteria production: Ongoing strategies and future perspectives. Biochemical Engineering Journal, (2021); 176: 108199.
- Sarmiento-López LG, López-Meyer M, Maldonado-Mendoza IE, Quiroz-Figueroa FR, Sepúlveda-Jiménez G, Rodríguez-Monroy M. Production of indole-3-acetic acid by Bacillus circulans E9 in a low-cost medium in a bioreactor. Journal of bioscience and bioengineering, (2022); 134(1): 21–28.
- Abdelkhalek A, Al-Askar AA, Elbeaino T, Moawad H, El-Gendi H. Protective and Curative Activities of Paenibacillus polymyxa against Zucchini yellow mosaic virus Infestation in Squash Plants. Biology, (2022); 11(8): 1150.
- Maurhofer M, Keel C, Haas D, Défago G. Influence of plant species on disease suppression by Pseudomonas fluorescens strain CHAO with enhanced antibiotic production. Plant pathology, (1995); 44(1): 40–50.
- Yassin Y, Aseel D, Khalil A, Abdel-Megeed A, Al-Askar A, Elbeaino T, et al. Foliar Application of Rhizobium leguminosarum bv. viciae Strain 33504-Borg201 Promotes Faba Bean Growth and Enhances Systemic Resistance Against Bean Yellow Mosaic Virus Infection. Current Microbiology, (2024); 81(8): 220.
- Moustafa HE, Abd-Elsalam HE, Abo-Zaid GA, Serour EA. as Biocontrol Agent, II. Biotechnology, (2009); 8(1): 35–43.
- Asaka O, Shoda M. Biocontrol of Rhizoctonia solani damping-off of tomato with Bacillus subtilis RB14. Applied and environmental microbiology, (1996); 62(11): 4081–4085.
- Van Dam-Mieras MCE, Jeu WH, Vries J, Currell BR, James JW, Leach CK, et al. Techniques used in bioproduct analysis. 1992.
- Hirpara DG, Gajera HP, Bhimani RD, Golakiya BA. The SRAP based molecular diversity related to antifungal and antioxidant bioactive constituents for biocontrol potentials of Trichoderma against Sclerotium rolfsii Scc. Current genetics, (2016); 62: 619–641.
- Sharma D, Pramanik A, Agrawal PK. Evaluation of bioactive secondary metabolites from endophytic fungus Pestalotiopsis neglecta BAB-5510 isolated from leaves of Cupressus torulosa D. Don. 3 Biotech, (2016); 6(2): 210.
- Khamis WM, Heflish AA, El-Messeiry S, Behiry SI, Al-Askar AA, Su Y, et al. Swietenia mahagoni Leaves Extract: Antifungal, Insecticidal, and Phytochemical Analysis. Separations, (2023); 10(5): 301.
- Muhammad M, Ahmad MW, Basit A, Ullah S, Mohamed HI, Nisar N, et al. Plant growth-promoting rhizobacteria and their applications and role in the management of soilborne diseases. In: Bacterial secondary metabolites, Elsevier; 2024. p. 59–82.
- Moawad H, Abd El-Rahim WM. Bioremediation of irrigation water contaminated with textile dyes. Fresenius Environmental Bulletin, (2003); 12(7): 786–792.
- Mnif I, Grau-Campistany A, Coronel-León J, Hammami I, Triki MA, Manresa A, et al. Purification and identification of Bacillus subtilis SPB1 lipopeptide biosurfactant exhibiting antifungal activity against Rhizoctonia bataticola and Rhizoctonia solani. Environmental Science and Pollution Research, (2016); 23: 6690–6699.
- Usta C. Microorganisms in biological pest control—a review (bacterial toxin application and effect of environmental factors). Current progress in biological research, (2013); 13: 287–317.
- Abdelkhalek A, El-Gendi H, Al-Askar AA, Maresca V, Moawad H, Elsharkawy MM, et al. Enhancing systemic resistance in faba bean (Vicia faba L.) to Bean yellow mosaic virus via soil application and foliar spray of nitrogen-fixing Rhizobium leguminosarum bv. viciae strain 33504-Alex1. Frontiers in Plant Science, (2022); 13: 933498.
- Elsharkawy MM, Sakran RM, Ahmad AA, Behiry SI, Abdelkhalek A, Hassan MM, et al. Induction of systemic resistance against sheath blight in rice by different Pseudomonas isolates. Life, (2022); 12(3): 349.
- Fira D, Dimkić I, Berić T, Lozo J, Stanković S. Biological control of plant pathogens by Bacillus species. Journal of biotechnology, (2018); 285: 44–55.
- Grahovac J, Pajčin I, Vlajkov V. Bacillus VOCs in the context of biological control. Antibiotics, (2023); 12(3): 581.
- Saraf M, Pandya U, Thakkar A. Role of allelochemicals in plant growth promoting rhizobacteria for biocontrol of phytopathogens. Microbiological research, (2014); 169(1): 18–29.
- Kumar P, Dubey RC, Maheshwari DK. Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiological research, (2012); 167(8): 493–499.
- Sicuia OA, Olteanu V, Ciuca M, Cîrstea DM, Cornea CP. Characterization of new Bacillus spp. isolates for antifungal properties and biosynthesis of lipopeptides. (2011); 54: 482–491.
- Boiu-Sicuia OA, Cornea CP. Bacterial strains involved in soilborne phytopathogens inhibition. Scientific Papers. Series A. Agronomy, (2021); 64: 641–646.
- Soliman A, Matar S, Abo-Zaid G. Production of Bacillus velezensis Strain GB1 as a biocontrol agent and its impact on Bemisia tabaci by inducing systemic resistance in a squash plant. Horticulturae, (2022); 8(6): 511.
- Ghasemi S, Ahmadzadeh M. Optimisation of a cost-effective culture medium for the large-scale production of Bacillus subtilis UTB96. Archives of Phytopathology and Plant Protection, (2013); 46(13): 1552–1563.
- Stamenković-Stojanović S, Karabegović I, Beškoski V, Nikolić N, Lazić M. Bacillus based microbial formulations: Optimization of the production process. Hemijska industrija, (2019); 73(3): 169–182.
- Carvalho ALU de, Oliveira FHPC de, Mariano R de LR, Gouveia ER, Souto-Maior AM. Growth, sporulation and production of bioactive compounds by Bacillus subtilis R14. Brazilian archives of biology and technology, (2010); 53: 643–652.
- Monteiro SM, Clemente JJ, Henriques AO, Gomes RJ, Carrondo MJ, Cunha AE. A procedure for high‐yield spore production by Bacillus s ubtilis. Biotechnology Progress, (2005); 21(4): 1026–1031.
- Cho JH, Kim YB, Kim EK. Optimization of culture media for Bacillus species by statistical experimental design methods. Korean Journal of Chemical Engineering, (2009); 26: 754–759.
- Posada-Uribe LF, Romero-Tabarez M, Villegas-Escobar V. Effect of medium components and culture conditions in Bacillus subtilis EA-CB0575 spore production. Bioprocess and biosystems engineering, (2015); 38: 1879–1888.
- Sahoo S, Rao KK, Suraishkumar GK. Reactive oxygen species induced by shear stress mediate cell death in Bacillus subtilis. Biotechnology and bioengineering, (2006); 94(1): 118–127.
- Sidorova DE, Plyuta VA, Padiy DA, Kupriyanova E V, Roshina N V, Koksharova OA, et al. The effect of volatile organic compounds on different organisms: Agrobacteria, plants and insects. Microorganisms, (2021); 10(1): 69.
- Poulaki EG, Tjamos SE. Bacillus species: factories of plant protective volatile organic compounds. Journal of Applied Microbiology, (2023); 134(3): lxad037.
- Yu SM, Oh BT, Lee YH. Biocontrol of green and blue molds in postharvest satsuma mandarin using Bacillus amyloliquefaciens JBC36. Biocontrol Science and Technology, (2012); 22(10): 1181–1197.
- Raza W, Wei Z, Ling N, Huang Q, Shen Q. Effect of organic fertilizers prepared from organic waste materials on the production of antibacterial volatile organic compounds by two biocontrol Bacillus amyloliquefaciens strains. Journal of Biotechnology, (2016); 227: 43–53.
- Lykholat YV, Khromykh NO, Didur OO, Drehval OA, Sklyar T V, Anishchenko AO. Chaenomeles speciosa fruit endophytic fungi isolation and characterization of their antimicrobial activity and the secondary metabolites composition. Beni-Suef University Journal of Basic and Applied Sciences, (2021); 10: 1–10.
- Amer MS, Barakat KM, Hassanein AEA. Phthalate derivatives from marine Penicillium decumbens and its synergetic effect against sepsis bacteria. Biointerface Research in Applied Chemistry, (2019); 9(4): 4070–4076.
- Huang L, Zhu X, Zhou S, Cheng Z, Shi K, Zhang C, et al. Phthalic acid esters: Natural sources and biological activities. Toxins, (2021); 13(7): 495.
- El-Enain A, Zeatar A, Zayed A, Elkhawaga M, Mahmoud Y. Diisooctyl Phthalate as A Secondary Metabolite from Actinomycete Inhabit Animal’s Dung with Promising Antimicrobial Activity. Egyptian Journal of Chemistry, (2023); 66(12): 261–277.
- Gheda SF, Ismail GA. Natural products from some soil cyanobacterial extracts with potent antimicrobial, antioxidant and cytotoxic activities. Anais da Academia Brasileira de Ciências, (2020); 92: e20190934.
- Blažević I, Radonić A, Mastelić J, Zekić M, Skočibušić M, Maravić A. Hedge mustard (Sisymbrium officinale): Chemical diversity of volatiles and their antimicrobial activity. Chemistry & biodiversity, (2010); 7(8): 2023–2034.
- Soliman SA, Khaleil MM, Metwally RA. Evaluation of the antifungal activity of Bacillus amyloliquefaciens and B. velezensis and characterization of the bioactive secondary metabolites produced against plant pathogenic fungi. Biology, (2022); 11(10): 1390.
- Yang G, Zhou B, Zhang X, Zhang Z, Wu Y, Zhang Y, et al. Effects of tomato root exudates on Meloidogyne incognita. PLoS One, (2016); 11(4): e0154675.
- Li X, Jing T, Zhou D, Zhang M, Qi D, Zang X, et al. Biocontrol efficacy and possible mechanism of Streptomyces sp. H4 against postharvest anthracnose caused by Colletotrichum fragariae on strawberry fruit. Postharvest Biology and Technology, (2021); 175: 111401.
- Yin YL, Zhou BL, Tang YP. Effects of three allelochemicals on Verticillium dahliae antagonistic microbes in eggplant rhizosphere. Allelopathy Journal, (2015); 35(1): 23–34.
- Varsha KK, Devendra L, Shilpa G, Priya S, Pandey A, Nampoothiri KM. 2, 4-Di-tert-butyl phenol as the antifungal, antioxidant bioactive purified from a newly isolated Lactococcus sp. International journal of food microbiology, (2015); 211: 44–50.
- Abdullah ASH, Mirghani MES, Jamal P. Antibacterial activity of Malaysian mango kernel. African Journal of Biotechnology, (2011); 10(81): 18739–18748.
- Murniasih T, Masteria Yunovilsa P, Untari F. Antibacterial activity and GC–MS based metabolite profiles of Indonesian marine Bacillus. Indonesian Journal of Pharmacy, (2022); 33: 475–483.
- Amborabé BE, Fleurat-Lessard P, Chollet JF, Roblin G. Antifungal effects of salicylic acid and other benzoic acid derivatives towards Eutypa lata: structure–activity relationship. Plant Physiology and Biochemistry, (2002); 40(12): 1051–1060.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()