

Review Article
The role of bacteriophages transferring virulence factors to Escherichia coli species
Ibrahim Alotibi
Adv. life sci., vol. 10, no. 1, pp. 17-21, March 2023
*– Corresponding Author: Ibrahim Alotibi (Email: ialotibi@kau.edu.sa)
Authors' Affiliations
[Date Received: 24/06/2022; Date Revised: 24/01/2023; Date Published Online: 31/03/2023; Date Updated: 05/09/2025]
Editorial Expression of Concern
20 June 2025: Following publication of this paper, the internal audit (consequent to concerns on quality raised by Web of Science) notified Advancements in Life Sciences about errors in metadata. By this Editorial Expression of Concern, we alert the scientific community of this incidence as we do the required corrections.
Editorial Note
You are viewing latest version of this article having language corrections.
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
Bacteria develop in order to adapt to new surroundings, colonize new niches, and become pathogenic. The presence of mobile genetic elements MGEs in E. coli can increase the genome size of a pathogenic strain by up to 1 Mb when compared to a commensal strain. Phage satellites make up one subset of MGEs; they are linked to specific temperate phages, known as helper phages, which parasitize bacteria for their own induction. In fact, various pathogenic E. coli differ in the presence of a subset of genes produced by MGEs that are crucial in hijacking host cell machinery and subverting host responses. Phages not only provide genetic variability through prophage integration, but they can also mediate horizontal genetic transfer HGT within bacterial populations through the transfer of either bacterial DNA or other MGEs, such as phage satellites. The phage-mediated transfer of bacterial DNA is known as transduction and plays a crucial role in bacterial biology, diversity and evolution. Recently, it has been noticed that phage transduction occurs at an astonishing magnitude, much higher than previously anticipated. Importantly, some of the genes transferred by transduction are virulence and antibiotic resistance genes, highlighting the impact that this process has in driving the evolution of pathogenic bacteria.
Keywords: E. coli; HGT; MGEs; Bacteriophages; Transduction; virulence Genes
Introduction![]()
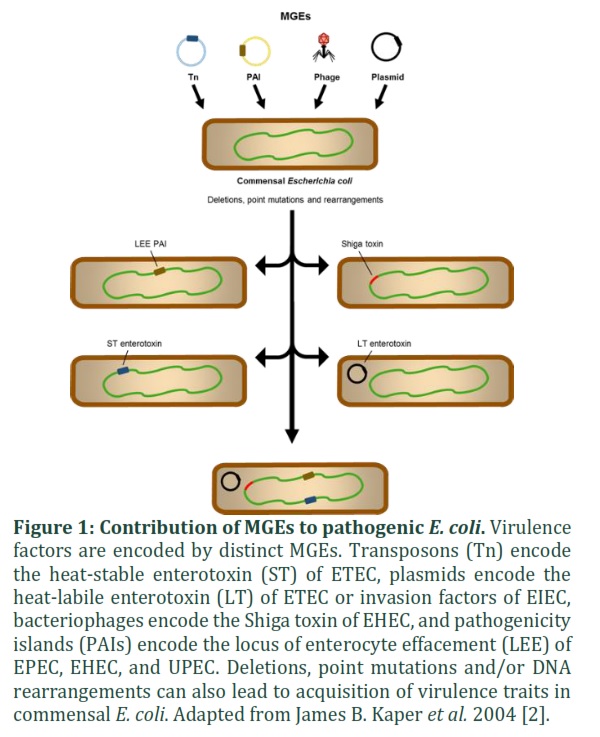
Escherichia coli belongs to the Proteobacteria phylum and is a facultative anaerobic Gram-negative bacterium that does not sporulate. E. coli forms rod-shaped colonies that can be grouped singly or in pairs, and it is motile due to peritrichous flagellae. E. coli can be a harmless commensal or a clinically important opportunistic pathogen [1]. E. coli colonizes the gastrointestinal system of new-borns, with the colon, notably the mucous layer, as its habitat. To become an adapted pathogen, E. coli just needs to acquire one or a combination of various mobile genetic elements (MGEs).
MGEs have the capability to adapt to new niches and cause a wide range of illnesses, including gastroenteritis (diarrhoea), dysentery, bloodstream infections, sepsis, and urinary tract and central nervous system infections (meningitis). Furthermore, virulent E. coli strains can arise from deletions, point mutations, and genomic rearrangements [2].
Different criteria can be used to divide E. coli, including type, serotype, pulsotype, phage type, and biotype [4]. The pathotype, which refers to the various diseases that pathogenic E. coli may cause, is the most prevalent characteristic used to categorize pathogenic E. coli.
Several pathotypes, eight of which are well characterized, as intestinal (diarrheagenic) or extraintestinal E. coli (ExPEC) [2,3]. Six different pathotypes are included as intestinal: i) enteropathogenic E. coli (EPEC), ii) enterohaemorrhagic E. coli (EHEC), iii) enterotoxigenic E. coli (ETEC), iv) enteroinvasive E. coli (EIEC; including Shigella), v) enteroaggregative E. coli (EAEC); and vi) diffusely adherent E. coli (DAEC) [2,5-16]. The two most common pathotypes categorized as extraintestinal are: i) uropathogenic E. coli (UPEC); and ii) neonatal meningitis E. coli (NMEC) [17-26]. Other pathotypes have been proposed, although they have not been adequately defined. They include the following: necrotoxigenic E. coli (NTEC) or adherent invasive E. coli (AIEC) [27-30].
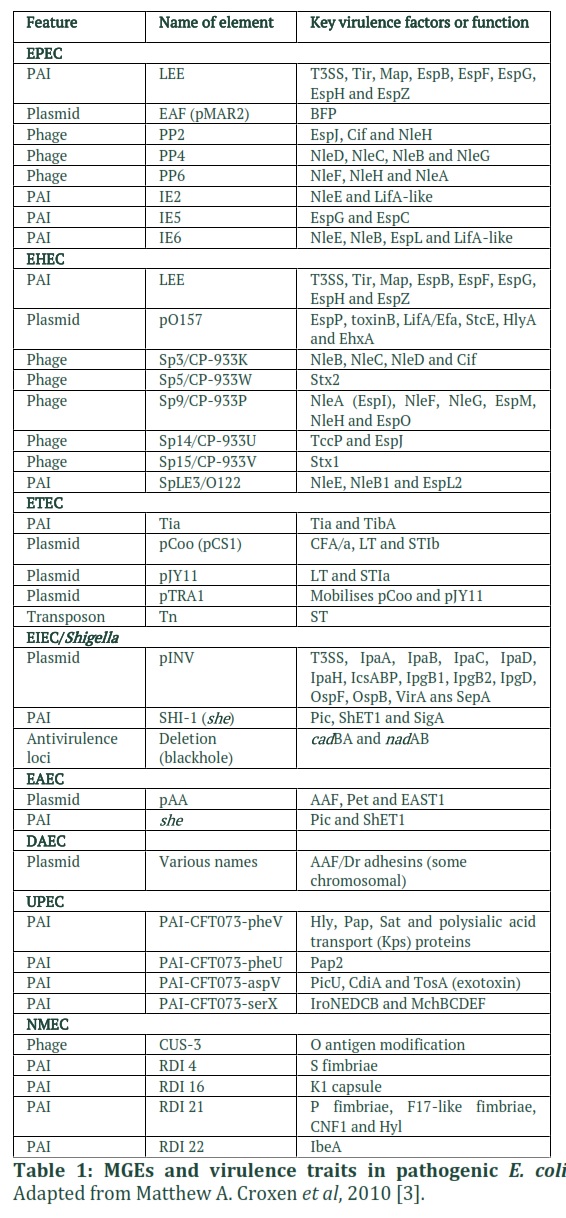
MGE acquisition or loss is essential for pathogenic bacteria to adapt to new or changing environmental conditions. In reality, each pathotype is distinguished by the presence of a group of genes involved in the hijacking of host cell machinery and the subversion of host responses [3,10]. Despite the fact that the same host machines or processes are attacked, the mechanisms and results are different. When compared to commensal E. coli, virulence-associated genes expressed by MGEs can increase the genome size of pathogenic E. coli by up to 1 Mb (Table 1) [18]. For example, in the UPEC strain CFT073, A total of 13 genomic islands have been discovered, accounting for up to 13% of the bacterial genome [31].
Methods![]()
Literature Search and Selection Criteria
Raw data was organized using Microsoft® Excel® 2010. The NCBI BLAST server program (www.ncbi.nlm.nih.gov) has been used to compare sequences with the GenBank database for homology. Sequences were assembled to a reference, and multiple nucleotide or protein sequence alignments were performed using CLC Genomics Workbench 7.
Figures & Tables
Discussion![]()
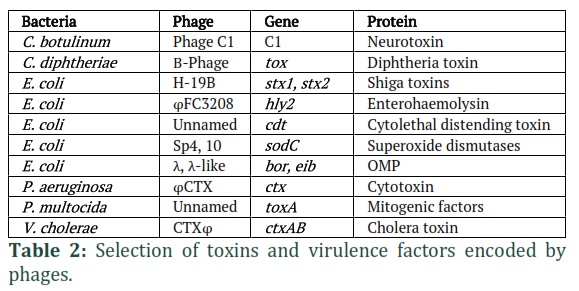
Temperate phages have an impact the genomes of their hosts, causing genetic variation in their cognate cells [32]. In the genome of the E. coli strain O157:H7, for example, 18 distinct prophages have been discovered. This strain belongs to the EHEC pathotype and can contain phages that encode the Shiga toxin protein (Stx) [34]. This difference is important in virulent strains because lysogenic conversion (phage integration into the bacterial genome) provides the host bacterium with virulence proteins and other phage-encoded genes that are necessary for colonization of new habitats. As a result, prophages constitute a significant source of genetic diversity that contributes to the pathogenicity of bacteria [35-38]. Toxins, adhesion factors, invasion factors, and superantigens, among other phage-encoded virulence factors, are included in Table 2 [32,39]. Toxin-mediated illnesses, including as botulism, cholera, diarrhoea, diphtheria, and scarlet fever, are caused by toxins encoded by phages [40]. Virulence genes in E. coli phages, especially Stx, can cause attaching and effacing (A/E) lesions, bloody diarrhoea, and haemolytic uremic syndrome (HUS) [2].
The expression, release, and mobilization of these toxins are all linked to the phage lytic cycle in some situations. This is relevant in the treatment of prophage-containing pathogenic bacteria because certain antibiotics can cause the SOS response and, as a result, the phage lytic cycle to begin. Finally, phage induction will increase toxin expression, and the toxins will be released once the cells are lysed by the phage. The Stx encoded by E. coli EHEC phages is one example of toxin production associated with prophage induction. Stx expression raises the risk of A/E lesions, diarrhea, and HUS when EHEC strains are treated with antibiotics such as fluoroquinolones [33].
Conclusion
Bacteria develop to adapt to new surroundings, colonize new niches, and become pathogenic. HGT and MGEs are important participants in this evolutionary process because they can transmit advantageous characteristics or virulence factors between bacterial species [41, 42]. The presence of MGEs in E. coli can increase the genome size of a pathogenic strain by up to 1 Mb when compared to a commensal strain [18]. Indeed, in the UPEC strain CFT073, 13 distinct genomic islands have been found, accounting for 13% of the bacterial genome [31]. In fact, various pathogenic E. coli differs in the presence of a subset of genes produced by MGEs that are crucial in hijacking host cell machinery and manipulating host defences [3,10]. Phage satellites are one type of MGE. They are associated with specific temperate phages known as helper phages, which parasitize bacteria for their own induction, allowing transmission to a new host bacterium.
The authors declare that there is no conflict of interest.
![]()
References
- Sousa, CP. The versatile strategies of Escherichia coli pathotypes: a mini review. Journal of Venomous Animals and Toxins including Tropical Diseases, (2006) ; 12; 363–373.
- Kaper, JB, Nataro, JP. & Mobley, HL. T. Pathogenic Escherichia coli. Nature Reviews Microbiology, (2004) ; 2; 123–140.
- Croxen, MA & Finlay, BB. Molecular mechanisms of Escherichia coli pathogenicity. Nature Reviews Microbiology, (2010) ;
- 8;26–38.
- Roy, MR, Kathryn EH, Danielle JI, Dianna MH, JY and MT. Are Escherichia coli Pathotypes Still Relevant in the Era of Whole-Genome Sequencing. Frontiers in cellular and infection microbiology, (2016) ; 6; 141.
- Nataro,JP,Kaper,JB.Diarrheagenic Escherichia coli. Clinical microbiology reviews, (1998) ;11; 142–201.
- McDaniel,TK, Jarvis,KG,Donnenberg,MS,& Kaper, JBA. Genetic locus of enterocyte effacement conserved among diverse enterobacterial pathogens. Proceedings of the National Academy of Sciences of the United States of America, (1995) ; 92; 1664–8.
- Jerse, AE, Yu, JT, Kaper, JBA .Genetic locus of enteropathogenic Escherichia coli necessary for the production of attaching and effacing lesions on tissue culture cells. Proceedings of the National Academy of Sciences of the United States of America, (1990) ;87; 7839–43.
- Klapproth, J M, et al. A large toxin from pathogenic Escherichia coli strains that inhibits lymphocyte activation. Infection and immunity, (2000) ;68; 2148–55 .
- Toshima,H,et al. Enhancement of Shiga Toxin Production in Enterohemorrhagic Escherichia coli Serotype O157:H7 by DNase Colicins. Applied and Environmental Microbiology, (2007) ; 73; 7582–7588.
- Bhavsar,AP, Guttman, JA & Finlay, BB. Manipulation of host-cell pathways by bacterial pathogens. Nature,(2007) ;449; 827–834.
- Weiss, SM, et al. IRSp53 Links the Enterohemorrhagic E. coli Effectors Tir and EspFU for Actin Pedestal Formation. Cell Host & Microbe, (2009) ;5; 244–258.
- Tran VN,G, BS,R, D, G, Blocker,A, Sansonetti, PJ. Bacterial signals and cell responses during Shigella entry into epithelial cells. Cellular microbiology, (2000) ; 2; 187–93.
- Sansonetti, PJ et al. Caspase-1 activation of IL-1beta and IL-18 are essential for Shigella flexneri-induced inflammation. Immunity, (2000) ; 12; 581–90.
- Buchrieser,C et al.The virulence plasmid pWR100 and the repertoire of proteins secreted by the type III secretion apparatus of Shigella flexneri. Molecular microbiology, (2000) ;38;760–71.
- Bilge, SS, Clausen, CR, Lau, W, Moseley, SL. Molecular characterization of a fimbrial adhesin, F1845, mediating diffuse adherence of diarrhea-associated Escherichia coli to HEp-2 cells. Journal of bacteriology,(1989) ;171; 4281–9.
- Servin, AL. Pathogenesis of Afa/Dr diffusely adhering Escherichia coli. Clinical microbiology reviews, (2005).
- Wiles,TJ, Kulesus, RR, Mulvey, MA. Origins and virulence mechanisms of uropathogenic Escherichia coli. Experimental and Molecular Pathology, (2008) ; 85; 11–19.
- Welch, R et al. Extensive mosaic structure revealed by the complete genome sequence of uropathogenic Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America, (2002) ; 99; 17020–17024.
- Johnson, JR. Virulence factors in Escherichia coli urinary tract infection. Clinical microbiology reviews, (1991) ; 4; 80–128.
- Johnson,JR, Stell, AL. Extended Virulence Genotypes of Escherichia coli Strains from Patients with Urosepsis in Relation to Phylogeny and Host Compromise. The Journal of Infectious Diseases, (2000) ; 181; 261–272.
- Mulvey, MA ,et al. Induction and evasion of host defenses by type 1-piliated uropathogenic Escherichia coli. Science (New York, N.Y.), (1998) ; 282; 1494–7.
- Anderson,GG,et al. Intracellular Bacterial Biofilm-Like Pods in Urinary Tract Infections. Science, (2003) ; 301; 105–107.
- Dawson, KG, Emerson, JC, Burns, JL. Fifteen years of experience with bacterial meningitis. The Pediatric infectious disease journal, (1999) ; 18; 816–22.
- Badger, JL, Wass, CA, Weissman, SJ, Kim, KS. Application of signature-tagged mutagenesis for identification of escherichia coli K1 genes that contribute to invasion of human brain microvascular endothelial cells. Infection and immunity, (2000) ; 68; 5056–61.
- Deszo, EL,Steenbergen, SM, Freedberg, DI, Vimr, ER. Escherichia coli K1 polysialic acid O-acetyltransferase gene, neuO, and the mechanism of capsule form variation involving a mobile contingency locus. Proceedings of the National Academy of Sciences of the United States of America, (2005) ;102; 5564–9.
- Dietzman,D E, Fischer, GW, Schoenknecht, FD. Neonatal escherichia coli septicemia—bacterial counts in blood. The Journal of Pediatrics, (1974) ; 85; 128–130.
- Barnich,N,et al. CEACAM6 acts as a receptor for adherent-invasive E. coli, supporting ileal mucosa colonization in Crohn disease. Journal of Clinical Investigation, (2007) ; 117; 1566–1574.
- Darfeuille-Michaud,A. Adherent-invasive Escherichia coli: a putative new E. coli pathotype associated with Crohn’s disease. International Journal of Medical Microbiology, (2002) ; 292; 185–193.
- Carvalho,FA,et al.Crohn’s disease adherent-invasive Escherichia coli colonize and induce strong gut inflammation in transgenic mice expressing human CEACAM. The Journal of Experimental Medicine, (2009) ; 206; 2179–2189.
- De Rycke,J,Milon, A, Oswald, E.Necrotoxic Escherichia coli (NTEC): two emerging categories of human and animal pathogens. Veterinary research,(1999) ; 30 ; 221–33.
- Lloyd,AL,Rasko,DA, Mobley, HL,T. Defining Genomic Islands and Uropathogen-Specific Genes in Uropathogenic Escherichia coli. Journal of Bacteriology, (2007) ; 189; 3532–3546.
- Brussow,H,Canchaya,C,Hardt,WD. Phages and the Evolution of Bacterial Pathogens: from Genomic Rearrangements to Lysogenic Conversion. Microbiology and Molecular Biology Reviews, (2004) ; 68; 560–602.
- Zhang, X,et al. Quinolone antibiotics induce Shiga toxin-encoding bacteriophages, toxin production, and death in mice. The Journal of infectious diseases, (2000) ; 181; 664–670.
- Shaikh, N. ,Tarr, PI. Escherichia coli O157:H7 Shiga toxin-encoding bacteriophages: integrations, excisions, truncations, and evolutionary implications. Journal of bacteriology ,(2003) ; 185; 3596–605.
- Baba,T,et al. Genome and virulence determinants of high virulence community-acquired MRSA. Lancet,(2002) ; 359;1819–1827.
- Kwan,T,Liu,J ,DuBow, MG,Pelletier, J. The complete genomes and proteomes of 27 Staphylococcus aureus bacteriophages. Proceedings of the National Academy of Sciences,(2005) ; 102; 5174–5179.
- Sumby, P,Waldor, MK. Transcription of the toxin genes present within the Staphylococcal phage phiSa3ms is intimately linked with the phage’s life cycle. Journal of bacteriology,(2003) ; 185; 6841–51.
- Waldor, MK,Friedman, DI. Phage regulatory circuits and virulence gene expression. Current Opinion in Microbiology,(2005) ; 8; 459–465.
- Ross, J,Topp, E. Abundance of antibiotic resistance genes in bacteriophage following soil fertilization with dairy manure or municipal biosolids, and evidence for potential transduction. Applied and Environmental Microbiology, (2015) ;81;7905–7913.
- Novick, RP.Mobile genetic elements and bacterial toxinoses: The superantigen-encoding pathogenicity islands of Staphylococcus aureus. Plasmid, (2003) ; 49;93–105.
- Brüssow, H, Hendrix, RW. Phage Genomics: Small is beautiful. Cell, (2002) ; 108; 13–16.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0