

Full Length Research Article
In Silico Approach for Identifying Potent Natural Peptide Deformylase Inhibitors to Combat Antibacterial Resistance
Amer Al Ali
Adv. life sci., vol. 12, no. 1, pp. 157-162, February 2025
*– Corresponding Author: Amer Al Ali (Email: ameralali@ub.edu.sa)
Authors' Affiliations
[Date Received: 19/06/2024; Date Revised: 30/10/2024; Date Published: 31/12/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Antibiotic resistance is a major worldwide health problem, caused by the emergence and spread of multidrug-resistant bacteria. Peptide deformylase (PDf) is crucial for many pathogenic microbes but is not required for cytoplasmic protein synthesis in eukaryotes, making it an attractive target for novel antibacterial agents.
Methods: This study computationally screened the ZINC database natural compounds against the PDf using the PyRx 0.8. The SwissADME and ADMET AI web tools were utilized to evaluate the physicochemical and ADMET characteristics of the five selected compounds.
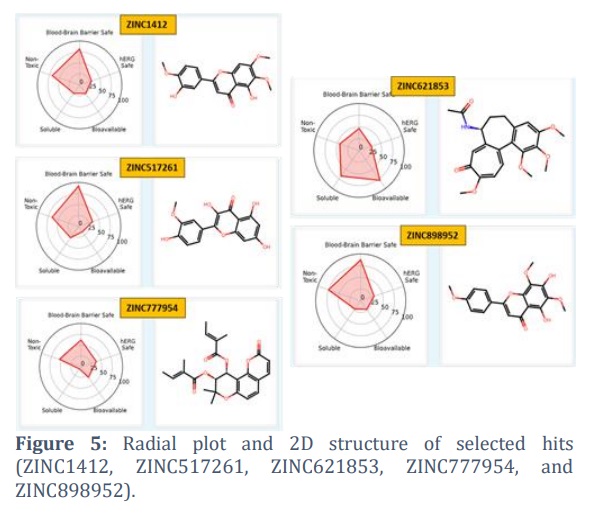
Results: The compounds ZINC1412, ZINC517261, ZINC621853, ZINC777954, and ZINC898952 were found to be higher-affinity binders than the positive control, interacting with critical PDf residues. Furthermore, the physicochemical and drug-like properties of these compounds demonstrated that they possess a range of safe and acceptable parameters. The ADMET properties of these compounds showed that they are within an acceptable range.
Conclusion: These compounds (ZINC1412, ZINC517261, ZINC621853, ZINC777954, and ZINC898952) can be used as PDf inhibitors against MDR bacteria after experimental validation.
Keywords: Antibiotic resistance, peptide deformylase, natural compound, drug-like
Introduction![]()
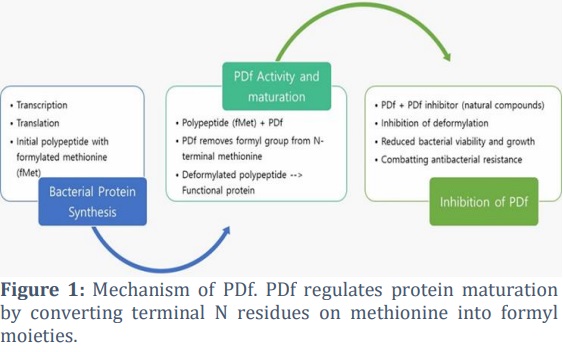
Antibiotic resistance, a critical global health challenge in the modern era [1,2], is primarily driven by the emergence, dissemination, and persistence of multidrug-resistant (MDR) bacteria, commonly known as ‘superbugs.’ These superbugs are responsible for infections that are resistant to conventional therapeutic interventions. The widespread and indiscriminate use of antibiotics in both human and animal health, combined with a lack of innovation in antibiotic research, as evidenced by a decline in the introduction of novel antibiotic classes, is a significant contributor to the development and spread of antimicrobial resistance [3]. It is imperative that we accelerate our efforts, not only to develop policies that curb the inappropriate and irrational use of antibiotics but also to focus on the development of novel chemical entities that can effectively combat bacterial infections [4].Peptide deformylase (PDf), a metalloenzyme that regulates protein maturation by converting terminal N residues on methionine into formyl moieties [5,6], holds immense promise as a target for the development of new antimicrobial agents (Fig. 1). Initially believed to be exclusive to bacteria and lacking targeted drugs, PDfs were seen as a beacon of hope for the development of new antibacterial agents [7-9]. Despite the identification and characterization of functional PDf homologs in eukaryotes, including a mitochondrial isoform in humans [10-14], which has posed challenges to the enzyme’s unambiguous designation as a relevant antibiotic target, enzymatic and structural studies have revealed significant differences in PDf ligand binding sites between prokaryotic and organellar cells [15-17], confirming the enzyme's status as a compelling and relevant therapeutic target.
Structure-based drug discovery has emerged as a critical methodology for facilitating rapid and cost-effective lead identification and optimization. The rational application of structure-based drug design proves to be far more effective than traditional drug discovery approaches. This efficacy is attributable to its focus on comprehending the molecular underpinnings of disease and leveraging insights derived from the three-dimensional structure of the biological target throughout the drug development process [18]. This study aimed to find natural PDf using the in-silico tool.
Methods![]()
The 3D structure of PDf in conjunction with Actinonin and zinc ion was obtained from the Protein Data Bank (PDB ID: 1SZZ). It is a homo 2-mer structure; the co-crystallized ligands, and water molecules were removed, and the cleaned protein structure was saved in ‘.pdb’ format.
Ligand Preparation
The natural compounds (N=200) utilized in the study were collected from the ZINC database. These compounds have been retrieved in .sdf format, energy minimized with a universal force field and converted to ‘.pdbqt’ format for further virtual screening (VS) analysis.
Virtual Screening
The AutoDock Vina platform, which is available in PyRx 0.8, was used to carry out the structure-based virtual screening of the prepared library of natural compounds [19]. The compounds and receptor enzyme were converted into .pdbqt format within the PyRx environment before a docking protocol was implemented. Actinonin was used as positive control during virtual screening, the lowest binding energy criterion was used to rank the best-docked conformations of compounds to the enzyme. Discovery studio visualizer was used to analyze and visualize the docking results and poses.
Physicochemical and ADMET Properties prediction
The SwissADME and ADMET AI web tools were utilized to evaluate the physicochemical and ADMET characteristics of the five selected compounds. The tools were initially evaluated, assessing pertinent variables such as molecular weight, logP, solubility, bioavailability, blood-brain barrier permeability, hERG inhibition risk, and overall toxicity
Results![]()
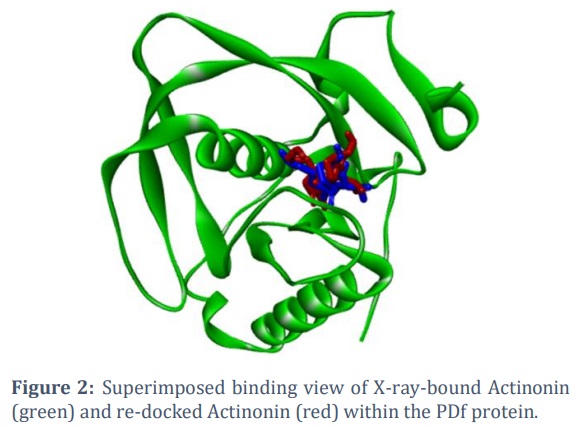
The PDf protein used in this screening had its crystal structure determined in combination with the inhibitor Actinonin (PDB ID: 1SZZ). To validate the docking process, Actinonin was withdrawn from the PDf protein complex and then re-docked to it. The binding posture of the re-docked Actinonin was computed and compared to the original co-crystal ligand orientation. The results showed that Actinonin binds in the same binding region of the PDf protein as seen in the co-crystal structure (Figure 2). This consistency verifies the docking protocol's ability to reliably anticipate ligand binding orientation within the PDf protein's binding pocket.
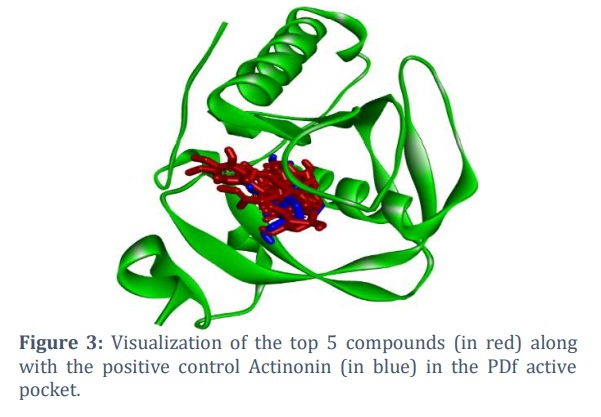
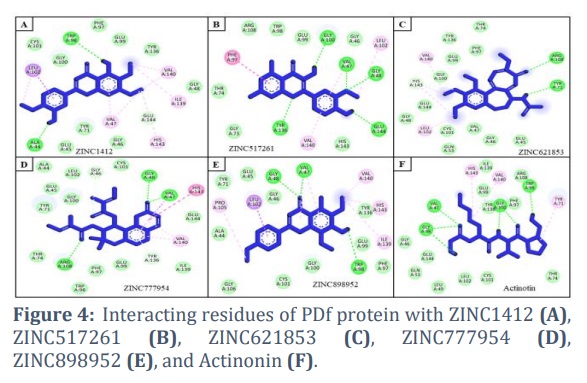
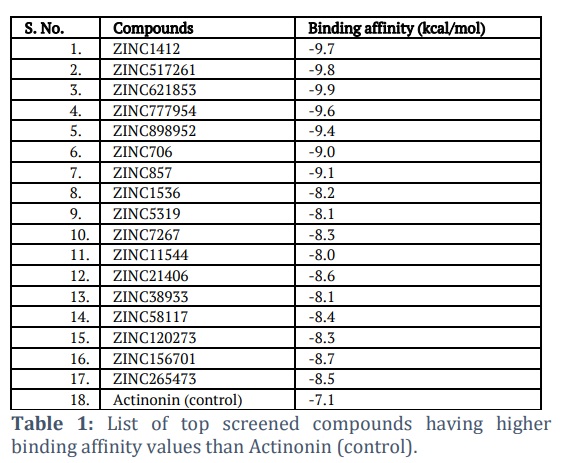
In this study, 200 natural compounds were computationally screened to identify novel natural inhibitors by targeting PDf active site residues. VS identified several potential compounds with higher binding affinities than the positive control, Actinonin (Table 1). Five compounds (ZINC1412, ZINC517261, ZINC621853, ZINC777954, and ZINC898952) were chosen for further investigation due to their stronger binding interactions with key PDf residues. This assessment was based on binding affinity values, as well as a thorough investigation and visualization of the interactions between the docked complexes (Figure 3).
ZINC1412 was found to interact with Leu102, Cys101, Gly100, Trp98, Phe97, Glu99, Tyr136, Val140, Gly48, Ile139, Glu144, His143, Gly46, Val47, Tyr71, Glu45, and Ala44 residues of PDf protein (Fig. 4A); while ZINC517261 interacted with Phe97, Arg108, Trp98, Glu99, Gly100, Gly46, Val47, Leu102, Gly48, Glu144, His143, Val140, Tyr136, Gly73, and Thr74 residues of PDf protein (Fig. 4B). ZINC621853 binds with Glu45, Gly46, Val47, Gln53, Cys101, Leu102, Gly48, Glu144, His143, Gly100, Val140, Glu99, Tyr136, Thr74, Arg108, and Tyr71 residues of PDf protein (Fig. 4C). ZINC777954 was found to bind with His143, Glu144, Val140, Ile139, Tyr136, Glu99, Phe97, Trp98, Arg108, Thr74, Tyr71, Gly100, Glu45, Ala44, Leu102, Gly46, Cys101, Gly48, and Val47 residues of PDf protein (Fig. 4D). ZINC898952 interacted with Pro105, Tyr71, Glu45, Leu102, Gly46, Gly48, Val47, Val140, His143, Tyr136, Ile139, Glu99, Phe97, Trp98, Gly100, Cys101, Gly106, and Ala44 residues of PDf protein (Fig. 4E). In addition, the control compound Actinonin was found to interact with Tyr71, Thr74, Cys101, Leu102, Leu49, Gln53, Glu144, Gly46, Gly48, Val47, His143, Ile139, Val140, Glu99, Tyr136, Gly100, Phe97, Trp98, and Arg108 residues of PDf protein (Fig. 4F).
The physicochemical, pharmacokinetic, and drug-likeness properties of the five chosen hits were evaluated using SwissADME. The analysis demonstrated that these compounds possess a range of safe and acceptable parameters. More precisely, the assessment of drug-likeness using Lipinski's, Ghose's, Veber's, Egan's, and Muegge's rules showed that none of the five hits violated any of these rules. These compounds can potentially serve as valuable lead compounds (Table 2).
The ADMET-AI online tool generated a radial plot, shown in Figure 5, that summarizes five key ADMET characteristics: blood-brain barrier permeability, risk of hERG inhibition, bioavailability, solubility, and toxicity. These characteristics are shown in relation to Drugbank's percentile rankings. Each compound chosen falls within the acceptable range for these five factors. The two-dimensional structures of the chosen compounds are also shown in Figure 5.
Figures & Tables

Antibiotic resistance is a serious health concern, emphasizing the urgent need for new antibacterial agents with novel mechanisms of action [6]. Current antibiotics only target a small number of molecules. The initiation process for bacterial and mammalian cytosolic protein synthesis differs significantly: bacteria use N-formylmethionine [9,20] [9086272; 8112305], whereas mammalian cells use methionine [21]. Bacterial N-formylmethionine is produced by formylmethionine tRNA transferase and then removed by PDf and methionine aminopeptidase, resulting in mature proteins [22]. This formylation-deformylation cycle is critical for bacterial growth but not in mammalian cells, making PDf a promising target for selective antibiotic development [8]. In this study, a natural compound library was screened against the PDf protein, and compounds ZINC1412, ZINC517261, ZINC621853, ZINC777954, and ZINC898952 were identified as high-affinity binders that interact with critical PDf residues.
Hydrogen bonds are critical for stabilizing ligand-protein complexes [23,24]. The identified compounds, ZINC1412, ZINC517261, ZINC621853, ZINC777954, and ZINC898952, form hydrogen bonds with multiple PDf amino acid residues. ZINC1412 makes hydrogen bonds with Trp98 and Ala44, whereas ZINC517261 H-bonded with Gly100, Val47, Gly48, Glu144, and Tyr136 residues of PDf. ZINC621853 forms hydrogen bonds with Arg108 and Tyr71, while ZINC777954 H-bonded with Gly48, Val47, and Arg108 residues of PDf. Furthermore, ZINC898952 forms hydrogen bonds with Gly48, Val47, Trp98, and Arg108 residues of PDf.
Actinonin, a naturally occurring antibacterial agent derived from the Streptomyces species, is the first known inhibitor of PDf with a metal ion-binding hydroxamate moiety [25,26]. In this study, key PDf protein residues involved in Actinonin binding include Tyr71, Thr74, Cys101, Leu102, Leu49, Gln53, Glu144, Gly46, Gly48, Val47, His143, Ile139, Val140, Glu99, Tyr136, Gly100, Phe97, Trp98, and Arg108. Interestingly, the compounds ZINC1412, ZINC517261, ZINC621853, ZINC777954, and ZINC898952 were found to interact with most of these residues, implying that they likely bind to the same PDf pocket as Actinonin.
High negative binding affinity values indicate a ligand's strong interaction with its target protein [27-30]. Notably, the identified compounds ZINC1412, ZINC517261, ZINC621853, ZINC777954, and ZINC898952 have higher negative binding affinity values than the control compound Actinonin, indicating that they have a stronger binding affinity for the PDf protein.
Natural products are important sources of novel medications and templates for synthetic drug development, including anticancer therapies and antibiotics. Many of these medications are created by microorganisms or interact with hosts [31]. Exploring natural compounds with antibacterial characteristics is critical given the growth in plasmid-mediated antibiotic resistance and the scarcity of viable treatments for certain respiratory and neurological illnesses. The WHO recommends using medicinal herbs as an alternate treatment when conventional options are unavailable. The research focuses on bioactive substances, their chemical composition, and pharmacological potential to develop less harmful alternatives to existing medications. Natural compounds are increasingly being employed to treat microbial diseases, inflammation, and cancer due to their accessibility and therapeutic potential [32]. The identified hit compounds in this study demonstrated strong affinity with the PDf protein, indicating efficacy against antimicrobial resistance.
PDf, a metallohydrolase required for bacterial growth, is a promising target for new antibiotic discovery. This in silico study screened the natural compounds against the PDf protein. ZINC1412, ZINC517261, ZINC621853, ZINC777954, and ZINC898952 were found to bind strongly to PDf and interact with critical PDf protein residues. These compounds can be employed as PDf inhibitors to combat MDR bacteria.
Acknowledgements
The author is thankful to the Deanship of Graduate Studies and Scientific Research at University of Bisha for supporting this work through the Fast-Track Research Support Program.
Author Contributions
AAA: wrote, analyze, and revised the manuscript.
The authors declare that there is no conflict of interest regarding the publication of this paper.
![]() References
References
- Brockhurst MA, Harrison F, Veening JW, Harrison E, Blackwell G, et al. Assessing evolutionary risks of resistance for new antimicrobial therapies. Nature Ecology & Evolution, (2019); 3(4): 515-517.
- Cattoir V, Felden B. Future Antibacterial Strategies: From Basic Concepts to Clinical Challenges. The Journal of Infectious Diseases, (2019); 220(3): 350-360.
- Fair RJ, Tor Y. Antibiotics and bacterial resistance in the 21st century. Perspectives in Medicinal Chemistry, (2014); 6: 25-64.
- Simpkin VL, Renwick MJ, Kelly R, Mossialos E. Incentivising innovation in antibiotic drug discovery and development: progress, challenges and next steps. The Journal of antibiotics (Tokyo), (2017); 70(12): 1087-1096.
- Fieulaine S, Alves de Sousa R, Maigre L, Hamiche K, Alimi M, et al. A unique peptide deformylase platform to rationally design and challenge novel active compounds. Scientific Reports, (2016); 6: 35429.
- Apfel CM, Locher H, Evers S, Takacs B, Hubschwerlen C, et al. Peptide deformylase as an antibacterial drug target: target validation and resistance development. Antimicrobial Agents and Chemotherapy, (2001); 45(4): 1058-1064.
- Giglione C, Pierre M, Meinnel T. Peptide deformylase as a target for new generation, broad spectrum antimicrobial agents. Molecular Microbiology, (2000); 36(6): 1197-1205.
- Yuan Z, Trias J, White RJ. Deformylase as a novel antibacterial target. Drug Discovery Today, (2001); 6(18): 954-961.
- Mazel D, Pochet S, Marliere P. Genetic characterization of polypeptide deformylase, a distinctive enzyme of eubacterial translation. The EMBO Journal, (1994); 13(4): 914-923.
- Giglione C, Serero A, Pierre M, Boisson B, Meinnel T. Identification of eukaryotic peptide deformylases reveals universality of N-terminal protein processing mechanisms. The EMBO Journal, (2000); 19(21): 5916-5929.
- Serero A, Giglione C, Meinnel T. Distinctive features of the two classes of eukaryotic peptide deformylases. Journal of Molecular Biology, (2001); 314(4): 695-708.
- Nguyen KT, Hu X, Colton C, Chakrabarti R, Zhu MX, Pei D. Characterization of a human peptide deformylase: implications for antibacterial drug design. Biochemistry, (2003); 42(33): 9952-9958.
- Lee MD, Antczak C, Li Y, Sirotnak FM, Bornmann WG, Scheinberg DA. A new human peptide deformylase inhibitable by actinonin. Biochemical and Biophysical Research Communications, (2003); 312(2): 309-315.
- Escobar-Alvarez S, Gardner J, Sheth A, Manfredi G, Yang G, et al. Inhibition of human peptide deformylase disrupts mitochondrial function. Molecular and Cellular Biology, (2010); 30(21): 5099-5109.
- Escobar-Alvarez S, Goldgur Y, Yang G, Ouerfelli O, Li Y, Scheinberg DA. Structure and activity of human mitochondrial peptide deformylase, a novel cancer target. Journal of Molecular Biology, (2009); 387(5): 1211-1228.
- Fieulaine S, Juillan-Binard C, Serero A, Dardel F, Giglione C, et al. The crystal structure of mitochondrial (Type 1A) peptide deformylase provides clear guidelines for the design of inhibitors specific for the bacterial forms. Journal of Biological Chemistry, (2005); 280(51): 42315-42324.
- Fieulaine S, Desmadril M, Meinnel T, Giglione C. Understanding the highly efficient catalysis of prokaryotic peptide deformylases by shedding light on the determinants specifying the low activity of the human counterpart. Acta crystallographica Section D biological crystallography, (2014); 70(Pt 2): 242-252.
- Lionta E, Spyrou G, Vassilatis DK, Cournia Z. Structure-based virtual screening for drug discovery: principles, applications and recent advances. Current Topics in Medicinal Chemistry, (2014); 14(16): 1923-1938.
- Dallakyan S, Olson AJ. Small-molecule library screening by docking with PyRx. Methods in Molecular Biology, (2015); 1263: 243-250.
- Mazel D, Coic E, Blanchard S, Saurin W, Marliere P. A survey of polypeptide deformylase function throughout the eubacterial lineage. Journal of Molecular Biology, (1997); 266(5): 939-949.
- Adams JM, Capecchi MR. N-formylmethionyl-sRNA as the initiator of protein synthesis. Proceedings of the National Academy of Sciences USA, (1966); 55(1): 147-155.
- Meinnel T, Blanquet S. Evidence that peptide deformylase and methionyl-tRNA(fMet) formyltransferase are encoded within the same operon in Escherichia coli. Journal of Bacteriology, (1993); 175(23): 7737-7740.
- Helmi N. Identification of therapeutic phytochemicals targeting B-cell lymphoma 2 (BCL2) as anti-acute myeloid leukemia agents: An in-silico approach. Advancements in Life Sciences, (2024); 10(4): 670-674.
- Sayed Murad HA, M Rafeeq M, Alqahtani SM, S Rajab B, Alghamdi S, J Almehmadi S, Alam Q. Molecular docking analysis of AGTR1 antagonists. Bioinformation, (2023); 19(3): 284-289.
- Chen DZ, Patel DV, Hackbarth CJ, Wang W, Dreyer G, et al. Actinonin, a naturally occurring antibacterial agent, is a potent deformylase inhibitor. Biochemistry, (2000); 39(6): 1256-1262.
- Gordon JJ, Kelly BK, Miller GA. Actinonin: an antibiotic substance produced by an actinomycete. Nature, (1962); 195: 701-702.
- Alkathiri AS. Structure-Based Virtual Screening of Natural Compounds for Inhibition of Protein Tyrosine Phosphatase 1B: A Promising Therapeutic Approach in Diabetes Management. Advancements in Life Sciences, (2024); 11(1): 200-205.
- Sait KHW, Alam Q, Anfinan N, Al-Ghamdi O, Malik A, et al. Structure-based virtual screening and molecular docking for the identification of potential novel EGFRkinase inhibitors against ovarian cancer. Bioinformation, (2019); 15(4): 287.
- Sait KHW, Mashraqi M, Khogeer AA, Alzahrani O, Anfinan NM, et al. Molecular docking analysis of HER-2 inhibitor from the ZINC database as anticancer agents. Bioinformation, (2020); 16(11): 882.
- Hakeem IJ, Alsharif FH, Aljadani M, Alabbas IF, Faqihi MS, et al. Molecular docking analysis of KRAS inhibitors for cancer management. Bioinformation, (2023); 19(4): 411.
- Newman DJ, Cragg GM. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. Journal of Natural Products, (2020); 83(3): 770-803.
- de Moraes Mello Boccolini P, Siqueira Boccolini C. Prevalence of complementary and alternative medicine (CAM) use in Brazil. BMC Complementary Medicine and Therapies, (2020); 20(1): 51.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()