

Full Length Research Article
Characterization of Lacticaseibacillus rhamnosus of Honey Origin from Saudi Arabia: Potential Probiotic Strains with Antibacterial Effects
Muhammad Barkaat Hussain1, Saleh M. Al-Maaqar2, Sayed Sartaj Sohrab3, Yasser Mahmoud Kamel Mohamed Aly1, Asif Ahmad M. Jiman-Fatani4, Muhammad Yasir3, Muhammad Absar5*
Adv. life sci., vol. 12, no. 1, pp. 141-149, February 2025
*– Corresponding Author: Muhammad Absar (Email: mashwani82@gmail.com)
Authors' Affiliations
2. Department of Biological Sciences, Faculty of Science, King Abdulaziz University, P.O. Box 80203, Jeddah – 21589 – Saudi Arabia
3. Special Infectious Agents Unit, King Fahd Medical Research Center, King Abdulaziz University, Jeddah – 21589 – Saudi Arabia
4. Department of Medical Microbiology and Parasitology, Faculty of Medicine, King Abdulaziz University (KAU), P.O. Box 80203, Jeddah – 21589 – Saudi Arabia
5. Department of Pathology and Laboratory Medicine, King Abdulaziz Hospital Ministry of National Guard Health Affairs, AlAhsa – Saudi Arabia
[Date Received: 11/10/2024; Date Revised: 17/11/2024; Date Published: 31/12/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: We recently isolated five strains of lactic acid bacteria (LAB) from honey and identified them via 16S ribosomal RNA gene sequencing. The sequences were deposited in the NCBI GenBank database and given the corresponding accession numbers: OL539410 to OL539414. Since LAB are well known for their probiotic properties, in this study, we characterized the probiotic potential and safety of these isolates.
Methods: Five LAB strains, including two strains of Lacticaseibacillus rhamnosus and three strains of Enterococcus faecium isolated from honey samples, were examined for their probiotic potential through acid and bile salt resistance assays, bacterial cell surface hydrophobicity, autoaggregation, and coaggregation assays. In addition, cell free culture supernatant antibacterial activity of mentioned LAB strains was tested against E. coli (ATCC 25922), P. aeruginosa (ATCC 27853), K. pneumoniae (ATCC 13883), S. aureus (ATCC 25923), S. epidermidis (ATCC 12228), and S. pneumoniae (ATCC 6303) Further tests, such as those measuring hemolytic activity and antibiotic susceptibility, were also performed to evaluate their safety profile.
Results: All the LAB strains tolerated and survived the simulated gastrointestinal conditions: pepsin at acidic pH (3.0) and bile salt concentration (0.3%) at alkaline pH (8.0), with different levels of viability. According to antibacterial assays, L. rhamnosus (P0-2 L) and E. faecium (P0-4 L) exhibited the strongest inhibitory activities. Also, L. rhamnosus (P0-2 L) demonstrated high adhesive and aggregative properties. All strains tested negative for hemolytic activity, indicating safety for human consumption.
Conclusion: This study revealed that L. rhamnosus derived from Sidr honey may be used as a potential probiotic. This study also showed that honey from different regions can contain probiotics.
Keywords: Lacticaseibacillus rhamnosus, Sidr honey, lactic acid bacteria, probiotics
Introduction![]()
Probiotic-containing food is in great demand worldwide because of the ongoing and active creation of research evidence indicating potential health benefits for its users. There are many different products that include probiotics, such as fermented dairy products, functional foods, and dietary supplements, which have been shown to have health benefits [1]. Natural food products and supplements have been most extensively accepted as vehicles for probiotic delivery, improving overall community health [2]. However, it is still unclear whether probiotics from supplements can withstand bile salts and acids, which could reduce their ability to offer health advantages [3]. Owing to the buffering effects of food on probiotics as they pass through the digestive tract, the availability of essential nutrients that maintain probiotic bacterial activity and efficacy, the synergistic effects of dietary components that promote probiotic growth, and consumer preferences for food containing probiotics over capsules, tablets, and other therapeutic formulations, foods may therefore be preferred over probiotic supplements [4]. An organism must meet certain criteria to qualify as a probiotic: it must be essentially nonpathogenic, tolerant of acidic pH and high quantities of bile salts, able to decrease pathogens attachment to mucosal surfaces and the ability to adhere to epithelial cells, and exhibit antibacterial activity against potential pathogens [5]. Furthermore, these organisms should not transfer antibiotic resistance genes through horizontal genetic mechanisms to potential pathogens [6].
Lactobacilli, bifidobacteria, and enterococci are the three types of probiotic bacteria that are widely used. According to research studies, probiotics can be beneficial for the human body in numerous ways [7]. The results from such studies include successful reports of probiotics adjuvant treatment for infectious diarrhea, diarrhea induced by chemotherapy and antibiotics, irritable bowel syndrome, Crohn’s disease, Clostridioides difficile colitis, and ulcerative colitis[8]. Furthermore, probiotics have a profound effect on the stimulation of the immune system, anti-cancer properties, antimicrobial properties, lowering cholesterol levels, anti-high blood pressure effects, and lowering adverse symptoms associated with lactose intolerance[9]. The latest research studies highlight the microbes origin probiotic substances and provide health benefits against different clinical conditions, like diabetes mellitus, mental and cardiac diseases [10].
Although dairy products have long been used as carriers for probiotics, they have several adverse effects, including lactose intolerance and milk protein allergies. Consequently, there is a growing body of research investigating nondairy foods such as vegetables, fruits, cereals, and honey as potential probiotic carriers [11].
The compounds present in honey defend against oxidative stress, exert prebiotic effects, and lower redox potential, all of which encourage the growth of probiotics [12]. According to recent research, honey contains a variety of LAB with probiotic properties [13].
The isolation of probiotic bacteria from sources other than dairy can increase the possibilities of obtaining probiotic bacterial strains with substantial functional properties. In the current study, we have isolated five strains of LAB from Somra, Talah, and Sidr honey, including two strains of L. rhamnosus and three E. faecium strains. These strains were identified by traditional phenotypic methods and also characterization by 16S rRNA gene sequence analysis[14]. To the best of our knowledge, so far the probiotic properties of lactic acid bacteria isolated from Saudi honey have never been conducted. The traits of probiotics differ by the type of strains, it is highly recommended to do further investigations to enhance our understanding of the probiotic capabilities of novel LAB strains. The current research study was based on the guidelines outlined by the Food and Agriculture Organization and World Health Organization, with the prime objective of evaluating the safety and probiotic properties of honey-derived LAB strains.
Methods![]()
Bacterial Strains
A total of five LAB strains were used in this study to evaluate their probiotic potential, comprising two strains of L. rhamnosus and three strains of E. faecium. The LAB strains were isolated from the following honey types: Somra, Talah, and Sidr, which were produced by Apis mellifera jementica (a subspecies of the western honey bee), from the Aseer and Al-Baha regions of Saudi Arabia. The 16S ribosomal RNA gene sequencing was performed to characterize these strains and the sequences were submitted to the NCBI GenBank database and assigned the corresponding accession numbers: OL539410 to OL539414 [14]. Furthermore, a reference probiotic commercial strain of L. rhamnosus (Protexin), was obtained from Nahdi Pharmacy in Jeddah. Other quality control strains tested as reference strains, included, S. aureus (ATCC 25923), E. coli (ATCC 25922), S. pneumoniae (ATCC 6303), S. epidermidis (ATCC 12228), P. aeruginosa (ATCC 27853), and K. pneumoniae (ATCC 13883), were obtained from the Biology Department at King Abdulaziz University (KAU) in Jeddah, Saudi Arabia, and were utilized for evaluating the antimicrobial and coaggregation assay of the LAB strains.
Acid and Bile Salt Resistance Assays
The tolerance of each LAB isolate to acid (pH 3.0) and bile (0.3%) was assessed using Al-Masaudi’s method [15]. Simulated gastric fluid was made from buffered peptone water. To achieve a pH of 3.0, one molar hydrochloric acid was used. Sterilized pepsin (Sigma, Spain) was added to buffered peptone water to 1000 U/ml. Bile salts (Biolife, Italy) at 0.3% (w/v) were added to create simulated intestinal fluid (SIF), and sodium hydroxide was used to adjust the buffered peptone water pH to 7. LAB strains were inoculated in MRS broth medium (Hi Media) at 37°C for 24 h and then centrifuged (3500×g, 6 min). Phosphate-buffered saline was used to wash harvested cells before they were resuspended in MRS broth (pH 7.0), which served as a control, and MRS broth (pH 3) containing pepsin (3 g/L) (Sigma‒Aldrich, USA), which was adjusted with 0.1 M HCl and incubated at 37°C. For enumeration, aliquots (100 μL) were obtained after 0, 1, and 3 h, plated on MRS agar, and then incubated for 3 days at 37°C. After a 3-h incubation period in SGF, the microorganisms were separated via centrifugation (3500 × g, 6 min), transferred to SIF, and incubated for 3 h at 37°C. After 1 and 3 h, samples (100 μL) were taken for plate counting. The experiment was repeated two times on the same day.
Antibacterial Activity Assay
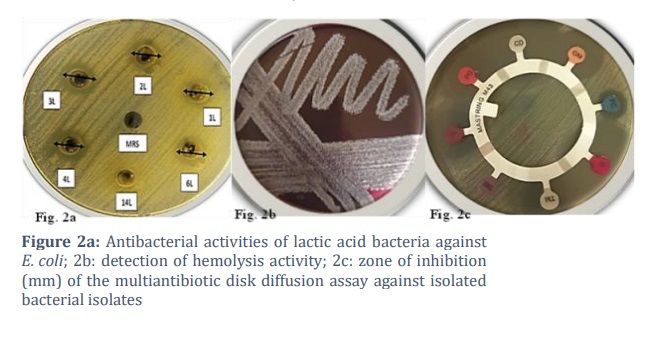
The antibacterial activity of cell-free culture supernatants (CFCSs) of LAB against selected ATCC strains, including E. coli (ATCC 25922), P. aeruginosa (ATCC 27853), K. pneumoniae (ATCC 13883), S. aureus (ATCC 25923), S. epidermidis (ATCC 12228), and S. pneumoniae (ATCC 6303), was assessed by an agar well diffusion assay based on the procedure described by Hussain et al. (2015)[16] with slight modifications. Briefly, the CFCS of LAB was obtained via centrifugation (3500 × g for 5 min) and filtration of the overnight culture of each isolate of LAB inoculated in MRS broth under anaerobic conditions at 37°C. Overnight cultures of ATCC reference strains were used to inoculate (107 colony-forming units) Muller‒Hinton agar plates (20 ml/plate). Using a sterile agar drill, 7-mm wells were punched in the inoculated agar plates. Each designated well was then filled with one hundred twenty microlitres of CFCS from each isolate of LAB. The same volume of MRS broth was used as a negative control. For 24 h, the plates were incubated at 37°C. On the same day, experiments were replicated, and the mean ± standard deviation (SD) was calculated to determine the inhibitory zone. L. rhamnosus, a probiotic strain available commercially, served as the control.
Hydrophobicity Assay
The basic idea of this method is to gauge the affinity of the suspension for the hydrocarbon solvent xylene. The LAB strains were cultured for eighteen h at 37°C in MRS broth and then centrifuged (1500 × g for 15 min). After the medium was removed, the cells were washed and resuspended in phosphate-buffered saline (PBS) at pH 7.4. The absorbance was set at approximately 0.25 ±0.05 (A0) to maintain a uniform count of bacteria (107-108 CFU/mL). An equal volume of xylene (Brixworth Northants, UK) was then added. After a 10-minute preincubation at 37°C, the cell suspensions were vortexed for 2 minutes and incubated at 37°C for 1 hour to separate the aqueous and organic phases. The hydrophobicity assay was conducted to determine the microbes' ability to adhere to hydrocarbons, which is a marker for gut epithelial cell adherence. The procedure by which LAB cell surfaces adhere to hydrocarbons is called microbial adhesion to hydrocarbons (MATH) [17].
This method measures the suspension's xylene affinity. The LAB strains were incubated in MRS broth at 37°C for 18 hours before centrifuging at 1500 g for 15 minutes. The media was discarded, and cells were washed and resuspended in PBS set at pH 7.4. The bacterial concentration was adjusted to 107-108 CFU/mL by measuring absorbance at 0.25 ± 0.05 (A0). Next, we added an equal volume of xylene (Brixworth, Northants, UK). After incubating the mixture for 10 minutes at 37°C, vortexed for 2 minutes and then incubated the suspension for 1 hour to separate the aqueous and organic phases. After incubation, the aqueous phase was carefully removed, and its absorbance was measured at 600 nm (A1). The following formula was used to calculate the percentage of bacteria adhering to the solvent:
Bacterial cell surface hydrophobicity % = 1 – (A1/A0) × 100
A1 = absorbance at 1 h (600 nm),
A0 = absorbance at 0 h (600 nm).
Autoaggregation assay and coaggregation assay
A spectrophotometry approach was used to assess the aggregation activity of the isolates. For eighteen h, the isolates were cultured in MRS media at 37°C. Centrifugation was used to harvest the cultures for 15 minutes at 5,000 × g to obtain the turbidity of the bacterial suspensions, which were adjusted to 0.5 (approximately 108 CFU/mL), washed 3 times with phosphate-buffered saline (PBS), and then resuspended in the same buffer following overnight incubation. After that, for four hours without stirring, the mixture was vortexed and incubated at 37°C. The percentage of autoaggregation was calculated as % = 1 – (ODt/OD0) × 100, where ODt and OD0 represent the absorbances at 4 h and 0 h (620 nm), respectively. In the same manner, as in the autoaggregation assay, bacterial suspensions for coaggregation were made. Equal amounts of S. aureus (ATCC 25923), E. coli (ATCC 25922), and LAB isolate bacterial suspensions were mixed and incubated at 37°C for 24 h without agitation for coaggregation analysis. Coaggregation was determined using the formula coaggregation % = [(Apath + ALAB)/2 – (Amix)/(Apath + ALAB)/2] × 100, where Apath and ALAB indicate the OD of tubes containing pathogens and LAB, respectively, and Amix is the OD of tubes containing the mixture, calculated at 620 nm.
Hemolytic activity analysis
To assess hemolysis, the LAB isolates were grown in MRS media for 24 h at 37°C. The isolates were then streaked on a blood agar (HiMedia) plate containing 5% (w/v) sheep blood for 48 h. If a halo formed around the colonies or a change in the color of the media was observed, the result was considered positive.
Antibiotic Susceptibility Assay
The antimicrobial susceptibility of five isolated LAB strains and one probiotic strain sold commercially, L. rhamnosus (Con-6 L), was assessed via the conventional disc diffusion method [18]. To prepare the overnight culture in the appropriate media, a single colony of each isolate was picked. After being diluted to approximately 105 CFU/mL, 200 μL of the inoculum was equally spread on MRS agar plates and allowed to dry for 15 minutes at room temperature. Antibiotic-containing discs (MAST Diagnostics, Merseyide, UK) were applied to the plate using sterile forceps. Following a 16-h incubation period at 37°C, the sizes of the inhibitory zones were calculated. The isolates were grouped as resistant (≤15 mm), intermediate (16–20 mm), or sensitive (≥21 mm) as described previously in the literature[18]. Antibiotic susceptibility tests were carried out with 16 antibiotics (Table 4).
Results![]()
Acid and Bile Salt Resistance Assays
The tolerances of the LAB honey isolates and the commercially available L. rhamnosus strain (Con-6 L) to simulated gastric and intestinal fluids were determined at different time intervals (after 1 h and 3 h), as shown in Table 1 and Fig. 1. All the LAB strains survived and maintained a viable count after 1 and 3 h at pH 3 in the gastric fluid. Similarly, they also tolerated and survived in intestinal fluid containing 0.3% bile at pH 8, maintaining viability for 3 h. As benchmarks for probiotic culture acid and bile tolerance, survival at pH 3 for two h and growth in a medium supplemented with 0.3% bile acid were recognized. Compared with the commercially available L. rhamnosus strain, there was no significant difference in the viability of all the tested strains; rather, E. faecium (P0-4 L) had a greater survival rate (42.16%) in the gastric fluid after 3 h than the L. rhamnosus strain (Con-6 L) (29.00%). Similarly, E. faecium (P01-1 L) also had a much greater survival rate (50.72%) in the intestinal fluid after 3 h than did the control strain (30.52%). E. faecium (P01-1 L) had the lowest survival rate (23.41%) in the simulated gastric fluid after 3 h, and L. rhamnosus (P14-5 L) had the lowest survival rate (23%) in the simulated intestinal fluid after 3 h.
Antimicrobial Activity
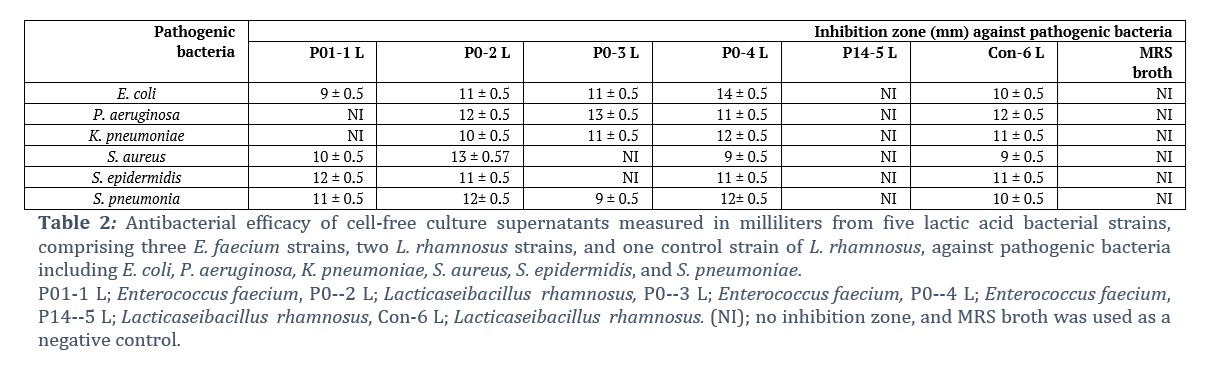
The antibacterial activity of cell-free culture supernatants (CFCSs) obtained from LAB strains against selected human pathogens is summarized in Table 2. The antagonistic activity of these strains was compared to that of L. rhamnosus (Con-6 L), a commercial probiotic strain. With varying degrees of antibacterial activity, the majority of the CFCSs of the tested LAB were able to inhibit the growth of selected pathogens. The CFCSs L. rhamnosus (P0-2 L) and E. faecium (P0-4 L) inhibited all the tested pathogenic bacterial isolates, similar to commercially available probiotic strains (Con-6 L) (Table 2). However, the CFCSs of Lacticaseibacillus rhamnosus (P14-5 L) did not show antagonistic activity against any of the tested pathogenic isolates. E. faecium (P01-1 L and P0-3 L) CFCSs showed variable or limited antibacterial activity against the tested pathogens; P. aeruginosa and K. pneumoniae were not inhibited by the CFCSs of E. faecium (P01-1 L), whereas S. aureus and S. epidermidis were resistant to the CFCSs of E. faecium (P0–3 L). The growth of the other tested pathogens was inhibited by E. faecium (P01-1 L and P0-3 L). This means that antibacterial activity does not exist in all lactic acid bacteria; rather, it is strain-specific. The CFCSs of E. faecium (P0-4 L) showed maximum inhibition (14 ± 0.5 mm) against E. coli, which was even greater than that of the control probiotic strain.
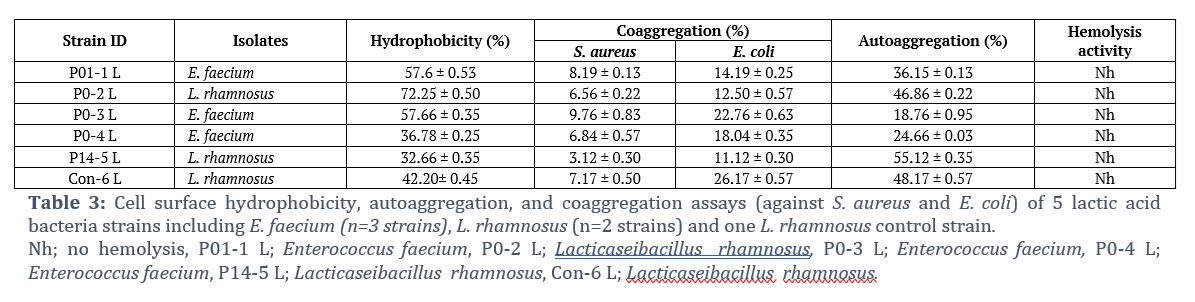
Cell Adhesion Activity of Isolates
The ability of LAB to adhere to intestinal epithelial cells is crucial for maintaining the presence of bacteria in the human gastrointestinal system since it can promote competition with other pathogenic microbes. The intestinal adhesion parameters are used in the selection of probiotic bacteria. Table 3 shows that the hydrophobicity values of the tested strains are variable in this study, ranging from 32.66 ± 0.35% to 72.25 ± 0.50%. A difference in hydrophobicity was observed among the tested isolates, even between the same species. Compared with the other isolates, L. rhamnosus (P0-2 L) was more hydrophobic (72.25 ± 0.50%) and even more hydrophobic than the control strain (42.20 ± 0.45%). In terms of adhesiveness, this isolate might be considered a possible probiotic culture. Variations in cell surface hydrophobicity are due to differences in the level of protein expression on the cell surface and in environmental conditions. The study findings demonstrated that every strain had a different autoaggregation percentage, ranging from 8.17 ± 0.57% to 55.12 ± 0.35% after 4 h. L. rhamnosus (P14-5 L and P0-2 L) presented the highest autoaggregation percentages of 55.12 ± 0.35 and 46.86 ± 0.22%, respectively, after 4 h. Coaggregation analysis was performed with S. aureus and E. coli (Table 3). The ability of all the tested probiotic strains to aggregate with pathogenic bacteria was demonstrated; however, the rates of coaggregation varied among the isolates. The bacterial isolate E. faecium (P0-3 L) had the greatest coaggregation percentage (22.76 ± 0.63%), whereas L. rhamnosus (P14-5 L) had the lowest coaggregation percentage with E. coli. Overall, all the tested isolates exhibited greater percentages of coaggregation with E. coli than with S. aureus.
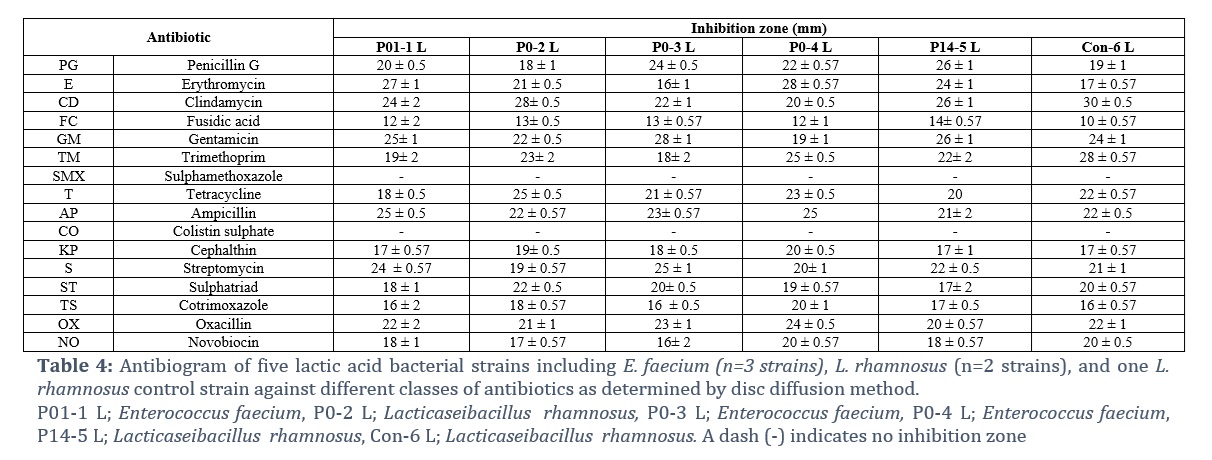
Antimicrobial Susceptibility Pattern
The isolates were divided into three groups: resistant (≤15 mm), intermediate (16–20 mm), and sensitive (≥21 mm)[18]. The examined LAB strains demonstrated varying levels of antibiotic susceptibility. The isolates were susceptible to 13 of the 16 tested antibiotics, and resistance to sulfamethoxazole, colistin sulfate, and fusidic acid was detected in 100% of the isolates (Table 4)
Figures & Tables
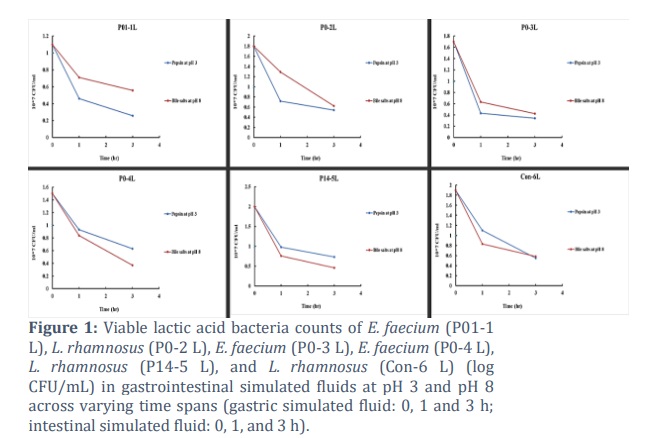
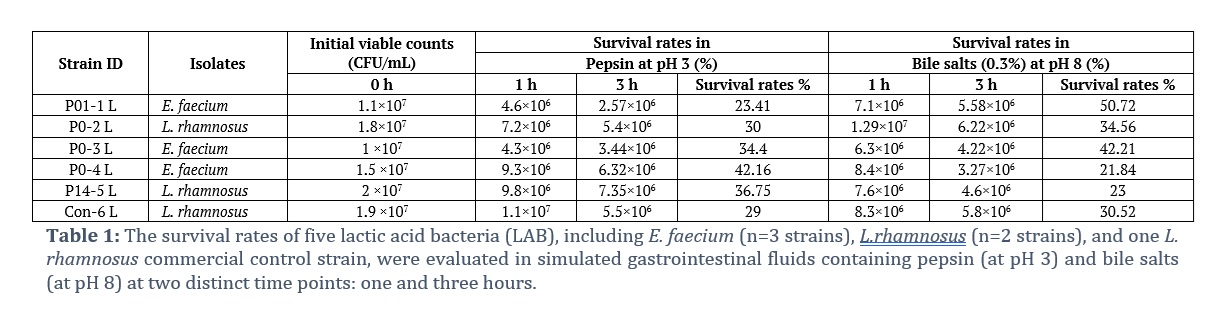
Honey has great nutritional and therapeutic value and is widely consumed worldwide. In both human and veterinary medicine, honey has been revived as a treatment for mild to moderate burns, skin ulcers, gastrointestinal disorders, allergic coughs, and infected wounds [19]. Owing to the presence of prebiotics, zinc, phenolic compounds, reducing sugars, and organic acids, its matrix offers the perfect environment for probiotic bacteria. However, honey, being a natural compound, exhibits variability in content and composition based on its botanical origin, geographical areas, and climatic conditions. The ability of probiotic bacteria to survive in the digestive system is the most crucial component of probiotic research; however, there are certain other barriers to their survival. The two most important criteria for choosing probiotic strains are tolerance to low stomach pH and high concentrations of bile salts [20]. The stomach pH is approximately 3.0, whereas severe GIT conditions are typically simulated in vitro at a pH of 2.0. In this study, all five tested LAB isolates were able to survive with varying degrees of viability in simulated gastric fluid at pH 2.0 after 3 h of incubation. (Table 1, Fig. 1). These results are consistent with earlier research showing that LAB strains isolated from honey survive moderately to well in simulated gastric juice at a pH of 2.0 [21]. The ability of LAB to withstand acid is a beneficial characteristic that enhances their ability to ferment food, restores a balanced gut microbiome, and has several other beneficial effects on human health.
In the present study, P0-2 L, P14-5 L, and Con-6 L were found to belong to the same species of L. rhamnosus but exhibited differences in viability at pH 3 in the presence of pepsin and pH 8 in the presence of bile salts. These results are consistent with previous studies suggesting that resistance to gastric acidity varies depending on the duration of incubation and is strain-dependent [22]. Similarly, P01-1 L, P0-3 L, and P0-4 L are Enterococcus faecium species, but their viability differs under simulated gastric and intestinal conditions (Table 1, Fig. 1). It is commonly recognized that distinct bacterial strains belonging to the same genus and species can have entirely distinct effects on their host. As a result, each strain's unique characteristics should be identified, and each strain's impact on health should be evaluated individually in clinical trials.
It is well-recognized that probiotics, such as Lacticaseibacillus, Enterococcus, and Bifidobacterium, prevent the growth of numerous intestinal pathogens in humans. In this study, we described the inhibitory activity of CFCS from a total of six LAB isolates (including one L. rhamnosus control strain), namely, L. rhamnosus and E. faecium, against clinically important pathogens. The tested LAB strains exhibited varying levels of antagonistic activity against the selected pathogens (Table 2). The CFCSs derived from L. rhamnosus (P0–2 L) and E. faecium (P0–4 L) demonstrated broad-spectrum activity against pathogenic microbes. P0-3 L, on the other hand, inhibited all gram-negative bacteria but was unable to inhibit gram-positive bacteria other than S. pneumoniae. P01-1 L inhibited all the gram-positive bacteria but did not impede the growth of all the gram-negative bacteria except E. coli. P14 did not inhibit any of the tested pathogenic bacteria. The results clearly show that the antibacterial activity of LAB against pathogenic bacteria is strain-specific, which is consistent with the results of previous studies [23].
One of the principal criteria for the selection of presumed probiotic strains is their ability to adhere to the intestinal epithelium and subsequently colonize the gastrointestinal tract. A study conducted by Lee and Yii (1996) classified bacterial strains into three groups on the basis of their degree of adherence to hydrocarbons: a bacterial cell is considered strongly hydrophobic if its adhesion percentage is at least 50%, hydrophile if it is less than 20%, and moderately hydrophobic if the rate falls within these two ranges [24]. Based on these criteria, E. faecium (P01-1 L), L. rhamnosus (P0-2 L), and E. faecium (P0-3 L) were strongly hydrophobic, whereas E. faecium (P0-4 L), L. rhamnosus (P14-5 L), and L. rhamnosus (Con-6 L) were moderately hydrophobic (Table 3). There were no strains that were hydrophiles. The results showed that the three tested isolates displayed greater hydrophobicity than did the commercially available probiotic strain L. rhamnosus (Con-6 L).
To eliminate the pathogenic bacteria from the digestive tract, the coaggregation is a desired feature of probiotics, needed for inhibiting adherence to the host tissue. Therefore, the probiotics initial screening methods employed pathogenic bacterial coaggregation assay, signifying its safety for both humans and animals, as described by Dlamini et al. (2019)[25]. All LAB strains were coaggregated with pathogenic strains, depending on the combination of the strain and the pathogen [25]. On the contrary, one study reported no coaggregation between the pathogens and LAB isolates examined [26]. In this study, all the tested LAB isolates presented moderate to high percentages of colonization attributes, including cell surface hydrophobicity, coaggregation, and autoaggregation. Among them, E. faecium (P0-3 L) presented the highest percentage of these properties. Safety aspects of probiotic strains are of utmost importance and prior to marketing probiotic cultures, safety factors like hemolysis in blood agar media must be assessed. In this study, the LAB strains showed no hemolytic activity on 5% sheep blood agar after 48 hours of incubation at 37°C. These findings suggest that LAB isolates may be safe for human consumption. Previous studies [27] findings are in concordance with our study findings.
When evaluating the safety of probiotics, resistance to antibiotics is another crucial index parameter; therefore, the susceptibility of all potential probiotic strains to a variety of widely prescribed antibiotics should be evaluated. This process is crucial for identifying putative probiotic microorganisms that carry genes for transferable antibiotic resistance, which could be harmful to the host [27]. The results revealed that all the tested LAB isolates were susceptible to several different antibiotic classes (Table 4). However, they were resistant to fusidic acid, sulfamethoxazole, and colistin sulfate. Generally, antibiotic resistance in LAB does not represent a safety risk by itself when intrinsic resistance mechanisms or mutations cause a resistance phenotype; rather, the use of antibiotic-resistant LAB as a probiotic is advantageous since these bacteria can withstand antibiotic treatment and restore the gut microbiota [28]. Sulfamethoxazole, colistin sulfate, and fusidic acid resistance in LAB has been reported in previous studies and has always been linked to their innate and intrinsic resistance, most likely as a result of their membrane impermeability and cell wall structure, supplemented in certain instances by potential efflux mechanisms [29]. The major limitation of our study is that we could not check the probiotic properties of isolated lactic acid bacteria in animal models due to time and financial constraints.
Researchers have been interested in Lacticaseibacillus strains for a long time because of their possible use in the food sector and gastrointestinal tract disorders as probiotics. L. rhamnosus (P0-2 L) isolated from Sidr honey has potential probiotic characteristics, such as acid resistance and bile salt tolerance, a broad spectrum and potent antagonistic potential against six clinically important pathogens, even better than that of the commercially available probiotic strain L. rhamnosus, which has remarkably high adhesive potential and good autoaggregation capacity. With respect to the assessment of safety, the strain was negative for hemolytic activity and resistant to three out of sixteen tested antibiotics. The resistance of LAB to these antibiotics is generally innate and intrinsically driven, suggesting that this strain is nonvirulent and thus safe for human consumption.
Acknowledgements
This project was funded by the Deanship of Scientific Research (DSR) at King Abdulaziz University, Jeddah, under grant no. (GPIP: 1909-828- 2024). The authors, therefore, acknowledge and thank DSR for their technical and financial support.
Author Contributions
Conceptualization: [Muhammad Barkaat Hussain, Muhammad Absar], Methodology: [Muhammad Yasir, Yasser Mahmoud Kamel Mohamed Aly, Asif Ahmad M. Jiman-Fatani], Formal analysis and investigation: [Muhammad Barkaat Hussain, Asif Ahmad M. Jiman-Fatani, Sayed Sartaj Sohrab, Muhammad Absar], Writing – original draft preparation: [Muhammad Barkaat Hussain, Muhammad Absar]; Writing – review and editing: [Muhammad Barkaat Hussain, Muhammad Absar, Sayed Sartaj Sohrab, Saleh M. Al-Maaqar, Muhammad Yasir], Supervision: [Muhammad Barkaat Hussain]
The authors declare that there is no conflict of interest.
![]() References
References
- Ibrahim SA, Yeboah PJ, Ayivi RD, Eddin AS, Wijemanna ND, Paidari S et al. A review and comparative perspective on health benefits of probiotic and fermented foods. International Journal of Food Science & Technology, (2023); 58(10): 4948-4964.
- Pineiro M, Stanton C. Probiotic bacteria: legislative framework—requirements to evidence basis. The Journal of nutrition, (2007); 137(3): 850S-853S.
- Wang Y, Jiang Y, Deng Y, Yi C, Wang Y, Ding M et al. Probiotic supplements: hope or hype? Frontiers in microbiology, (2020); 11: 160.
- Rad AH, Elnaz VM, Beitullah A, Leila VM. The comparison of food and supplement as probiotic delivery vehicles. Critical reviews in food science and nutrition, (2016); 56(6): 896-909.
- Alkalbani NS, Osaili TM, Al-Nabulsi AA, Obaid RS, Olaimat AN, Liu SQ et al. In vitro characterization and identification of potential probiotic yeasts isolated from fermented dairy and non-dairy food products. Journal of fungi (Basel, Switzerland), (2022); 8(5): 544.
- Duche RT, Singh A, Wandhare AG, Sangwan V, Sihag MK, et al. Antibiotic resistance in potential probiotic lactic acid bacteria of fermented foods and human origin from Nigeria. BMC microbiology, (2023); 23(1): 142.
- Yadav MM, Shukla P. Probiotics of diverse origin and their therapeutic applications: a review. Journal of the American College of Nutrition, (2020); 39(5): 469-479.
- Dahiya D, Nigam PS. Biotherapy Using Probiotics as Therapeutic Agents to Restore the Gut Microbiota to Relieve Gastrointestinal Tract Inflammation, IBD, IBS and Prevent Induction of Cancer. International journal of molecular sciences, (2023); 24(6): 5748.
- Minj J, Chandra P, Paul C, Sharma RK. Bio-functional properties of probiotic Lactobacillus: current applications and research perspectives. Critical reviews in food science and nutrition, (2021); 61(13): 2207-2224.
- Li HY, Zhou DD, Gan RY, Huang SY, Zhao CN, Shang A et al. Effects and mechanisms of probiotics, prebiotics, synbiotics, and postbiotics on metabolic diseases targeting gut microbiota: A narrative review. Nutrients, (2021); 13(9): 3211.
- Kumar D, Lal MK, Dutt S, Raigond P, Changan SS, Tiwari RK et al. Functional Fermented Probiotics, Prebiotics, and Synbiotics from Non‐Dairy Products: A Perspective from Nutraceutical. Molecular nutrition & food research, (2022); 66(14): 2101059.
- Castro-López C, Romero-Luna HE, García HS, Vallejo-Cordoba B, González-Córdova AF, Hernández-Mendoza A. Key stress response mechanisms of probiotics during their journey through the digestive system: a review. Probiotics and antimicrobial proteins, (2023); 15(5): 1250-1270.
- Ferdouse J, Paul S, Chowdhury T, Ali F, Islam S, Hossain TJ. Probiotic characteristics of Pediococcus pentosaceus and Apilactobacillus Kunkeei strains: the lactic acid bacteria isolated from Bangladeshi natural honey. Applied Food Biotechnology, (2022); 10(1): 33-45.
- Hussain MB, Aly YMKM, Jiman-Fatani AAM, Ullah Z, Qureshi IA,Bakarman MA et al. Screening Of Lactic Acid Bacteria From Selected Saudi Honey Produced By Apis Mellifera Jementica. Journal of Animal & Plant Sciences, (2023); 33(2): 249-263.
- Al-Masaudi SB, Hussain MB, Al-Maaqar SM, Al Jaouni S, Harakeh S. In vitro antibacterial activity of honey against multidrug-resistant Shigella sonnei. Complementary Therapy in Clininical Practice, (2020); 41: 101257.
- Hussain MB, Hannan A, Akhtar N, Fayyaz GQ, Imran M, Saleem S et al. Evaluation of the antibacterial activity of selected Pakistani honeys against multi-drug resistant Salmonella typhi. BMC complementary and alternative medicine, (2015); 15(1): 1-9.
- Darmastuti A, Hasan PN, Wikandari R, Utami T, Rahayu ES, Suroto DA. Adhesion Properties of Lactobacillus plantarum Dad-13 and Lactobacillus plantarum Mut-7 on Sprague Dawley Rat Intestine. Microorganisms, (2021); 9(11): 2336.
- Kiehlbauch JA, Hannett GE, Salfinger M, Archinal W, Monserrat C, Carlyn C. Use of the National Committee for Clinical Laboratory Standards guidelines for disk diffusion susceptibility testing in New York state laboratories. Journal of clinical microbiology, (2000); 38(9): 3341-3348.
- Al-Kafaween MA, Alwahsh M, Mohd Hilmi AB, Abulebdah DH. Physicochemical Characteristics and Bioactive Compounds of Different Types of Honey and Their Biological and Therapeutic Properties: A Comprehensive Review. Antibiotics (Basel, Switzerland), (2023); 12(2): 337.
- Ayyash MM, Abdalla AK, AlKalbani NS, Baig MA, Turner MS, Liu SQ et al. Invited review: Characterization of new probiotics from dairy and nondairy products—Insights into acid tolerance, bile metabolism and tolerance, and adhesion capability. Journal of dairy science, (2021); 104(8): 8363-8379.
- Meradji M, Bachtarzi N, Mora D, Kharroub KJF: Characterization of Lactic Acid Bacteria Strains Isolated from Algerian Honeybee and Honey and Exploration of Their Potential Probiotic and Functional Features for Human Use. Foods (Basel, Switzerland), (2023); 12(12): 2312.
- Choi O, Lee Y, Kang B, Cho SK, Kang Y, Kang DW et al. Identification and characterization of gut-associated lactic acid bacteria isolated from the bean bug, Riptortus pedestris (Hemiptera: Alydidae). PloS one, (2023); 18(3): e0281121.
- Prabhurajeshwar C, Chandrakanth KJ. Evaluation of antimicrobial properties and their substances against pathogenic bacteria in-vitro by probiotic Lactobacilli strains isolated from commercial yoghurt. Clinical Nutrition Experimental, (2019); 23: 97-115.
- Lee KK, Yii KC. A comparison of three methods for assaying hydrophobicity of pathogenic vibrios. Letters in Applied Microbiology, (1996); 23(5): 343-346.
- Dlamini ZC, Langa RL, Aiyegoro OA, Okoh A. Safety evaluation and colonisation abilities of four lactic acid bacteria as future probiotics. Probiotics and antimicrobial proteins, (2019); 11(2): 397-402.
- Taheri H, Moravej H, Tabandeh F, Zaghari M, Shivazad M. Screening of lactic acid bacteria toward their selection as a source of chicken probiotic. Poultry science, (2009); 88(8): 1586-1593.
- Goh LPW, Molujin AM, Muthu K, Abdulla R, Sabullah MK, Faik AAM et al. Isolation and characterization of lactic acid bacteria from Sabah (North Borneo) stingless bees for probiotic and food applications. International Journal of Food Properties, (2021); 24(1): 564-578.
- Gueimonde M, Sánchez B, G. de los Reyes-Gavilán C, Margolles AJ. Antibiotic resistance in probiotic bacteria. Frontiers in microbiology, (2013); 4: 202.
- Nikodinoska I, Heikkinen J, Moran CA. Antimicrobial Susceptibility Data for Six Lactic Acid Bacteria Tested against Fifteen Antimicrobials. Data, (2023); 8(1):10.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0![]()