

Full Length Research Article
Significance of miR-122 and miR- 150 in the pathogenicity of hepatitis B infection
Lamiaa Al-Maliki1*, Yasir W. Issa2, Shahlaa M. Salih1
Adv. life sci., vol. 12, no. 1, pp. 205-210, February 2025
*– Corresponding Author: Lamiaa Al-Maliki (Email: lamiaafingan@yahoo.com)
Authors' Affiliations
2. Department of Anesthesia Technology, MadenatAlelem University College, Baghdad – Iraq
[Date Received: 03/07/2023; Date Revised: 04/08/2024; Date Published: 31/12/2024]
Editorial Note: This manuscript has minor post-publication changes in the authors’ affiliation section.
Editorial Expression of Concern:
18 May 2025: Following publication of this paper, the internal audit (consequent to concerns on quality raised by Web of Science) notified Advancements in Life Sciences about problems in use of English language. By this Editorial Expression of Concern, we alert the scientific community as we address the errors.
Editorial Note:
31 May 2025: You are viewing the latest version of this article having minor corrections related to the use of English language and in references section. Expression of concern is hereby revoked.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: HBV, a contagious infection, causes chronic hepatitis, cirrhosis, and hepatocellular cancer in many people. The WHO estimates that 2 billion people are exposed to HBV annually, depending on when. The Hepadnaviridae family virus has an unfinished 3.2-kilobase double-stranded DNA molecule. Increased ALT, AST, and HBsAg positive or HBV DNA viral load are used to identify the illness. Despite global immunization campaigns, nations with low vaccination coverage and no diagnosis remain concerned. The worldwide HBV burden remains high. MicroRNAs (miRNAs) bind to mRNAs after transcription to govern gene expression, hepatitis B virus replication, and extracellular matrix development.
Methods: Sixty six Iraqi HBV patients were involved in a case-sectional control study. From September 2021 to February 2022, blood samples were taken from Al-Yarmook Teaching Hospital and Central Public Health Laboratory in Baghdad. ELISA verified HBV markers, HBsAg, HBsAb, HBeAg, HBeAb, and HBcAb in all samples, and real-time PCR estimated viral load from DNA. The research found that miR122 and miR150 gene expressions affect HBV fibrosis severity.
Results: The gene expression of miR-122 and miR-150 was found using RT-PCR after normalization with Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), a common housekeeping gene. The analysis of variance found substantial differences between patients and controls. The research found a considerable downregulation of miRNA-122 and miRNA-150 in HBV-infected individuals.
Conclusions: Depending on HBV severity, miRNA levels may vary. HBV patients, especially those with fibrosis and chronic HBV, have reduced miR122 and miR150. MiR-122 and miR-150 may be interesting HBV diagnostic, fibrosis progression, and therapeutic targets.
Keywords: Hepatitis B virus; MicroRNAs (miRNAs); RT-PCR; Primers; Gene expression; Liver disease
Introduction![]()
Hepatitis B is a virus-induced illness that impacts the liver and has the potential to cause liver complications in the short and long term, including cirrhosis, fibrosis, and liver cancer, particularly if contracted during childhood. In Iraq, the prevalence of HBV is around 1% [1, 2]. It is reported that 400 million people are currently infected with the virus and the majority of them are asymptomatic and unaware of their disease [3, 4]. During birth, the virus can be passed from mother to child (parental transmission) or through horizontal transmission, which occurs when a person is exposed to infected blood and body fluids like saliva, menstrual fluids, vaginal fluids, and seminal fluids. The disease is currently diagnosed by measuring the increase in liver enzymes including ALT, AST and HBsAg positive or HBV DNA viral load [5]. Although all of the inclusive programs of vaccination through the world, there is still some concern in the countries that has poor coverage of vaccination and were unable to diagnose it. Thus, HBV global burden remains in elevation [6]. HBV belongs to Hepadnaviridae family; the DNA is incomplete double strands of 3.2kb [7]. Liver fibrosis resulting from chronic damage of liver, which can occur due to viral infections, alcohol abuse and non-alcoholic Steatohepatitis. It is known that hepatic stellate cells (HSCs) are responsible for collagen production and accumulation of extracellular matrix proteins (ECM). HSCs undergoes trans differentiation after activation induced by inflammatory stimuli leads to collagen deposition in the liver tissue and results in liver fibrosis [8]. MicroRNA (miRNA) is small noncoding RNA sequence approximately 22nucleotides that was found to be a great biomarker in diagnosis of gene expression and regulation [9]. For hepatitis B virus liver disease, a large number of studies found that miRNA was able to control hepatitis B virus reproduction by managing the creation of external cellular structures [9]. MiRNAs, which are tiny pieces of DNA that are responsible for suppressing tumor genes, are also implicated in the advancement of chronic hepatitis, liver fibrosis, cirrhosis, and hepatocellular carcinoma resulting from hepatitis B virus infection [10, 11]. MiR-122 binds to specific pregenetic HBV RNA region, which is highly conserved region and bicistronic mRNA which encodes HBV polymerase and core protein. This will regulate HBV gene expression and replication [12]. MiRNA-122 is the most prevalent among hepatic miRNAs unique to the liver and accounts for 70% of all liver miRNAs [13]. Studies have shown that miRNA-122 plays a crucial role in regulating the growth of liver cells and malignant changes [14]. It can also substantially impact controlling metabolisms, lipids, hepatic cirrhosis, and changes in HBV replication [14]. Studies have demonstrated that miRNA-122 is able to suppress HBV replication. Research has revealed that miRNA-122 can inhibit the replication of HBV by binding to HBV sequences, indicating a correlation between miRNA-122 expression levels and HBV replication [15, 16]. It is also found that miRNA can be existed in serums and tissues of HBV infections related tumors [17, 18]. Studies confirmed that miRNA dysregulation is likely present in all stages of hepatocarcinogenesis. At the same time, miRNA plays a role in distinguishing between individuals who are healthy and those who have HCC [19, 20]. That can differentiate between benign and malignant tissues and their sub-type [21]. Diagnosis of hepatic tumors can also be done through analysis of serum, plasma, and urine, which is seen as a non-invasive way to assess how patients respond to treatments and their prognosis in the disease. The origin of variations in miRNA expression could complicate the development of accurate biomarker tools because of differences in geographical factors causing delays in the onset of HCC.Mir-150 inhibits the proliferation of hepatic stellate cells and the production of EMC proteins. Collagen type I and IV are the primary fibrous proteins found in the extracellular matrix [11]. Type I collagen, which is most abundant protein comprises 36% of the total collagens in ECM of healthy liver [18]. In contrast, type IV collagen is the most elevated collagen in liver fibrosis, which accounting less than 10% of total collagen in the normal liver [19, 20]. The aim of this study was to evaluate two types of miRNAs, the first one is miR-122 and the second one is miRNA-150 in patients with liver infection at different stage of HBV infection. The miRs gene expression was determined through RT-PCR following the conversion of RNA to cDNA using a particular stem loop primer that elongates the intended miRNA.
Methods![]()
In this study, a case sectional control study was performed on sixty-six Iraqi patients infected with HBV was enrolled. Blood samples were obtained from Al-Yarmook Teaching Hospital, Baghdad, Iraq, and Central Public Health Laboratory, Baghdad, Iraq starting from September- 2021 till February 2022.All samples were confirmed previously for HBV markers, HBsAg, HBsAb, HBeAg, HBeAb, and HBcAb by ELISA and DNA was extracted for estimation viral load by real time PCR. The patients were sub-grouped according to the FibroScan results into F1 (acute), F2 (chronic) and F3 (cirrhosis) their age was ranged 29-62 years old. For comparison, 30 healthy individuals were enrolled in this study and their age was matched to the patients.
Collection and preparation of samples
5 milliliters of venous blood were taken from each patient and control subject, under sterile conditions. The blood samples were divided into separate EDTA tubes for gene expression analysis. RNA extraction was carried out using Tirol Reagent from ZYMO Research and ReliaPrep™ RNA Miniprep Systems from Promega in the USA. The concentration and purity of RNA were assessed using the Quantus™ Fluorometer.
The primers utilized in this research
The miR-122 Stem-loop primer had the sequence5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCACTGG-3′, while the forward primer sequence was 5′ CGCGCTTCT CCC AAC CCT 3′.
The miR-150 Stem-loop primer sequence was5′GTCGTATCCAGTGCA-GGGTCCGAGGTATTCGCACTGGATACGACCAAACA3′, and forward primer 5′ CGC GCG TGG AGT GTG ACA 3. The reference gene was GAPDH (glyceraldehyde-3-phosphate dehydrogenase eNM_002046.5). The forward primer was 5'GACAGTCAGCCGCATCTTCT3', and amplification reverse primer was 5' GCG CCCAATACGACCAAATC3'.In our study, we used a universal reverse primer that matched the stem loop region at the 5′ end CGCTTCACGAATTTGCGTGTCAT 3′. The primers were assumed using online MiRbase (http://www.mirbase.org).
Regulation of gene expression by MicroRNAs (two processes)
BioLabs in England provided the ProtoScript II reverse transcriptase reagent Kit, the process involved in converting single-stranded RNA into cDNA for real-time PCR purposes was conducted. Each reverse transcription reaction included 5uL of 5X prime script reaction buffer along with 0.5 µL of stem-loop primer for micro-RNAs tubes and 10uL of the extracted total RNA for each microRNA and GAPDH gene tubes, in combination with 4.5 µL RNA/DNAase free water. The RT reaction was completed using a two-step real-time RT SaCycler-48 thermal cycler from Sacace in Italy set at a temperature of 60°C for 15 minutes, followed by deactivation at 85°C for 5 minutes, and then at 4°C.A quantitative real-time polymerase chain reaction (qPCR) was performed with the LunaScript RT Master Mix Kit(5X) from BioLabs in the United Kingdom. The RT-PCR mix contained 10 µL of the master mix, 0.5 µL of both forward and reverse primers, 5 µL of cDNA from each sample, and 4 µL of nuclease free water to reach a total volume of 20 µL. The GAPDH gene was used as the endogenous control. The qPCR was carried out with the following steps: 95°C for 5 min for polymerase activation, followed by 40 cycles of 95°C for 20 seconds for denaturation, 60°C for 20 seconds for annealing, and 72°C for 20 seconds for extension. Ultimately, a melting curve examination was performed to evaluate the properties of double stranded cDNA as it underwent denaturation cycles. The Ct value of the specific miRNAs under investigation was normalized in relation to the GAPDH reference gene, and the miRNA expression levels were established using a relative quantification approach employing the comparative Ct equation: Fold change = 2^-ΔΔCT where: ΔCT represents the difference in Ct values between the target gene and the reference gene, while ΔΔCT denotes the contrast in ΔCT values between the patient group and the control group.
Statistical analysis
The information was statistically evaluated using GraphPad prism 8 software designed for Windows. The findings were presented using basic statistical measures like the mean and standard error. Differences in averages were assessed through independent samples T-test. Any probability of 0.05 or less was considered significant.
Results![]()
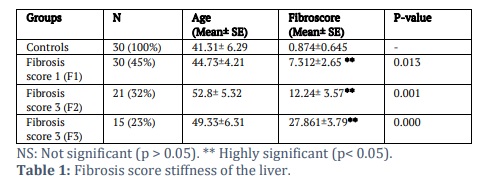
The result of FibroScan outcomes the fibrosis score stiffness of the liver, which is indicative of scarring, was interpreted in table (1). The patients were sub-grouped into F1, 30 (45%), F2 21 (32%), F3 15 (23%).
The gene expression of miR-122 and miR-150 were identified through the use of RT-PCR following adjustment with GAPDH. Analysis of variance showed that there were significant differences between patients and controls.
Gene expression of miR122
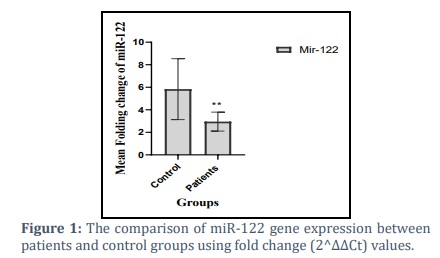
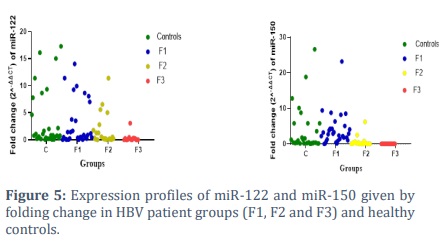
There was dramatically down regulated in the gene expression of microRNA-122 in HBV patients (2.951±0.8432), p<0.05 compared to controls (5.136±2.404) as shown in Figure (1), information were provided in supported materials table (1).
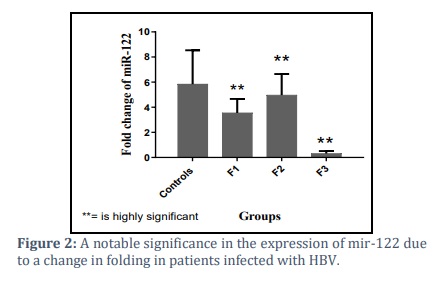
Analysis of the miR-122 expression data according to fibrosis scoring after adjustment with GAPDH exhibited fold change (2-∆∆Ct) level of mir-122was (3.549±1.114,4.964±1.680 and 0.3140±0.199 vs. 5.836±2.704), respectively, showing a notable decrease in mir-122 levels in the groups of patients as shown in figure (2)
Gene expression of miR150
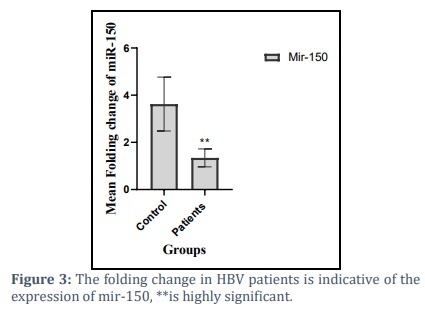
Analysis of variance showed that there were significant differences between patients and controls. There was a significant downregulation of microRNA-150 in HBV patients (1.3422±0.3817), p<0.05 compared to controls (3.624±1.141), as shown figure (3).
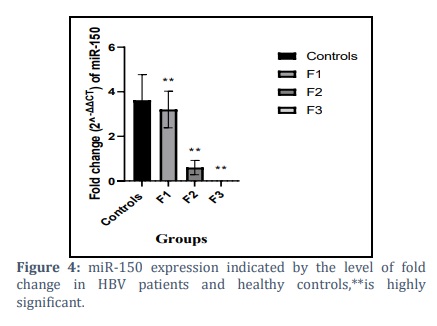
Normalization of miR-150 expression data with GAPDH as reference gene indicated a noticeable decrease in miR-150 levels in patient groups compared to controls. The fold changes were 3.205±0.8231, 0.6003±0.3213, and 0.002838±0.001 compared to controls at 3.624±1.141. as shown in figure (4).
Gene expression of both miR122 and miR150 to hepatic fibrosis grades
Results showed that a significant down regulation in gene expression of miR-122 and mir-150 in relation to hepatic fibrosis grades Figure (5).
Figures & Tables
MiRNAs in the liver are non-coding
Author Contributions
LF,Al-Maliki and SM,Salih conceived and designed the experiments. LF, Al-Maliki and YW, Issa performed the experiment and drafted the manuscript and data analysis. SM, Salih reviewed the manuscript. All authors read and approved the submission of the final version of the manuscript for publication.
The authors declare that there is no conflict of interestregarding the publication of this paper.
![]() References
References
- Lampertico P, Agarwal K, Berg T, Buti M, Janssen HL, et al. EASL 2017 Clinical Practice Guidelines on the management of hepatitis B virus infection. Journal of hepatology, (2017); 67(2): 370-398.
- Hou J, Liu Z, Gu F. Epidemiology and prevention of hepatitis B virus infection. International journal of medical sciences, (2005); 2(1): 50-57.
- Lavanchy D. Hepatitis B virus epidemiology, disease burden, treatment, and current and emerging prevention and control measures. Journal of viral hepatitis, (2004); 11(2): 97-107.
- Trépo C, Chan HL, Lok A. Hepatitis B virus infection. The Lancet, (2014); 384(9959): 2053-2063.
- Kim H, Lee S-A, Kim B-J. X region mutations of hepatitis B virus related to clinical severity. World journal of gastroenterology, (2016); 22(24): 5467-5478.
- Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nature reviews genetics, (2010); 11(9): 597-610.
- Friedman SL. Mechanisms of hepatic fibrogenesis. Gastroenterology, (2008); 134(6): 1655-1669.
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. cell, (2004); 116(2): 281-297.
- Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, et al. Widespread changes in protein synthesis induced by microRNAs. nature, (2008); 455(7209): 58-63.
- Sagnelli E, Potenza N, Onorato L, Sagnelli C, Coppola N, et al. Micro-RNAs in hepatitis B virus-related chronic liver diseases and hepatocellular carcinoma. World journal of hepatology, (2018); 10(9): 558-570.
- Kloosterman WP, Plasterk RH. The diverse functions of microRNAs in animal development and disease. Developmental cell, (2006); 11(4): 441-450.
- Zhang Y, Jia Y, Zheng R, Guo Y, Wang Y, et al. Plasma microRNA-122 as a biomarker for viral-, alcohol-, and chemical-related hepatic diseases. Clinical chemistry, (2010); 56(12): 1830-1838.
- Russo A, Potenza N. Antiviral effects of human microRNAs and conservation of their target sites. FEBS letters, (2011); 585(16): 2551-2555.
- Roderburg C, Urban GW, Bettermann K, Vucur M, Zimmermann H, et al. Micro‐RNA profiling reveals a role for miR‐29 in human and murine liver fibrosis. Hepatology, (2011); 53(1): 209-218.
- Zhang H, Li Q-Y, Guo Z-Z, Guan Y, Du J, et al. Serum levels of microRNAs can specifically predict liver injury of chronic hepatitis B. World journal of gastroenterology: WJG, (2012); 18(37): 5188-5196.
- Yu F, Zhou G, Li G, Chen B, Dong P, et al. Serum miR-181b is correlated with hepatitis B virus replication and disease progression in chronic hepatitis B patients. Digestive diseases and sciences, (2015); 602346-2352.
- Li L-M, Hu Z-B, Zhou Z-X, Chen X, Liu F-Y, et al. Serum microRNA profiles serve as novel biomarkers for HBV infection and diagnosis of HBV-positive hepatocarcinoma. Cancer research, (2010); 70(23): 9798-9807.
- Qi P, Cheng S-q, Wang H, Li N, Chen Y-f, et al. Serum microRNAs as biomarkers for hepatocellular carcinoma in Chinese patients with chronic hepatitis B virus infection. PloS one, (2011); 6(12): e28486.
- Bertino G, Ardiri AM, Calvagno GS, Bertino N, Boemi PM. Prognostic and diagnostic value of des-γ-carboxy prothrombin in liver cancer. Drug news & perspectives, (2010); 23(8): 498-508.
- Anwar SL, Lehmann U. MicroRNAs: emerging novel clinical biomarkers for hepatocellular carcinomas. Journal of clinical medicine, (2015); 4(8): 1631-1650.
- Szabo G, Bala S. MicroRNAs in liver disease. Nature reviews Gastroenterology & hepatology, (2013); 10(9): 542-552.
- Hu J, Xu Y, Hao J, Wang S, Li C, et al. MiR-122 in hepatic function and liver diseases. Protein & cell, (2012); 3364-371.
- Tsai W-C, Hsu S-D, Hsu C-S, Lai T-C, Chen S-J, et al. MicroRNA-122 plays a critical role in liver homeostasis and hepatocarcinogenesis. The Journal of clinical investigation, (2012); 122(8): 2884-2897.
- Li J, Ghazwani M, Zhang Y, Lu J, Li J, et al. miR-122 regulates collagen production via targeting hepatic stellate cells and suppressing P4HA1 expression. Journal of hepatology, (2013); 58(3): 522-528.
- Waidmann O, Bihrer V, Pleli T, Farnik H, Berger A, et al. Serum microRNA‐122 levels in different groups of patients with chronic hepatitis B virus infection. Journal of viral hepatitis, (2012); 19(2): e58-e65.
- Jopling CL, Yi M, Lancaster AM, Lemon SM, Sarnow P. Modulation of hepatitis C virus RNA abundance by a liver-specific MicroRNA. science, (2005); 309(5740): 1577-1581.
- Kumar S, Chawla YK, Ghosh S, Chakraborti A. Severity of Hepatitis C Virus (Genotype‐3) Infection Positively Correlates with Circulating MicroRNA‐122 in Patients Sera. Disease Markers, (2014); 2014(1): 435-476.
- Zheng J, Lin Z, Dong P, Lu Z, Gao S, et al. Activation of hepatic stellate cells is suppressed by microRNA-150. International journal of molecular medicine, (2013); 32(1): 17-24.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()