

Full Length Research Article
Androgen Receptor (AR) ablation encumbers the expansion and function of T regulatory cells (Treg) in a male mouse model
Saleh Altuwaijri*, Saleh Mohammed Albarrak
Adv. life sci., vol. 12, no. 1, pp. 197-204, February 2025
*– Corresponding Author: Abdulaziz Asiri (Email: amfasiri@ub.edu.sa)
Authors' Affiliations
[Date Received: 04/04/2024; Date Revised: 26/10/2024; Date Published: 31/12/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Many autoimmune diseases have a higher prevalence in females than males but there is no clear explanation for this phenomenon. Naturally occurring CD4+CD25+FOXP3+ Treg cells carry out a vital role in immune tolerance. We postulate that androgen may influence Treg cells differentiation and function.
Methods: Eight to twelve weeks old wild-type (WT) and androgen receptor knockout (ARKO) male mice (C57BL/6) were utilized. Treg cells in diverse lymphoid organs were separated with an easySep selection kit and sorted with FACSAria. Treg cells were phenotypically and functionally characterized by flow cytometry analysis (FACS) and an in vitro immune suppressive assay.
Results: Significantly Lower prevalence of thymic and peripheral Treg cells were noted in ARKO mice compared to the WT mice. Sorted ARKO Treg cells were functionally less suppressive than their counterparts in WT mice.
Conclusion: Our data suggest that the androgen receptor (AR) signaling pathway may be implicated in Treg cell expansion and function. To our knowledge, this study is the first to look into how AR knockout affects Treg cells. Therefore, it might flashlight the mechanisms of inflammatory and autoimmune disease.
Keywords: Androgen receptor; ARKO mice; Immunity; Immunosuppression; Treg cells.
Introduction![]()
Autoimmune illnesses in humans are more common in females than males [1]. This sexual dimorphism has also been found in the creation of infectious diseases [2, 3]. Immunotherapy response variations based on sex have been documented [4, 5]. Several reports indicated that the numbers of CD4+CD25+FOXP3+ T regulatory cells (Treg cells) that mediate marginal tolerance and defense against autoimmunity are significantly reduced in healthy females in comparison to healthy males [6]. A subset of T cells called regulatory T cells (Tregs) is necessary for maintaining immunological homeostasis and limiting uncontrolled immune reactions. Tregs exhibit certain cell surface markers, such as CD4, CD25, and the transcription factor FOXP3 [7]. Treg cells are used to block immunological reactions as they can inhibit effector T cells, B cells, and antigen-presenting cells from proliferating and performing their effector activities [8]. Other studies showed that lymphocytes express estrogen and androgen receptors (ER and AR) which might suggest that steroid sex hormones, estrogen, and androgen, could directly influence the functions of cells intricate in the immune response [8, 9]. In mice models, estradiol (E2) has been shown to increase the regulatory CD4+CD25+ compartment [10]. Similarly, the dual depletion of E2 and androgen resulted in a decline in CD4+CD25– T cells population including CD4+CD25+ Treg cells, and androgen replacement prevented this reduction in a rat model [11]. Clinical research revealed that patients with androgen insensitivity syndrome had a higher risk of developing asthma than control patients and that androgens and AR signaling improved lung function and reduced the symptoms of asthma [12].
Medical castration of males drastically lowers CD4+CD25+T cell counts in humans, with no effect on the ratio of CD4+ to CD8+ T cells and the percentages of CD4+ and CD8+T cells [13]. It’s interesting to note that multiple sclerosis patients have a favorable link between the use of steroid hormones like glucocorticoids and the proportion of Treg cells in the total CD4+ population [14]. Moreover, an increased number of CD4+ CD25+ T (Treg) cells have been increased in the peripheral blood of healthy and asthmatic pregnant women after systemic treatment with glucocorticoids [15]. These investigations support the potential role of steroids in the development of Treg cells and consequently the vulnerability to autoimmune diseases. However, the role of the androgen/ (AR) and Treg cells in this context is not clearly understood. Guan et al. [16] show that androgen receptor (AR) inhibition sensitizes tumor-bearing hosts to efficient checkpoint blockage by directly improving CD8 T cell activity [16]. In individuals with asthma, DHEA treatment and the rate of DHEA transformation into downstream androgens improved glucocorticoid responses and/or lung function [17]. An explanation for the sex variance in asthma was provided by Gandhi et al. [18], who demonstrated that AR signaling sustained Treg suppressive activity.
We showed in our former paper that depletion of AR in MSCs isolated from ARKO mice inhibits the induction of Treg cells via androgen TGF-B interaction [19]. In this study, we used the same mouse model to flash some light on the direct effects of androgens/AR depletion on the Treg cells.
Methods![]()
Animals and Ethical approval
The University Committee on Animal Resources of the University of Rochester approved all animal procedures. The current study used an ARKO mice model that was previously published. [20]. Floxed AR parents and cre parents mated to generate ARKO female mice. The B6129 mosaic strain, the chimera founder, was mated with the B6 female to produce female mice who were heterozygous for floxed AR. Male floxed AR mice were produced by the subsequent F2 offspring. A heterozygous KO of the AR female carrying the Cre was produced by breeding the heterozygous floxed AR female with the homozygous FVBN-TgN ACTB cre male that carries Cre under the -actin promoter. The heterozygous ARKO female carrying the Cre and the floxed AR male mated to produce homozygous ARKO Cre female mice in a 1:8 ratio. PCR was used to determine the genotype of WT and ARKO mice, as in our previously published studies [20, 21]. The Jackson Laboratory in Bar Harbor, Maine, USA, provided all the animals.
Peripheral blood mononuclear cell (PBMC) isolation
Half to one milliliter of blood was extracted via the facial vein (submandibular bleeding), placed in heparinized tubes, and centrifuged on Ficoll-Hypaque gradients (GE Healthcare Bioscience, USA). Following their recovery, the PBMC were counted using Trypan blue dye (Sigma Aldrich, USA), cleaned in AIM-V medium (Invitrogen Life Technologies, USA), and immediately used for experimentation.
Splenocytes isolation
Mice were euthanized by CO2 asphyxia, and their spleens were removed and weighed. The spleens were homogenized on top of a cell strainer in a cell culture dish using 5 ml of cold media (FACS wash), and were stored on ice until the cells were labeled with antibodies for phynotypic analysis using flow cytometry.
Bone marrow cells isolation
Bone marrow cells were collected from each animal's tibia and femur by washing the cavity of the marrow with RPMI 1640 medium (Life Technologies, Inc., Grand Island, NY) using a syringe equipped with a 26-gauge needle. To create single-cell suspensions, tissues were either homogenized using a powdered glass homogenizer or between the frosted edges of microscope slides. Following a ten-minute centrifugation, the cell suspensions were then added to a 50 ml conical tube, mixed with 5 ml of RBC lysis buffer (Sigma Aldrich, USA), and allowed to sit at room temperature for three minutes. Cells were then suspended at 5X106 ml-1 after being centrifuged twice with 15 ml of FACS wash buffer (350 g for 5 min).
Immune cells sorting
Following the guidelines provided by the manufacturer (Stem Cell Technologies, Cambridge, MA, USA), immune cells in the various organs were separated using EasySep selection kits (Cat #: 19852, 18782, and 18970 for CD4+, CD25+ and CD11b, respectively), and then they were sorted using a FACSAria Cell Sorter. The cells were rinsed with Fluorescence-activated Cell Sorting (FACS) buffer (PBS, 1% heat-inactivated fetal bovine serum +0.1% sodium azide). Following two FACS buffer washes, 1-3 x 106 cells were exposed to capture antibodies at 4 C for 30 min. If necessary, cells were given an additional 30 minutes of streptavidin-conjugated fluorochrome treatment at 4 C. Becton Dickinson and Co., Franklin Lakes, NJ, USA, used FACS CantoII Flow Cytometry to analyze the tagged cells after resuspending them in FACS buffer. For the data analysis, FlowJo V7.5.5 (Tree Star, Inc., Ashland, OR) was used.
Suppression assays
In a 96-well plate, CD4+CD25+ Treg cells from WT or ARKO mice were seeded alongside CD4+CD25- T cells and WT antigen-presenting cells (APC) from the peritoneal wash (CD11b+). 2.5 µg/ml anti-mouse CD3 was used to stimulate cells for 72 hours. In the last 16 hours, 1 Ci (methyl-3H) thymidine was supplied to the culture. Thymidine incorporation was used to measure T-cell proliferation [22].
Thymocyte isolation and phenotypic characterization
The protocol that was previously described by Gray et al. [23] was slightly adjusted for this procedure (Gray et al., 2008). Three PBS washes were used to remove fat and connective tissue from each thymus. Fine scissors were used to chop the thymus into pieces measuring about 1 mm3, which were then suspended in 10% fetal bovine serum (FBS)-containing RPMI 1640 medium and repeatedly stirred for 5 minutes before the pellet was left to settle. For collection, the supernatants were discarded, and pellets were incubated for 10 mins at 37°C with moderate agitation in 10% FBS RPMI 1640 media with 0.125% (wt/vol) collagenase D and 0.1% (wt/vol) deoxyribonuclease I (Roche, Indianapolis, IN). The supernatant was collected after each digestion, and the process was then carried out three more times. The remaining aggregates were incubated in RPMI 1640 medium with 0.125% Collagenase/Dispase (Roche) and 0.1% (wt/vol) deoxyribonuclease I for an additional 10 minutes. To separate the macroscopic fragments, the remaining thymic aggregates were periodically triturated with a 1 ml blue tip. The cells were then collected from all of the supernatant fractions, rotated at 450g for 5 mins, and then suspended in
chilled FACS buffer. A 70-m strainer was used to collect cells, which were then counted. 1-2 106 cells were tagged with the proper fluorescent antibodies to characterize the phenotype of thymic stromal cells.
Statistical analysis
GraphPad Prism version 10 was utilized for statistical analysis and curve sketching (Bangalore, 560035 Karnataka, India). The data were represented by the means of a minimum of three experiments plus one standard error of the mean (SE). The data was assessed using the unpaired Student's t-test with a significance level of α = 0.05.
Results![]()
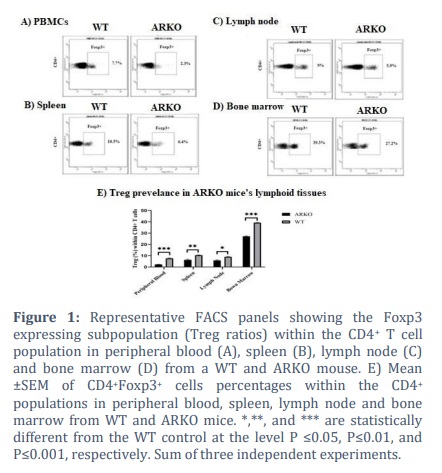
Evaluating Treg prevalence in the ARKO mice
Expression of the Treg cells marker, Foxp3+, in CD4+ T cells isolated from PBMCs, spleen, lymph node (LN), and bone marrow (BM) was examined using flow cytometry (FACS). The percentage of CD4+ cells expressing the Foxp3+ phenotype significantly declined (p ≤ 0.005) in the ARKO mice model compared to the WT mice in prepheral blood and in all the examined lymphoid organs as shown in Fig. 1 (2.31 vs 7.70% in peripheral blood, 6.38 vs 10.48% in the spleen, 5.88 vs 9.06% in the lymph nodes, and 27.18 vs 39.31% in the bone marrow).
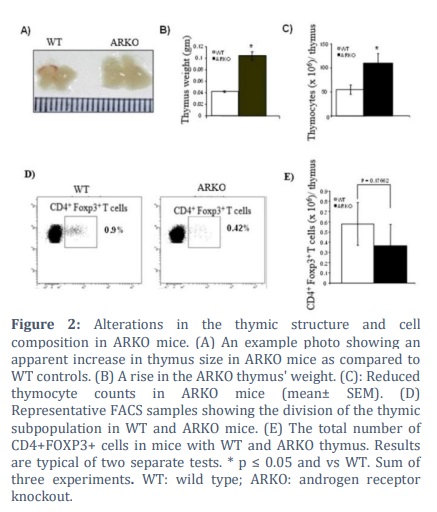
Next, we sought to determine if the decrease in Treg frequency in the ARKO mice's peripheral blood and lymphoid tissues was due to defects in thymic processes as Tregs typically originate in the thymus and sex hormones induce thymic hypocellularity [24]. Thus, we investigated the structure and composition of the thymus. The weights of the thymus in ARKO mice significantly increased (p ≤ 0.01) and were hypertrophied as shown in Fig. 2A and B. Moreover, thymus cellularity icreased by half on average (p ≤ 0.01) as shown in Fig. 2C. The percentages of CD4+Foxp3+ T (Treg) cells among thymic CD4+ T cells were notebly lower in the ARKO mice compared to the WT mice (Fig.2D). Likewise, the absolute numbers of CD4+FOXP3+ T cells per thymus were remarkably decreased in the ARKO mice compared to the WT mice (Fig. 2E).
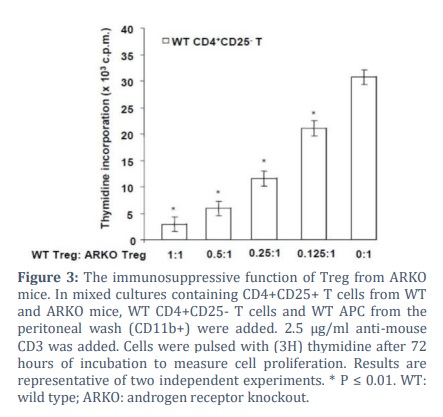
Evaluating the suppressive activities of Treg cells
Treg cells from pregnant women and Treg cells treated with E2 in suppression assays reduced the expansion of effector T cells [25]. These earlier findings suggest that androgen/AR and other steroid hormones may promote CD4+CD25+ Foxp3+ Treg cell suppression activities. To test this observation, we performed an immunosuppression assay to compare the immunosuppressive properties of ARKO Treg cells to their WT counterparts. In anti CD3 stimulated mixed cultures containing WT CD4+CD25– T cells and WT APC from the peritoneal wash (CD11b+), WT but not ARKO Treg cells exhibited a dose-dependent suppressive effect. ARKO Treg cells were less suppressive than their counterparts in the WT mice (Fig. 3).
Figures & Tables
Immune tolerance is greatly influenced by the function of normally existing CD4+CD25+FOXP3+ Treg cells. We hypothesize that androgen/AR may affect the development and functionality of Treg cells. Steroid hormones influence metabolism, physiology, and sexual development in a variety of ways. The immunomodulatory properties of androgens are well-recognized [26]. It has been suggested that male gonads affect the thymus’ involution. Thymic involution, a condition that occurs during adolescence, is the process by which the thymus gradually shrinks after reaching its maximum size at puberty. When mice were given androgens, the size of the thymus was quickly reduced. Castration ceases the thymic involution. Additionally, it is widely known that sex bias influences the frequency of autoimmune and allergy illnesses [27]. Steroid hormones also have a role in the circadian regulation of the immune system since the biological clock regulates their plasma levels [28]. Testicular feminization AR-mutant (ARtfm) mice exhibited thymus hypertrophy and androgen inconsiderateness, according to a research by Wilhelmson et al. [29]. These characteristics were unaffected by the transplanting of healthy bone marrow cells into these animals. On the other hand, typical thymus shrinkage occurred when ARtfm bone marrow cells were transplanted into control animals along with androgen therapy. They concluded that AR in thymic stromal cells is a factor in thymic atrophy. Additionally, AR deficient animals exhibit thymus expansion that is unique to thymic epithelial cells but not T cells. These findings imply that thymic involution is encouraged by androgen/AR signaling via thymic epithelial cells [30]. Androgens have pleiotropic effects on the immune system and increase the suppressive activity of Tregs, which reduces inflammation [30]. As a result, it is probable that the AR has an immuno regulatory activity. Tregs are thought to be the most numerous and important cells that provide peripheral immunological tolerance and homeostasis, according to Diky and Rudensky [31]. Treg cells are crucial for maintaining self-tolerance, regulating misdirected immune responses, and preventing too aggressive immune responses [32]. Interestingly, deficiencies in Treg cell function and/or quantity have been identified in the majority of autoimmune diseases and conditions related to chronic inflammation, including rheumatoid arthritis, multiple sclerosis, diabetes, etc. [33]. Therefore, improving regulatory T cell activity has tremendous therapeutic promise [34]. Hence, it can be considered the first-line therapy for some autoimmune diseases and malignant cells, and results in apoptosis of the cells of primary tumor. However, castration-resistant illness typically recurs years after the main tumor has been treated. Therefore, researching the long-term impact of androgen removal on the immune system will aid in the development of more effective immunological therapies for antineoplastic immune remedies.
To separate the direct effect of androgen ablation on Treg cells expansion, we evaluated CD4+ FOXP3+ T-cells in ARKO mouse. Since the thymus is the main place of Treg expansion, we directly examined thymic Treg development in an androgen ablation mouse model, and we show that ARKO mice had a significantly reduced number of Tregs produced in their thymus. It's probable that a change in the kinetics of differentiation in the thymus of ARKO caused relatively lower Treg populations.
In this study, we found that the ARKO mice's bone marrow had significantly lower numbers of Treg cells compared to the WT mice. A FOXP3 bicistronic reporter knock-in mouse model verified this observation [35]. However, it is unknown how pathological circumstances affect bone marrow Treg cells. We have noticed less bone marrow Treg cells in ARKO mice. This observation was confirmed by the reduced prevelance of Treg cells in PBMC of the ARKO mice. Although it is generally accepted that Treg cells and conventional T cells migrate into lymphoid organs, it is unclear if they use the same mechanisms [36]. In this investigation, we report lower percentages of Tregs in the spleen and LNs of ARKO mice. Many mechanisms thought to contribute to Treg growth in the periphery have been proposed. Compared to the WT, we report less Tregs accumulations in the studied organs of the ARKO mice. This scarcity in peripheral T reg cell numbers in the ARKO mice suggest that the AR may act as a survival signal. Howvere, more experiments are needed to examine whether defect in the thymus reduced the numbers of T regs and diminished thier suppressive functions in the ARKO mice model.
The current investigation demonstrated that ARKO Treg cells were less suppressive than their counterparts in WT mice. Treg cells from pregnant women and Treg cells treated with E2 reduced the expansion of effector T cells in suppression assays [25]. These earlier findings suggest that androgen/AR and other steroid hormones may promote CD4+CD25+ Foxp3+ Treg cell suppression activities. To test this observation, we performed an immunosuppression assay to compare the immunosuppressive properties of ARKO Treg cells to their WT counterparts. Both WT and ARKO Treg cells exhibited a dose-dependent suppressive effect. However, ARKO Treg cells were less suppressive than their counterparts in the WT mice which comes in line with former in vitro experiments that Dihydrotestosterone (DHT)-treated CD4+ T lymphocyte showed elevated IL-10 production [36]. Interestingly, targeting IL-10 in Treg might be responsible for the blockade of suppressive activity of Treg cells as compared to WT Treg [38]. Therefore, it might be expected that androgen ablation will impede suppressive functions of Treg cells.
Tregs were once referred to as suppressor T cells because of their capacity to reduce autoimmune reactions. Although there are numerous distinct Treg subtypes, the major and extensively studied subtype that expresses the master transcriptional factor forkhead box P3 (FoxP3) and significant quantities of the CD25 cell surface marker [24]. Tregs are produced in the thymus during T cell development and are then referred to as thymus-derived Tregs (Tregs) or naturally occurring Tregs (nTregs) [39]. All lymphoid organs contain mature Tregs, which can make up to 5–10% of all blood CD4+ T cells [40]. Additionally, circulating CD4+ T cells that react with auto-antigens can differentiate into FoxP3+ Tregs in the periphery (pTregs). Although tTregs and pTregs can be found in the same places, their differences are really small to distinguish between them [41]. Although FoxP3+ Tregs are the most stable Tregs, they can change into anergic T cells and lose own suppressive function under specific circumstances [42]. Although some inherent ability to alter their phenotype to the micro-environment cannot be discarded, the existence of Treg populations with different degrees of lineage commitment accounts for the adaptability of FoxP3+ Tregs [42, 43].
It's crucial for the therapeutic uses of FoxP3+ Tregs to comprehend the elements that affect their plasticity. A direct treatment strategy to control abnormal or excessive immune reactions is Adoptive Treg Cell Transfer (AtregCT) [44]. In AtregCT therapy, Tregs are removed from patients, multiplied ex vivo, and then donated (autologous) back to the patient [45]. Accordingly, it has been demonstrated that alloantigen-specific Tregs are most effective in regulating Graft-versus-host disease (GVHD) than polyclonal Tregs [46].
In addition to controlling thymic T lymphopoiesis (thymopoiesis) and producing recent thymic emigrants (RTEs), or premature T cells that leave the thymus and continue to develop into mature naive T cells in peripheral lymphoid tissues, androgens have a substantial impact on T cell homeostasis [47]. Wilhelmson et al. [29] investigated how the androgen target cell affected thymopoiesis and RTEs in the spleen and lymph nodes. They examined how androgens/AR affected T cell homeostasis using AR knockout animal models. Rodrigues et al. provided more evidence that full androgen restriction alters thymus cellularity by boosting the quantity of bone marrow-derived thymus-seeding precursors [48]. Surprisingly, after androgen ablation, studies showed inconsistent outcomes regarding Treg levels and function. These contradictions might be explained by understanding that castration still leaves androgen and AR expression while ARKO completely abolish AR expression therefore some non-androgen-mediated pathways might still be functional through remaining AR in castrated WT mice to indorse Treg expansion. Additionally, it appears that variations in study approaches may also be to blame for these contradicting data. For example, castration intensified Treg cells expansion only when immune reactions against tumor antigens were stimulated [49].
The precise mechanisms by which Androgen/AR affects Treg formation and function have not yet been determined. However, Transforming growth factor-beta (TGF-ß) and interleukin-10 (IL10) are most investigated because of their capacity to block an antitumor immune response and their inhibitory effect on immune cells [50]. Androgen/androgen receptors (AR) contribute to the growth of T cells by upregulating the expression of TGF-ß and IL-10, which are necessary for the conversion of CD4+ T cells into Foxp3+ CD25high T cells (Tregs) [51, 52]. The findings of the current study are consistent with the finding that testosterone induces an increase in TGF-ß and IL-10 expression.
In conclusion, our results clearly point toward a route of intervention against such immunological disturbances. Nevertheless, further studies are needed to identify the molecular mechanism of how AR influences Treg development and differentiation. To sum up, the current study demonstrates for the first time in a knockout mice model that reduction of AR associated with a significant reduction in the numbers of Treg cells and their suppressive activities. Allogenic T cell growth was likewise unhindered by ARKO Treg cells. Therefore, this study likely points to evidence for androgens' functional significance in the development and activities of Treg cells. This role is not solely central for discerning the pathophysiology of male reproductive dysfunctions like autoimmune orchitis but also for autoimmune diseases at large. Additional research into the cellular and molecular mechanisms regulating the role of androgens in Treg proliferation and activity may uncover the fundamental processes underlying novel treatment approaches to autoimmune disorders.
Acknowledgement
The researchers would like to thank the Deanship of Scientific Research, Qassim University, for the support of this project.
Funding
The National Institute of Health (NIH Grant IR01DK073414-01.2007-2010) funded this research.
Author Contributions
Altuwaijri designed and conducted the experiments. Altuwaijri and Albarrak analyzed the data and drafted the manuscript.
The authors declare that there is no conflict of interest.
![]() References
References
- Walecki M, Eisel F, Klug J, Baal N, Paradowska-Dogan A, et al. Androgen receptor modulates Foxp3 expression in CD4+ CD25+ Foxp3+ regulatory T-cells. Molecular Biology of The Cell, (2015); 26(15): 2845-2857.
- DeBoer MD, Phillips BR, Mauger DT, Zein J, Erzurum SC, et al. Effects of endogenous sex hormones on lung function and symptom control in adolescents with asthma. BMC Pulmonary Medicine, (2018); 18 (58): 1-10.
- Zein JG, McManus JM, Sharifi N, Erzurum SC, Marozkina N, et al. Benefits of airway androgen receptor expression in human asthma. American Journal of Respiratory and Critical Care Medicine, (2021); 204(3): 285-293.
- Conforti F, Pala L, Bagnardi V, De Pas T, Martinetti M, et al. Cancer immunotherapy efficacy and patients' sex: a systematic review and meta-analysis. The Lancet Oncology, (2018); 19(6): 737-746.
- Abo-Aziza FAM, Zaki AA, Amer AS, Lotfy RA. Dihydrotestosterone and 17b-Estradiol Enhancement of In Vitro Osteogenic Differentiation of Castrated Male Rat Bone Marrow Mesenchymal Stem Cells (rBMMSCs). International Journal of Hematology-Oncology and Stem Cell Research, (2019); 13(4): 208-219.
- Dinesh R, Hahn B, Singh R; Sex hormones and gender influence the expression of Foxp3 and regulatory T cells in SLE patients. The Journal of Immunology, (2011); 186 (1_Supplement): 115-118.
- Kamal M, Gabr H, Anwar S, Bastawy S, Salah L. The relation of CD4+ CD25+ Foxp3+ regulatory T cells concentration with disease activity and damage index in systemic lupus erythematosus. Lupus, (2022); 31(4): 463-471.
- Fiyouzi T, Pelaez-Prestel HF, Reyes-Manzanas R, Lafuente EM, Reche PA. Enhancing regulatory T cells to treat inflammatory and autoimmune diseases. International Journal of Molecular Sciences, (2023); 24(9): 7797.
- Giefing-Kröll C, Berger P, Lepperdinger G, Grubeck-Loebenstein B. How sex and age affect immune responses, susceptibility to infections, and response to vaccination. Aging Cell, (2015); 14(3): 309–321.
- Polanczyk MJ, Carson BD, Subramanian S, Afentoulis M, Vandenbark AA, et al; Cutting edge: estrogen drives expansion of the CD4+CD25+ regulatory T cell compartment. Journal of Immunology, (2004); 173(40): 2227–2230.
- Fijak M, Schneider E, Klug J, Bhushan S, Hackstein H, et al; Testosterone replacement effectively inhibits the development of experimental autoimmune orchitis in rats: evidence for a direct role of testosterone on regulatory T cell expansion. Journal of Immunology, (2011); 186(9): 5162–5172.
- Marozkina N, Zein J, DeBoer MD, Logan L, Veri L, Ross K, et al; Dehydroepiandrosterone supplementation may benefit women with asthma who have low androgen levels: a pilot study. Pulmonary Therapy, (2019); 5: 213-20.
- Page ST, Plymate SR, Bremner WJ, Matsumoto AM, Hess DL, et al; Effect of medical castration on CD4+ CD25+ T cells, CD8+ T cell IFN-γ expression, and NK cells: a physiological role for testosterone and/or its metabolites. American Journal of Physiology-Endocrinology and Metabolism, (2006); 290(5): E856-E863.
- Braitch M, Harikrishnan S, Robins RA, Nichols C, Fahey AJ, et al. Glucocorticoids increase CD4+ CD25high cell percentage and Foxp3 expression in patients with multiple sclerosis. Acta Neurologica Scandinavica, (2009); 119(4): 239-245.
- Bohács A, Cseh Á, Stenczer B, Müller V, Gálffy G, et al; Effector and regulatory lymphocytes in asthmatic pregnant women. American Journal of Reproductive Immunology, (2010); 64(6): 393-401.
- Guan X, Polesso F, Wang C, Sehrawat A, Hawkins RM, et al. Androgen receptor activity in T cells limits checkpoint blockade efficacy. Natur, (2022); 606(7915): 791-796.
- Zein J, Gaston B, Bazeley P, DeBoer MD, Igo Jr RP, et al. HSD3B1 genotype identifies glucocorticoid responsiveness in severe asthma. Proceedings of the National Academy of Sciences, (2020); 117(4): 2187-2193.
- Gandhi VD, Cephus JY, Norlander AE, Chowdhury NU, Zhang J, et al. Androgen receptor signaling promotes Treg suppressive function during allergic airway inflammation. The Journal of Clinical Investigation, (2022); 132(4): 1-15.
- Alawad A, Altuwaijri S, Aljarbu A, Kryczek I, Niu Y, et al. Depletion of androgen receptor (AR) in mesenchymal stem cells (MSCs) inhibits induction of CD4+ CD25+ FOX3+ regulatory T (Treg) cells via androgen TGF-β interaction. Journal of Applied Biomedicine, (2015); 13(4): 263-271.
- Yeh S, Tsai MY, Xu Q, Mu XM, Lardy H, et al; Generation and characterization of androgen receptor knockout (ARKO) mice: an in vivo model for the study of androgen functions in selective tissues. Proceedings of the National Academy of Sciences, (2002); 99(21): 13498-13503.
- Altuwaijri S, Chuang KH, Lai KP, Lai JJ, Lin HY, et al. Susceptibility to autoimmunity and B cell resistance to apoptosis in mice lacking androgen receptor in B cells. Molecular Endocrinology, (2009); 23(4): 444-453.
- Duque A, Rakic P; Different effects of bromodeoxyuridine and [3H] thymidine incorporation into DNA on cell proliferation, position, and fate. Journal of Neuroscience, (2011); 31(42): 15205-15217.
- Gray DH, Fletcher AL, Hammett M, Seach N, Ueno T, et al. Unbiased analysis, enrichment and purification of thymic stromal cells. Journal of immunological methods, (2008); 329(1-2): 56-66.
- Kim R, Emi M, Tanabe K. Cancer immunosuppression and autoimmune disease: beyond immunosuppressive networks for tumour immunity. Immunology, (2006); 119(2): 254-264.
- Rowe JH, Ertelt JM, Xin L, Way SS. Pregnancy imprints regulatory memory that sustains anergy to fetal antigen. Nature, (2012); 490(7418): 102-106.
- Moulton VR. Sex hormones in acquired immunity and autoimmune disease. Frontiers in immunology, (2018); 9: 2279.
- Klein SL, Flanagan KL. Sex differences in immune responses. Nature Reviews Immunology, (2016); 16: 626-638.
- Shimba A, Ikuta K. Glucocorticoids regulate circadian rhythm of innate and adaptive immunity. Frontiers In Immunology, (2020); 11: 2143.
- Wilhelmson AS, Lantero Rodriguez M, Johansson I, Svedlund Eriksson E, Stubelius A, et al. Androgen receptors in epithelial cells regulate thymopoiesis and recent thymic emigrants in male mice. Frontiers In Immunology, (2020); 11: 1342.
- Ikuta K, Ejima A, Abe S, Shimba A. Control of immunity and allergy by steroid hormones. Allergology International, (2022); 71(4):432-436.
- Dikiy S, Rudensky AY. Principles of regulatory T cell function. Immunity, (2023); 56(2): 240-255.
- Eggenhuizen PJ, Ng BH, Ooi JD. Treg enhancing therapies to treat autoimmune diseases. International Journal of Molecular Sciences, (2020); 21(19): 7015.
- Tough DF, Lombardi G. Therapeutic opportunities for regulatory T-cell enhancing approaches. Clinical and Experimental Immunology, (2023); 211(2): 93-95.
- McCallion O, Bilici M, Hester J, Issa F. Regulatory T-cell therapy approaches. Clinical and Experimental Immunology, (2023); 211(2): 96-107.
- Wan YY, Flavell RA. Identifying Foxp3-expressing suppressor T cells with a bicistronic reporter. Proceedings of the National Academy of Sciences, (2005); 102(14): 5126-5131.
- Sakaguchi S. Naturally arising Foxp3-expressing CD25+ CD4+ regulatory T cells in immunological tolerance to self and non-self. Nature immunology, (2005); 6(4): 345-352.
- Liva SM, Voskuhl RR. Testosterone acts directly on CD4+ T lymphocytes to increase IL-10 production. The Journal of Immunology, (2001); 167(4): 2060-2067.
- Liu Z, Falo LD, You Z. Knockdown of HMGB1 in tumor cells attenuates their ability to induce regulatory T cells and uncovers naturally acquired CD8 T cell-dependent antitumor immunity. The Journal of Immunology, (2011); 187(1):118-125.
- Shevyrev D, Tereshchenko V. Treg heterogeneity, function, and homeostasis. Frontiers In Immunology, (2020); 10: 3100.
- Okeke EB, Uzonna JE. The pivotal role of regulatory T cells in the regulation of innate immune cells. Frontiers In Immunology, (2019); 10: 680.
- Lee W, Lee GR. Transcriptional regulation and development of regulatory T cells. Experimental & molecular medicine, (2018); 50(3): e456- e456.
- Schiavon V, Duchez S, Branchtein M, How-Kit A, Cassius C, et al. Microenvironment tailors nTreg structure and function. Proceedings of The National Academy of Sciences, (2019); 116(13): 6298-6307.
- Malviya V, Yshii L, Junius S, Garg AD, Humblet‐Baron S, Schlenner SM. Regulatory T‐cell stability and functional plasticity in health and disease. Immunology and Cell Biology, (2023); 101(2): 112-29.
- Amini L, Kaeda J, Fritsche E, Roemhild A, Kaiser D, Reinke P. Clinical adoptive regulatory T Cell therapy: State of the art, challenges, and prospective. Frontiers In Cell and Developmental Biology, (2023); 10: 1081644.
- Baron KJ, Turnquist HR. Clinical Manufacturing of Regulatory T Cell Products For Adoptive Cell Therapy and Strategies to Improve Therapeutic Efficacy. Organogenesis, (2023); 19(1): 2164159.
- Selck C, Dominguez-Villar M. Antigen-specific regulatory T cell therapy in autoimmune diseases and transplantation. Frontiers In Immunology, (2021); 12: 661875.
- Abramson J, Anderson G. Thymic epithelial cells. Annual Review of Immunology, (2017); 35: 85-118.
- Rodrigues PM, Ribeiro AR, Serafini N, Meireles C, Di Santo JP, Alves NL. Intrathymic deletion of IL-7 reveals a contribution of the bone marrow to thymic rebound induced by androgen blockade. The Journal of Immunology, (2018); 200(4):1389-98
- Tang S, Dubey P. Opposing effects of androgen ablation on immune function in prostate cancer. Oncoimmunology, (2012); 1(7): 1220-1.
- Samiea A, Yoon JS, Ong CJ, Zoubeidi A, Chamberlain TC, et al. Interleukin‐10 induces expression of neuroendocrine markers and PDL1 in prostate cancer cells. Prostate cancer, (2020); 2020(1): 5305306.
- Hsu P, Santner-Nanan B, Hu M, Skarratt K, Lee CH, et al. IL-10 potentiates differentiation of human induced regulatory T cells via STAT3 and Foxo1. The Journal of Immunology, (2015); 195(8): 3665-3674.
- Liu KS, Fan XQ, Zhang L, Wen QN, Feng JH, et al.; Effects of recombinant human interleukin‑10 on Treg cells, IL‑10 and TGF‑β in transplantation of rabbit skin. Molecular medicine reports, (2014); 9(2):639-644.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()